巴西橡胶树HbPSKR2基因克隆及互作蛋白鉴定
2024-06-09杜晓愚赵一杰张世鑫田维敏晁金泉
杜晓愚 赵一杰 张世鑫 田维敏 晁金泉


关键词:巴西橡胶树;HbPSKR2;酵母双杂交;荧光素酶互补成像;互作蛋白
中图分类号:S794.1 文献标志码:A
磺肽素(phytosulfokine, PSK)是一种高等植物特有的小肽类激素,广泛参与植物的生长发育、分裂分化、逆境胁迫等生物学过程[1-3]。磺肽素基因编码的前体多肽长度约为80~110 个氨基酸,通过翻译后剪切形成5 个氨基酸的小肽( Tyr-Ile-Tyr-Thr-Gln),而后在酪氨酸硫化转移酶的作用下对其第1 位和第3 位的酪氨酸(tyrosine, Tyr)发生特异的磺基化修饰,最终形成有活性形式的磺肽素[4-5]。与其他植物激素一样,磺肽素信号传导也依赖于细胞膜定位的受体蛋白。磺肽素受体(phytosulfokine receptor, PSKR)是一种典型的跨膜蛋白,其膜外的亮氨酸重复元件(leucine richrepeat, LRR)主要用于感知磺肽素信号,而膜内的激酶结构域主要用于下游信号的传导[6-7]。PSKR 在植物中一般有2 个成员,目前对其功能的研究还比较少。ZHANG 等[8]发现番茄的SlPSKR1和SlPSKR2 均可与PSK 结合,并且证明PSKR1通过促进钙调素与生长素合成蛋白YUC 的互作进而激活番茄对疫霉病的响应。HOLZWART 等[9]报道拟南芥的PSKR1 可以和RLP44 蛋白互作,在调控形成层向木质部分化过程中起关键作用。
天然橡胶是一种重要的工业原料,在医疗卫生、交通运输、航空航天等领域具有广泛应用[10-11]。巴西橡胶树(Hevea brasiliensis Muell. Arg)原产南美洲亚马逊雨林地区,是当前世界天然橡胶的主要来源[12]。位于橡胶树树皮内层的次生乳管是天然橡胶合成和贮存的主要场所。橡胶树胶乳本质上是乳管细胞的细胞质部分,含30%~50%的天然橡胶[13]。生产上人们通过割胶的方式收集从被切断乳管中流出的胶乳用于天然橡胶加工,因此橡胶树的次生乳管是决定天然橡胶产量的结构基础。前人研究表明,橡胶树次生乳管由形成层分化而来,茉莉酸信号途径在该过程中起关键作用[14-15]。最近课题组研究发现磺肽素参与了茉莉酸诱导次生乳管分化[16]。本研究克隆了橡胶树磺肽素受体HbPSKR2,并对其互作蛋白进行筛选和鉴定,为深入揭示橡胶树乳管分化分子机制提供新的思路。
1 材料与方法
1.1 材料
橡胶树形成层区cDNA、橡胶树形成层区酵母双杂交文库、酵母双杂交诱饵载体pGBKT7、pCAMBIA1300-nLUC、pCAMBIA1300-cLUC 均由本实验室保存;本生烟播种于中国热带农业科学院橡胶研究所植物培养室,培养至4~7 叶期备用;农杆菌菌株GV3101、酵母菌株Y187、酵母菌株AH109、大肠杆菌DH5α 感受态细胞均购自上海唯地生物技术有限公司;橡胶树全基因组序列从NCBI 下载(https://www.ncbi.nlm.nih.gov/nuccore/LVXX00000000.1);组织特异性及割胶前后胶乳的转录组原始数据从国家基因组数据中心下载(https://ngdc.cncb.ac.cn),GSA 编号为CRA004268;冠菌素(COR)处理形成层的转录组原始数据从NCBI 下载(https://www.ncbi.nlm.nih.gov/s),数据编号分别为SRR3423347~SRR3423350。
1.2 方法
1.2.1 HbPSKR2 基因的克隆 以拟南芥AtPSKR2蛋白序列对橡胶树基因组进行BLASTn 搜索,得到含橡胶树完整HbPSKR2 序列的scaffold0093_1558507。根据序列信息设计扩增HbPSKR2 完整开放阅读框的特异性引物HbPSKR2-F 和HbPSKR2-R(表1),以橡胶树形成层区cDNA 为模板进行PCR 扩增。扩增体系为:2×PCR buffer 10 μL,正、反向引物(10 μmol/L)各1 μL,cDNA 模板1 μL,ddH2O 7 μL;扩增程序为:98 ℃预变性5 min;98 ℃变性20 s,56 ℃退火30 s,72 ℃延伸3 min,共设置33 个循环;最终72 ℃延伸10 min。扩增产物用普通琼脂糖凝胶DNA 回收试剂盒(天根生化科技有限公司)进行回收纯化,通过pEASYBluntCloning Kit(全式金生物科技有限公司)连接载体并转化大肠杆菌DH5α。过夜培养后挑选阳性克隆送至生工生物工程(上海)股份有限公司进行测序。
1.2.2 HbPSKR2 的生物信息学分析 利用在线工具Compute pI/Mw( http://web.expasy.org/compute_pi/)分析HbPSKR2 的分子量和等电点;利用在线结构域分析工具SMART(http://smart.embl-heidelberg.de/)分析HbPSKR2 的结构域;利用DNAMAN 软件分析HbPSKR2 与其他植物同源蛋白间的序列比对;利用在线工具iTQL(https://itol.embl.de/) 繪制HbPSKR2 与其他植物同源蛋白间的系统发育树;利用依托单位服务器,采用Tophat 软件将所下载的转录组原始数据比对到橡胶树基因组上,根据Cufflink 软件计算基因的FPKM 值。
1.2.3 HbPSKR2 的自激活检测 以测序正确的pEASY-HbPSKR2 质粒为模板,用特异性引物pGBDKT7-HbPSKR2-F 和pGBDKT7-HbPSKR2-R扩增目的片段,采用ClonExpress Ultra One StepCloning Kit(南京诺唯赞生物科技股份有限公司)同源重组方法构建pGBDKT7-HbPSKR2 的Bait 载体。将测序正确的质粒转化至酵母Y187菌株,涂布于SD/-Trp/-Leu、SD/-Trp/-Leu/-His/-Ade 固体培养基上,30 ℃倒置培养3 d 观察菌落生长情况。
1.2.4 HbPSKR2 的酵母双杂交文库筛选 将pGBDKT7-HbPSKR2 和橡胶树形成层酵母cDNA文库分别转化至Y187 和AH109 酵母感受态细胞中,分别涂布于SD/-Trp 和SD/-Leu 筛选平板,30 ℃恒温培养3~4 d。用涂布棒刮取上述Prey 菌株和Bait 菌株,分别接种于相应筛选培养基中,30 ℃振荡培养至OD600=0.8。离心收集菌体,用YPDA 液体培养基重悬菌体并调整至细胞密度大于1×108mL–1。将二者等体积混匀,接种于YPDA液体培养基中扩大培养。在相差显微镜下观察酵母菌液产生合子后收集菌体,涂布SD/-Leu/-Trp筛选平板置于30 ℃倒置培养。挑取候选阳性菌落分别点板SD/-Leu/-Trp 二缺和SD/-Trp/-Leu/-His/-Ade 四缺筛选平板,30 ℃倒置观察菌落生长情况。挑选四缺平板阳性菌落送至生工生物工程(上海)股份有限公司进行测序。
1.2.5 荧光素酶互补验证 用特异性引物HbPSKR2-nLUC-F 和HbPSKR2-nLUC-R 扩增测序正确的pEASY-HbPSKR2 质粒并连接pCAMBIA1300-nLUC 载体;用特异性引物HbPBL8- cLUC-F/HbPBL8-cLUC-R 和HbPIX13-cLUC-F/HbPIX13-cLUC-R 分别扩增测序正确的阳性酵母菌株并连接pCAMBIA1300-cLUC 载体。将连接好的载体转化大肠杆菌DH5α 过夜培养并送样测序,将测序正确的pCAMBIA1300-HbPSKR2-nLUC、pCAMBIA1300-HbPBL8-cLUC 、pCAMBIA1300-HbPIX13-cLUC 质粒分别转化农杆菌GV 3101 感受态,并于YEB 液体培养基中28 ℃过夜培养。离心收集菌体,用培养液调整OD600=1.0,通过注射法注射烟草叶片。注射后的烟草暗培养24 h 后在相同注射部位注射荧光素酶底物D-荧光素钾盐溶液,置于活体成像系统中观察。
2 结果与分析
2.1 HbPSKR2 基因克隆及其编码蛋白分析
以HbPSKR2 开放阅读框引物对橡胶树形成层区cDNA 扩增,获得一条与预期大小一致的条带。测序结果显示,扩增产物长度为3159 bp,编码1052 个氨基酸(图1A)。HbPSKR2 的理论分子量为114.84 kDa,理论等电点为6.34。蛋白结构域分析显示HbPSKR2 是个典型的跨膜蛋白,其中前640 个氨基酸是由22 个LRR 结构域组成的膜外天线,第692 个氨基酸至第714 个氨基酸是跨膜结构域,第765 个氨基酸至第1052 个氨基酸是介导膜内信号途径的激酶结构域(图1B)。
2.2 HbPSKR2 系统进化及多序列比对分析
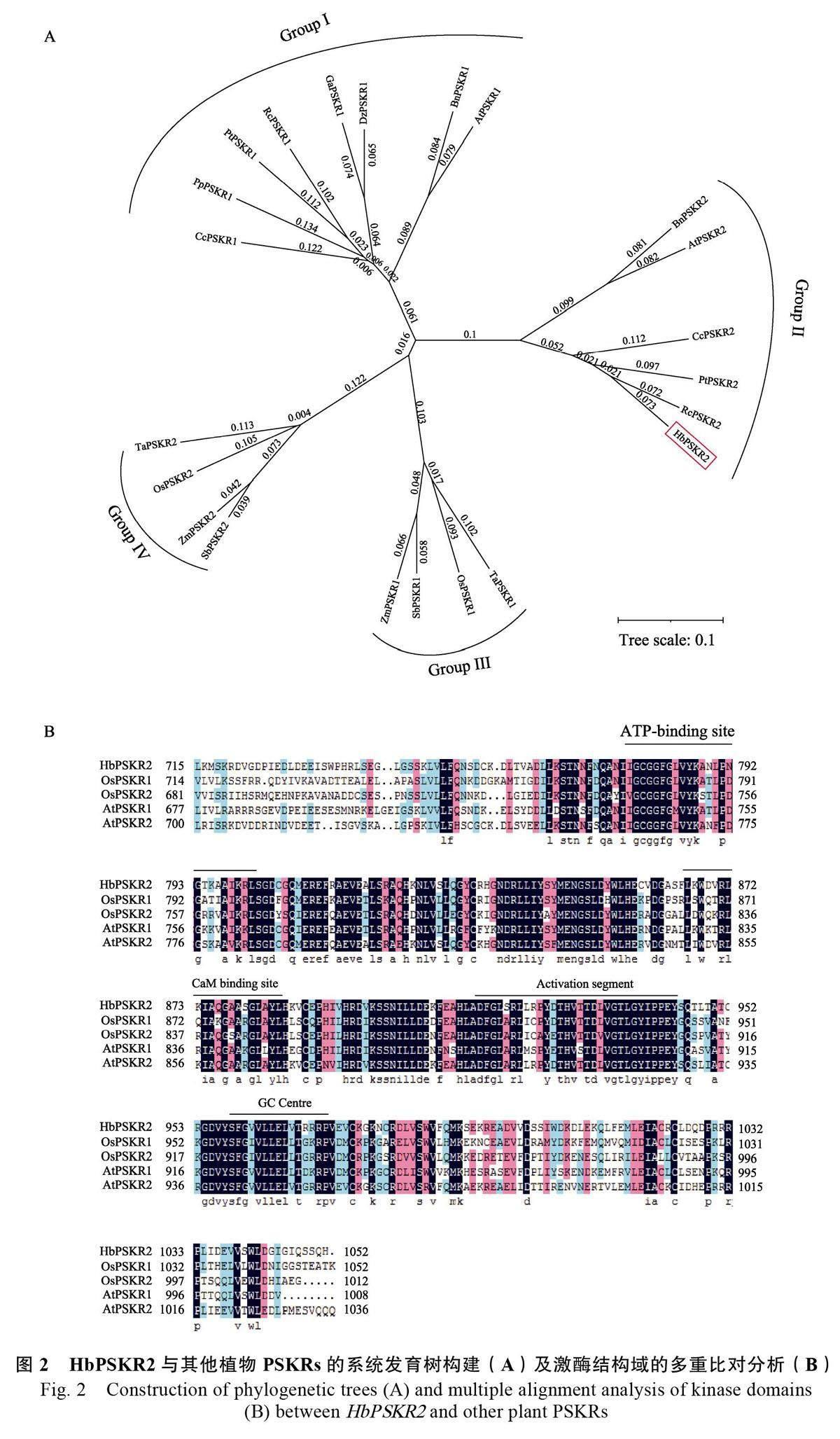
选取拟南芥(Arabidopsis thaliana, At2g02220,At5g53890)、樱桃(Prunus persica, XP_007227028.1)、榴莲(Durio zibethinus, XP_022776608.1 )、蓖麻(Ricinus communis, XP_002518809.2, EEF34126.1)、柚子(Citrus clementina, XP_024035321.1, XP_006451809.1)、树棉(Gossypium arboretum, XP_017632150.1 )、油菜( Brassica napus, XP_013707217.1, XP_022546871.2)、毛果杨(Populustrichocarpa, XP_002312507.3, XP_002317487.3)8种双子叶植物和水稻(Oryza sativa, LOC_Os02g41890.1, LOC_Os04g57630.1)、玉米(Zea mays,AQK72791.1, NP_001340572.1)、高粱(Sorghumbicolor, XP_002454207.1, XP_021318925.1)、小麦( Triticum aestivum, XP_044413269.1, XP_044335016.1)4 种单子叶植物的PSKR 序列,与HbPSKR2 共同构建系统发育树。结果显示所有PSKR 成员可以划分为4 个类群:Group I 类群由双子叶植物PSKR1 构成;Group II 類群由双子叶植物PSKR2 构成;Group III 类群由单子叶植物PSKR1 构成;Group IV 类群由单子叶植物PSKR2构成(图2A)。其中HbPSKR2 属于Group II 类群,且与来自蓖麻的RcPSKR2 亲缘关系最接近。
作为跨膜受体蛋白,PSKR 的膜内激酶结构域是介导信号传导的关键部分。采用DNAMAN软件对HbPSKR2 的膜内激酶结构域以及拟南芥和水稻的PSKR 同源蛋白的膜内激酶结构域进行多重比对,结果显示均存在ATP binding site、CaMbinding site、Activation segment、GC Centre 等保守性位点(图2B)。
2.3 HbPSKR2 基因表达模式分析
组织特异性分析显示,HbPSKR2 在橡胶树各个组织中均有表达,其中在形成层、雌花、雄花中表达丰度较高,而在胶乳、叶片、树皮中表达丰度较低(图3A)。冠菌素(coronatine, COR)是一种茉莉酸类似物,可以有效诱导橡胶树形成层分化次生乳管。HbPSKR2 的表达量在冠菌素处理前期(COR-A)较对照(CK-A)有显著上升,而在处理后期(COR-B)与对照(CK-B)相比则差异不显著(图3B)。割胶是收获天然橡胶的唯一方式。与未开割树相比,HbPSKR2 在开割树胶乳中的表达量极显著上升(图3C)。
2.4 HbPSKR2 诱饵载体自激活检测及互作蛋白筛选
采用重组法构建pGBKT7-HbPSKR2 诱饵载体。自激活实验结果显示,含pGBKT7-53 阳性对照载体的酵母菌在SD-TL 二缺培养基和SDTLHA四缺培养基上均可以生长,并且加入x-gal后菌落变蓝。而含pGBKT7-Lam 阴性对照载体的酵母菌以及含pGBKT7-HbPSKR2 载体的酵母菌只能在SD-TL 二缺培养基上生长(图4A),表明HbPSKR2 无细胞毒性及自激活活性,可以用于酵母文库筛选。
将含橡胶树形成层区酵母cDNA 文库质粒的酵母AH109 菌株和含pBGKT7-HbPSKR2 的酵母Y187 菌株共培养,镜检发现合子后收集菌体涂布至SD-TL 二缺平板进行培养,挑选阳性菌落接种于SD-TLHA 四缺平板进行进一步筛选。最终得到12 个阳性克隆(图4B)。对阳性克隆插入片段进行测序,测序结果在橡胶树数据库进行BLASTx同源比对,鉴定到2 个核糖体相关成员、2 个泛素化相关成员、2 个生长发育相关成员、2 个蛋白激酶、1 个RNA 聚合酶、1 个几丁质酶以及2 个功能未知蛋白(表2)。
2.5 HbPSKR2 与蛋白激酶HbPBL8 和HbPIX13 的互作验证
蛋白激酶在信号传导中起重要作用。在HbPSKR2互作蛋白中鉴定到2 个蛋白激酶HbPBL8 和HbPIX13,推测其可能介导橡胶树PSK 的信号传导。分别构建HbPSKR2-nLUC 载体、HbPBL8-cLUC 以及HbPIX13-cLUC 载体,采用LCI 技术在烟草叶片中验证HbPSKR2 与HbPBL8 和HbPIX13的互作关系。结果显示共转HbPSKR2-nLUC/HbPBL8-cLUC 和HbPSKR2-nLUC/HbPIX13-cLUC 可以检测到强烈的荧光信号(图5A),进一步证明HbPSKR2 在体内可以与HbPBL8 激酶和HbPIX13激酶互作。
2.6 HbPBL8 和HbPIX13 响应COR 处理的表达模式分析
通过分析COR 处理的橡胶树形成层区转录组数据,HbPBL8 表达量在COR 处理前期和后期均较对照显著上调,而HbPIX13 表达量则在COR处理前后均无差异(图5B)。
3 讨论
PSKR 是PSK 的直接受体,在PSK 信号传导中起重要作用[6-7] 。本研究克隆了橡胶树的HbPSKR2 基因。序列分析显示,HbPSKR2 是一个跨膜蛋白,含有PSKR 蛋白典型的膜外LRR 结构域和膜内激酶结构域。组织特异性分析显示HbPSKR2 在橡胶树形成层区域相对表达丰度最高,并且在COR 处理早期被显著诱导。课题组前期研究发现茉莉酸信号在诱导形成层分化次生乳管过程中起重要作用[14-15],近期已有研究证明PSK 参与了茉莉酸诱导乳管分化过程[16]。鉴于HbPSKR2 在形成層区域高丰度表达,且可被茉莉酸类似物诱导,因此推测该基因可能参与了橡胶树的乳管分化。
酵母双杂交文库筛选是鉴定互作蛋白的常用方法[17]。本研究构建了HbPSKR2 诱饵表达载体,对橡胶树形成层区域酵母双杂交文库进行筛选,共得到12 个候选蛋白,测序分析显示这些成员涉及初生代谢、生长发育、信号传导等多个生物学途径。蛋白质是生命活动的主要承担者,其生物合成和降解过程直接关系着生命活动的进行[18]。核糖体是真核生物中与蛋白质合成密切相关的细胞器,由不同大小的亚基组成[19]。泛素化是真核生物普遍存在的翻译后修饰方式,通过将靶标蛋白标记泛素标签进而启动蛋白质的降解[20-21]。本研究从HbPSKR2 互作蛋白中发现2 个核糖体相关成员和2 个泛素化相关成员,表明PSK 信号途径可能直接参与橡胶树蛋白质合成和降解过程。
蛋白激酶是一种催化蛋白质发生磷酸化修饰的酶[22]。生物体内的蛋白磷酸化是调控蛋白活力和功能的重要机制之一,在细胞信号传导过程中起重要作用[23-25]。作为一种受体蛋白,已有报道PSKR 可以和多种激酶互作,参与不同的生物学过程。如WANG 等[26]报道拟南芥中PSK 与PSKR互作后可以改变结合位点附近的空间构象,促使PSKR 与体细胞胚胎发育受体激酶(somatic embryogenesisreceptor kinase,SERK)发生互作;HOLZWART 等[9]报道拟南芥的受体蛋白RLP44可以介导油菜素内酯关键成员BRI1-ASSOCIATEDKINASE 1(BAK1)与PSKR1 互作,以复合体的形式介导形成层分化木质部。本研究中通过荧光素酶实验证明2 个蛋白激酶HbPIX13 和HbPBL8 与HbPSKR2 之间存在强烈互作,基因表达模式分析显示HbPBL8 可被COR 显著诱导,推测HbPSKR2-HbPBL8 信号通路可能在橡胶树乳管分化过程中起一定作用。揭示HbPBL8 的靶标蛋白将是下一步的工作重点。
