外源独脚金内酯和丛枝菌根真菌对多枝柽柳抗盐性的影响
2024-04-30徐宁吴亮朱建俊
徐宁 吴亮 朱建俊

摘要:以盐碱地区优势植物多枝柽柳(Tamarix ramosissima Ledeb.)幼苗为研究对象,采用盆栽方式,探究根际施用独脚金内酯并接种丛枝菌根真菌(arbuscular mycorrhizal fungi,简称AMF)对盐胁迫下多枝柽柳生长情况、气体交换参数、叶绿素荧光参数及内源激素变化的影响。结果表明,与非盐处理相比,盐胁迫下多枝柽柳株高减少,叶绿素含量以及光合作用不断降低,PSⅡ光系统中叶绿素荧光参数呈下降趨势,内源激素平衡遭到破坏。而根际施用独脚金内酯能够促进AMF侵染发育,增加泡囊数、丛枝数和侵入点位数,菌根侵染率最高达58.5%。接种AMF和/或独脚金内酯处理均能够增加盐胁迫下多枝柽柳株高和根冠比,提高光合作用,维持PSⅡ光系统中叶绿素荧光参数,盐害指数从65.1%下降到41.5%,多枝柽柳枝条相对含水量也明显增加。盐胁迫下,接种AMF和/或独脚金内酯处理的多枝柽柳叶片净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、胞间二氧化碳浓度(Ci)、气孔导度(Gs)以及气孔限制值(Ls)分别提高20.7%~40.7%、42.6%~176.6%、27.6%~96.6%、38.5%~99.1%、21.2%~75.9%、60.9%~160.9%,叶绿素荧光参数中PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、表观电子传递速率(ETR)分别提高0.2%~4.0%、24.8%~51.0%、37.3%~189.8%,热耗散速率(HDR)下降9.7%~32.3%;内源激素中独脚金内酯(SLs)、生长素(IAA)以及赤霉素(GA)含量分别增加5.1%~5.8%、7.3%~98.8%、31.8%~121.1%,脱落酸(ABA)含量下降20.7%~45.2%。结论认为,接种AMF和/或根施独脚金内酯处理均能提高多枝柽柳叶绿素含量以及枝条相对含水量,增强植物光合作用能力,维持PSⅡ光系统中叶绿素荧光参数以及内源激素含量来增加植物株高和根冠比,显著降低盐害指数,本试验认为根施独脚金内酯并接种AMF是提高多枝柽柳幼苗耐盐性的有效手段。
关键词:丛枝菌根真菌;盐胁迫;独脚金内酯;激素;光合作用;叶绿素荧光参数
中图分类号:S182;S184 文献标志码:A
文章编号:1002-1302(2024)06-0150-09
收稿日期:2023-10-22
基金项目:河南省高校人文社会科学研究一般项目(编号:2024-ZDJH-662);国家自然科学基金(编号:32001100)。
作者简介:徐宁(1987—)女,河南杞县人,硕士,讲师,主要从事园林植物应用及景观设计研究。E-mail:bzfl93771@163.com。
盐胁迫严重影响植物生长发育和繁殖等生命活动,也是人们研究最多的逆境因子之一。盐害限制了我国滨海地区绿化树种的选择范围,影响园林绿化景观效果[1]。独脚金内酯(strigolactones,简称SLs)是一类植物倍半萜类化合物,也是一种新型植物激素,作为植物生长发育所必需的基本调节物质,普遍存在于植物体中,调控着各种植物的生长发育过程[2-3]。此外,独脚金内酯还有助于提高植物对生物和非生物胁迫的抗逆性[4]。研究发现,独脚金内酯在参与调控盐胁迫的过程中,主要通过影响生长素、赤霉素、脱落酸等植物激素的稳态平衡和信号转导途径,从而共同调控植物的生长进程并抵御外界环境胁迫带来的影响,进而适应环境条件[5-6]。有研究发现,盐胁迫条件下喷施独脚金内酯人工合成类似物(GR24)可减轻月季叶片的受害程度,减缓叶绿素的损耗速率,降低相对电导率[7]。王乔健也发现,外施GR24能够提高乌桕幼苗抗盐胁迫的能力,增加乌桕幼苗的生物量,还能够调控氧化还原系统来增加清除自由基和过氧化物的能力[8]。此外,独脚金内酯作为根部来源的信号物质,能够增强植物与丛枝菌根真菌(arbuscular mycorrhizal fungi,简称AMF)之间的共生关系,促进菌丝分枝[9-10]。
丛枝菌根真菌在系统发育上属于球囊霉亚门(Glomeromycotina),能与超过80%的陆生植物根系形成共生关系[11-12]。通过AMF庞大的菌丝网络,寄主植物能够从土壤中获取水分和养分,尤其是土壤磷元素[13]。大量的研究已经证明,AMF能不同程度地提高寄主植物对盐胁迫的耐受力,这种潜在机制包括增强寄主植物水分利用效率、增强光合作用以及活性氧(reactive oxygen species,简称ROS)的清除能力等[14-16]。有研究发现,盐胁迫能显著降低榉树的苗高以及地径净增长量,而接种摩西球囊霉1号(GM1)、2号(GM2)处理显著提高了盐胁迫下榉树的地径净增长量,同时也能够提升榉树叶片净光合速率、气孔限制值、水分利用效率,并降低胞间CO2浓度[17]。崔令军等发现,随盐浓度逐渐增加,接种AMF处理的桢楠根系与对照处理的赤霉素(GA)、生长素(IAA)、玉米素(ZR)含量均呈下降趋势,脱落酸(ABA)含量则呈上升趋势,AMF通过打破激素间的平衡,增加GA、IAA、ZR含量而降低ABA含量来增强桢楠的耐盐性,促进根系生长[18]。
多枝柽柳(Tamarix ramosissima Ledeb.)是柽柳科柽柳属的灌木或小乔木,广泛分布在我国内陆盐碱地区。多枝柽柳具有防风固沙、涵养水源的作用,且能够有效改善盐碱地生态环境。在内陆滨海地区,多枝柽柳是重要的抵抗海岸带侵蚀的优势植物,也是改造盐碱地绿化环境的优良树种,对维持滨海地区生态系统稳定具有重要作用。近年来,对多枝柽柳的研究成果主要集中生长特性、种群分布等方面[19-20],而关于盐胁迫下接种AMF并添加外源独脚金内酯对多枝柽柳幼苗光合以及内源激素的研究更是鲜有报道。鉴于AMF极为广泛的寄主范围和多样化的生理生态功能,在众多有益土壤微生物中地位突出。本研究拟通过温室盆栽试验,探究盐胁迫处理下AMF与根施独脚金内酯二者联合促进多枝柽柳生长特性、光合作用、叶绿素荧光参数以及激素变化的生理机制,对研究盐胁迫下植物生长发育具有重要意义。
1 材料与方法
1.1 试验材料
供试植物材料为多枝柽柳种子,由河南豫艺种业科技发展有限公司提供。独脚金内酯(rac-GR24,纯度≥98%)类似物购买自北京索莱宝科技有限公司,在室温下避光保存。选用的AMF菌种为地表球囊霉(Glomus versiforme)和摩西斗管囊霉(Funneliformis mosseae)混合1∶1组成,购自北京市农林科学院植物营养与资源研究所,接种物主要是栽培基质中的AMF孢子、菌丝和菌根根段,经检测AMF孢子密度260~305个/50 g,菌根侵染率70%。栽培基质选用表层园土(取自河南农业大学校园绿地)和河沙按照体积比1∶1均匀混合后晒干,过2 mm筛,并进行高温高压蒸汽灭菌(121 ℃,0.15 MPa)2 h后备用,经检测栽培基质的pH值为6.78,有机质含量为1.26%,全氮含量为1.35 g/kg,全钾含量为36.5 g/kg,全磷含量为0.89 g/kg,速效磷含量为5.66 mg/kg,速效钾含量为 106.33 mg/kg。
1.2 试验方法
选取均匀度高,大小一致的多枝柽柳种子,用5% NaClO2溶液浸泡消毒5 min,再用蒸馏水冲洗5~6次。随后将消毒后的种子在光照培养箱(PGX-1000B,温度28 ℃恒溫)中催芽,后播种在50孔穴盘中,待幼苗长至5~7 cm高度时上盆,每盆栽植1株(盆高为30 cm,上口直径为30 cm,下口直径为25 cm),盆内装基质3.5 kg。
试验于2022年4—10月在河南农业大学空地进行,采用盆栽试验。试验共设置8个处理:(1)CK,不添加盐处理,(2)AMF(接种地表球囊霉和摩西斗管囊霉混合1∶1菌种),(3)SLs(根施 50 nmol/L 独脚金内酯),(4)AMF+SLs(接种混合AMF并喷施50 nmol/L独脚金内酯),(5)SS(进行0.6%盐浓度处理),(6)SS+AMF(0.6%盐浓度处理下混合接种AMF),(7)SS+SLs(0.6%盐浓度处理下根施 50 nmol/L 独脚金内酯),(8)SS+AMF+SLs(0.6%盐浓度处理下混合接种AMF并根施 50 nmol/L 独脚金内酯),每个处理6个重复。研究发现,多枝柽柳幼苗在土壤盐分为0.5%~0.8%间出现叶片变黄的现象[21],故本次试验盐浓度设置为0.6%。接种AMF处理的接种物数量为100 g/盆,对照(CK)则接种等量灭菌接种物(121 ℃,20 min下高温高压灭菌),以保持相同的其他根围微生物区系环境。施用独脚金内酯的处理用蒸馏水配成 50 nmol/L 的溶液进行灌根(浓度参照范吴蔚等的研究[22]设定),每间隔7 d灌根1次,其余组施用等量去离子水灌根。待多枝柽柳移苗生长25 d后进行盐处理,每隔3 d 加入1次盐溶液,使盐含量达到栽培基质质量的0.6%。管理期间保证温度、水分、养分、通风等条件的控制,每周定期随机调换盆栽位置,确保条件的一致性,观察记录各处理组多枝柽柳生长情况,盐胁迫70 d后收获并测量各项指标。
1.3 指标测定
1.3.1 AMF侵染指标的测定
随机选取10株多枝柽柳,将根系用水冲洗干净,洗净后加入10%氢氧化钾溶液,在90 ℃水浴锅(DZKW-S-6型)中放置30 min,冷却后清洗干净,后加1%HCl酸化,加0.1%酸性品红-乳酸甘油染色液放置12 h,在显微镜下制片镜检,根据刘润进等的描述方法计算菌根侵染率、泡囊数、丛枝数以及侵入点位数[23]。
1.3.2 生长指标的测定
用直尺测量多枝柽柳株高,将植株的根、茎、叶分别进行收获。注意收获根部分时,小心去除根部杂质,尽量不要损伤根部,用水将杂质冲洗干净。分别将根、茎、叶放入85 ℃烘箱中烘干至恒重,称取各部分重量并记录。根冠比=根生物量/地上部生物量。
1.3.3 盐害指数的测定
盐害分级的方法参照付晴晴等的分类,在测定各项指标前2 d测定[24]。
1.3.4 叶绿素含量以及枝相对含水量的测定
采用乙醇提取法提取叶绿素,用紫外可见分光光度计分别在波长663、645 nm下测定吸光度,计算叶绿素含量;采用饱和称重法测定枝相对含水量,枝相对含水量=(枝鲜重-枝干重)/(枝饱和重-枝干重)×100%。
1.3.5 气体交换参数的测定
选择晴朗少云的天气,采用美国CIRAS-3便携式光合仪,在08:30—11:30进行测定。每个处理选取6~8株植株,每株植株选取1张中上部健康的、完全展开的成熟叶进行测量。测量过程中记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、水分利用效率(WUE)以及气孔限制值(Ls)。
1.3.6 叶绿素荧光测定
选取晴朗少云的天气,测定时间为08:30—11:30,采用Pocket PEA植物效率仪进行叶片的叶绿素荧光参数测定。测量的叶片与气体交换测量选择的叶片相同。叶片暗适应 30 min 后,测定PSⅡ最大光化学效率( Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、表观电子传递速率(ETR)、热耗散速率(HDR)等参数,所有测定重复3次,取平均值。
1.3.7 内源激素含量测定
内源激素含量的测定采用酶联免疫吸附测定法(enzyme-linked immunosorbent assay,简称ELISA),独角金内酯、生长素、赤霉素以及脱落酸试剂盒均购买于武汉吉立德生物科技有限公司,各项激素均重复3次测量。
1.4 数据处理
采用Microsoft Excel 2010软件对数据进行处理和绘图,所有试验均采用SPSS 16.0进行统计分析,通过单因素方差分析(one-way ANOVA)双因素方差分析(two-way ANOVA)以及差异显著性检验(LSD法,α=0.05)处理数据。表中数据为平均值±标准误。
2 结果与分析
2.1 盐胁迫下多枝柽柳菌根发育情况
本试验条件下,AMF能够侵染多枝柽柳根系,并形成稳定的菌根共生关系。而盐胁迫下AMF的菌根侵染率、根内泡囊数、丛枝数和根上菌丝侵入点数均有所降低,说明盐胁迫能抑制AMF对多枝柽柳根系的侵染。非盐胁迫或者盐胁迫下,根施独脚金内酯有助于促进AMF的侵染(表1)。与AMF处理相比,AMF+SLs处理下的菌根侵染率、根内泡囊数、丛枝数和根上菌丝侵入点数分别提高18.2%、17.2%、23.0%和18.2%;盐胁迫下,与SS+AMF处理相比,SS+AFM+SLs处理下的菌根侵染率、根内泡囊数、丛枝数和根上菌丝侵入点数分别提高10.0%、31.7%、56.1%和15.6%。可知,非盐胁迫或者盐胁迫下,根施SLs处理均能促进AMF发育。
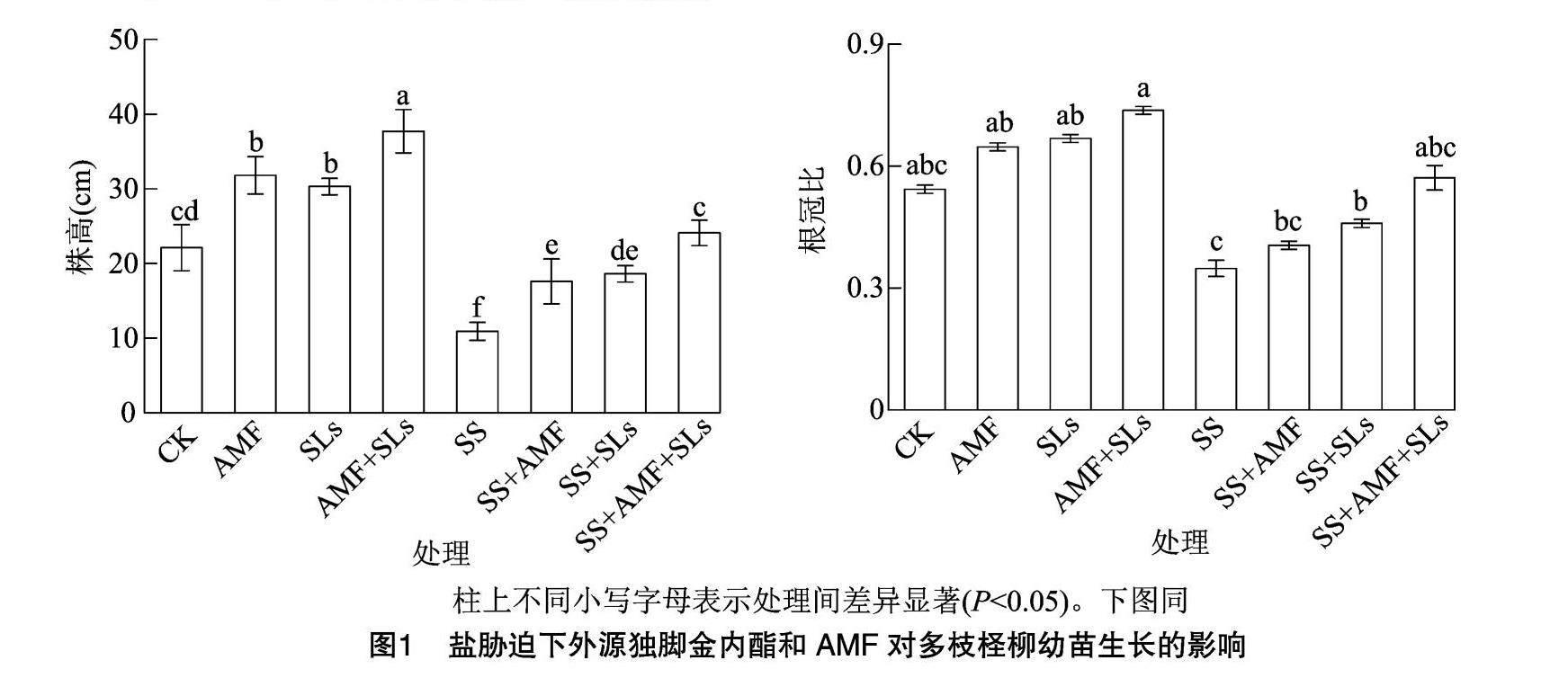
2.2 盐胁迫下外源独脚金内酯和AMF对多枝柽柳生长的影响
由图1可知,盐胁迫下,多枝柽柳的株高和根冠比均呈下降趋势,非盐胁迫或者盐胁迫下接种AMF或者根施SLs均有助于促进多枝柽柳的生长。非盐胁迫下,与CK处理相比,AMF处理下的多枝柽柳的株高和根冠比增加43.9%和19.2%,SLs处理下分别增加37.1%和23.0%,AMF+SLs处理分别增加70.6%和35.7%,AMF与SLs处理下的多枝柽柳株高和根冠比差异均不显著。盐胁迫下,与SS处理相比,SS+AMF处理下的多枝柽柳株高和根冠比分别提高61.5%、16.4%,SS+SLs处理下的株高和根冠比分别提高70.6%、31.9%,SS+AMF+SLs处理下分别提高121.1%、64.1%。可见,非盐胁迫或者盐胁迫下,接种AMF并根施SLs促进多枝柽柳生长的效果优于单一AMF处理或者SLs处理,且增幅效果在盐胁迫下更大。
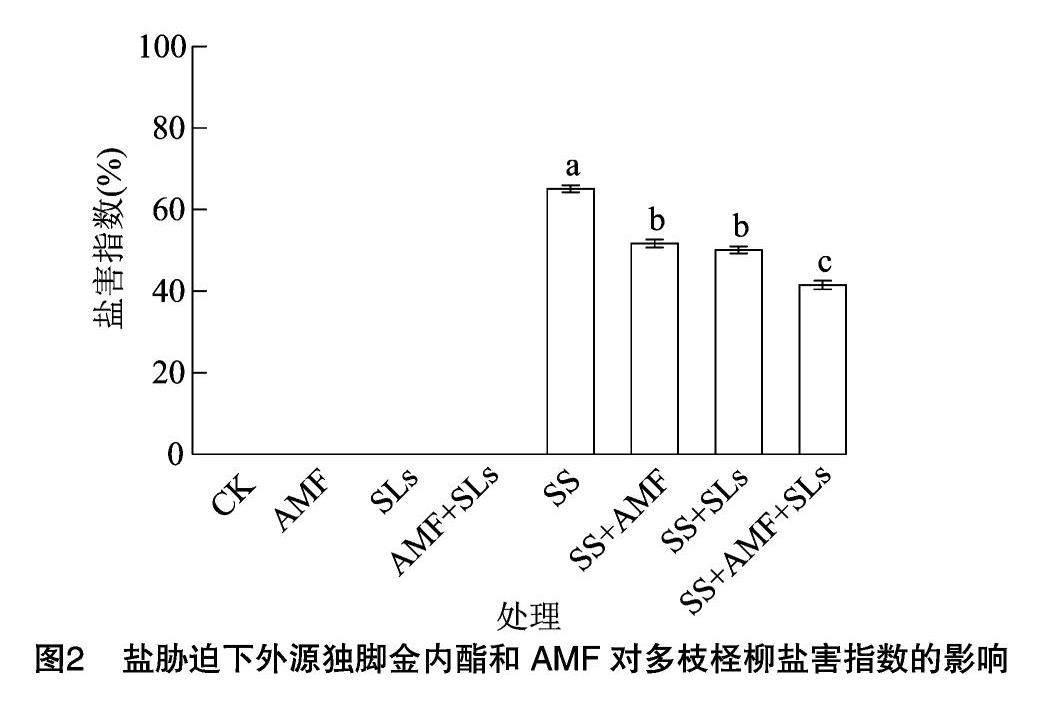
2.3 盐胁迫下外源独脚金内酯和AMF对多枝柽柳盐害指数的影响
盐胁迫处理的多枝柽柳幼苗叶片表现出叶缘黄化并萎蔫的现象,植株生长较为缓慢,而非盐胁迫处理下的多枝柽柳幼苗生长较为健壮,并未出现盐害症状。盐胁迫下,SS处理的多枝柽柳幼苗盐害指数为65.1%,SS+AMF处理的盐害指数为51.7%,SS+SLs处理的盐害指数为50.1%,而SS+AMF+SLs处理下的盐害指数为41.5%(图2)。可见,盐胁迫下接种AMF或者根施SLs均可以缓解盐害,但SS+AMF与SS+SLs处理差异不显著。
2.4 盐胁迫下外源独脚金内酯和AMF对多枝柽柳叶绿素和枝条含水量的影响
由图3可知,盐胁迫下,多枝柽柳叶片叶绿素含量和枝相对含水量表现为下降的趋势,而非盐胁迫或者盐胁迫下接种AMF或者根施SLs均有助于多枝柽柳叶片积累叶绿素含量以及枝条含水量。非盐处理下,与CK处理相比,AMF处理多枝柽柳的叶绿素含量44.7%,枝相对含水量无显著差异,SLs处理下的叶绿素含量增加34.9%,SLs处理下的枝相对含水量与CK处理无显著差异,AMF+SLs处理的叶绿素含量和枝相对含水量分别提高82.5%和6.1%。盐胁迫下,与SS处理相比,SS+AMF处理下的多枝柽柳叶绿素含量和枝相对含水量分别提高33.9%和2.3%,SS+AMF+BRs处理下叶绿素含量和枝相对含水量分别提高86.4%和5.0%,但SS+SLs处理下的叶绿素含量和枝相对含水量与SS处理无显著差异,其他处理下差异均显著。可见,非盐胁迫或者盐胁迫下,接种AMF并根施SLs提高多枝柽柳叶绿素含量以及枝相对含水量的效果优于单一AMF处理或者SLs处理,对叶绿素含量的增幅效果在盐胁迫下更大一些。
2.5 盐胁迫下外源独脚金内酯和AMF对多枝柽柳气体交换参数的影响
由图4可知,盐胁迫下,多枝柽柳净光合速率、蒸腾速率、水分利用效率、胞间二氧化碳浓度、气孔导度以及气孔限制值的变化趋势基本一致,均呈现下降的趋势;AMF或者SLs处理多枝柽柳幼苗叶片的Pn、Tr、WUE、Ci、Gs及Ls显著高于CK处理。非盐胁迫下,与CK处理相比,AMF处理下多枝柽柳的Pn、Tr、WUE、Ci、Gs及Ls分别提高12.6%、103.9%、48.6%、48.7%、40.5%和110.0%,SLs处理下各指标分别增加4.2%、55.3%、22.9%、27.2%、24.3%和66.7%,AMF+SLs处理下各指标分别增加16.8%、181.6%、94.3%、81.3%、53.3%和176.7%,但AMF处理与AMF+SLs处理下的Pn差异不显著。盐胁迫下,与SS处理相比,SS+AMF處理下多枝柽柳的Pn、Tr、WUE、Ci、Gs及Ls分别提高21.3%、97.9%、62.1%、60.8%、40.2%和117.4%,SS+SLs处理下各指标分别增加20.7%、42.6%、27.6%、38.5%、21.2%和60.9%,SS+AMF+SLs处理下各指标分别增加40.7%、176.6%、96.6%、99.1%、75.9%和160.9%,但 SS+AMF处理与SS+SLs处理下的Pn无显著差异。可见,非盐胁迫或者盐胁迫下,接种AMF并根施SLs处理对多枝柽柳光合作用的提升效果优于单一处理。
2.6 盐胁迫下外源独脚金内酯和AMF对多枝柽柳叶绿素荧光参数的影响
由图5可知,盐胁迫下,多枝柽柳叶片中PSⅡ最大光化学效率、PSⅡ潜在活性、表观电子传递速率下降,而热耗散速率则表现为增加的趋势,接种AMF处理或者SLs处理下能够提高多枝柽柳 Fv/Fm、Fv/Fo以及ETR等参数,而降低HDR参数。非盐胁迫下,与CK处理相比,AMF处理下多枝柽柳的Fv/Fm和Fv/Fo分别增加2.6%和31.8%,ETR下降,HDR下降47.6%,SLs处理下的Fv/Fm值与CK处理差异不显著,SLs处理下的HDR与CK处理相比下降14.3%,AMF+SLs处理下的Fv/Fm、Fv/Fo以及ETR分别增加6.5%、44.5%、33.9%,HDR下降61.9%。盐胁迫下,与SS处理相比,SS+AMF处理下多枝柽柳的Fv/Fm、Fv/Fo、ETR分别增加1.3%、33.1%和116.8%,HDR下降25.8%,SS+SLs处理下Fv/Fm、Fv/Fo、ETR分别增加0.2%、24.8%和37.3%,HDR下降9.7%,SS+AMF+SLs处理下的Fv/Fm、Fv/Fo以及ETR分别增加4.0%、51.0%、189.8%,HDR下降32.3%。可见,非盐胁迫或者盐胁迫下,接种AMF并根施SLs处理对多枝柽柳Fv/Fm、Fv/Fo以及ETR等参数的提升效果优于单一处理,降低HDR的效果也高于单一处理。
2.7 盐胁迫下外源独脚金内酯和AMF对多枝柽柳内源激素含量的影响
盐胁迫下,多枝柽柳叶片中独脚金内酯、生长素以及赤霉素含量下降,而脱落酸含量上升,接种AMF处理和/或SLs处理均能够提高多枝柽柳叶片中内源SLs、IAA以及GA含量,而降低ABA含量。非盐胁迫下,与CK处理相比,AMF处理下多枝柽柳叶片中的内源SLs稍下降,IAA以及GA含量分别增加95.2%和116.1%,ABA含量下降33.6%;SLs处理下内源SLs、IAA以及GA含量分别增加3.6%、48.5%和33.5%,ABA含量下降28.2%;AMF+SLs处理下内源SLs、IAA以及GA含量分别增加7.7%、189.6%和125.9%,ABA含量下降46.4%。盐胁迫下,与SS处理相比,SS+AMF处理下多枝柽柳叶片中的内源SLs下降,IAA以及GA含量分别增加45.4%和65.8%,ABA含量下降26.1%;SS+SLs
处理下内源SLs、IAA以及GA含量分别增加5.1%、7.3%和31.8%,ABA含量下降20.7%;SS+AMF+SLs处理下内源SLs、IAA以及GA含量分别增加5.8%、98.8%和121.1%,ABA含量下降45.2%。可见,非盐胁迫或者盐胁迫下,接种AMF并根施SLs处理对内源激素的调控作用优于单一处理。
3 讨论
土壤盐含量直接影响植物干物质的积累和生长发育,随着土壤盐含量增加,植物生长发育状况也会随之降低。有研究发现,盐胁迫下,AMF可以主动向植物运输水分和养分,并且形成菌根后根系的输水效率更高,水分运输阻力更小,同时能够保护寄主植物的光合作用,调节气孔导度以提高水分利用率等方式来提高植物的抗鹽能力[25-26]。目前已有研究表明,独脚金内酯在植物中不仅可作为一种信号分子调控植物生长发育以及株型的形成,还能抵御各种生物和非生物胁迫,增强植物抗逆性[27]。独脚金内酯不仅可以调控植物的分枝能力,还参与植物根系的建成,通过对黄芪、苹果等植物的研究发现,独脚金内酯能够抑制侧根的生长,而促进主根的生长发育,调控根系构型来改善植物营养[28-29]。此外,独脚金内酯对菌根共生也有积极的影响,作为植物与根际微生物之间的交流信号,能够增加丛枝菌根真菌菌丝的分枝数量,特别是营养元素缺乏的土壤中,可加速刺激植物合成活性更强
的独脚金内酯并快速释放到土壤当中,改善土壤中氮磷钾的平衡状态,促进寄主植物生长[30-31]。本试验也证明,盐胁迫下,AMF的侵染情况下降,独脚金内酯能促进AMF的生长发育,增加丛枝数和根上菌丝侵入点数,帮助其侵染多枝柽柳并形成稳定的共生关系,显著增加多枝柽柳的株高,并形成更高的根冠比来增加根系的生物量。盐胁迫不仅影响AMF的繁殖,还会使寄主植物出现盐害等现象,接种AMF或者独脚金内酯根施后可以使多枝柽柳的幼苗盐害情况减弱,说明二者可以提高多枝柽柳幼苗的耐盐性。
植物通过光合作用将大气中的CO2固定转化为有机营养物质,在物质循环以及能量流动过程中被利用。有研究表明,盐胁迫处理会影响植物的光合作用,主要表现在碳同化效率的降低以及有机物合成能力的下降[32]。盐浓度50 mmol/L下对植株幼苗的净光合速率影响不大,超过50 mmol/L后Pn开始下降,气孔限制值也减小[33]。接种AMF可以维持植物正常的生长发育状态,改善植物光合能力[34]。本试验中,盐胁迫下多枝柽柳叶片叶绿素含量和枝相对含水量表现为下降的趋势,而接种AMF或根施独脚金内酯后,叶绿素含量均有增加的趋势,说明AMF或根施独脚金内酯能够增加叶绿素在细胞内合成的能力,显著提高植物的光和效率。叶绿素荧光参数在植物进行光合作用过程中也发挥着重要作用,其中Fv/Fm、Fv/Fo、ETR和HDR是 PSⅡ 反应系统的重要参数,可反映环境胁迫对植物光合作用的影响[35]。本试验发现,Fv/Fm、Fv/Fo、ETR在盐处理下表现为下降的趋势,HDR增加,接种AMF或根施独脚金内酯后,表现则相反。说明AMF或独脚金内酯可以有效减缓盐浓度处理带来的光抑制现象,提高PSⅡ反应系统对原初光能的捕获效率,从而使光合器官的破坏程度变小,光合效率进一步增加。净光合速率、蒸腾速率、气孔导度、水分利用率、胞间CO2浓度以及气孔限制值等是反映植物光合作用的主要参数,包括植物碳同化以及有机物的合成能力。本试验条件下,盐胁迫显著降低了植物光合作用,各参数值整体显著下降;接种AMF或者根施独脚金内酯后,光合作用显著改善,净光合速率增加,这与梁倩倩等的研究结果一致,其发现盐处理下接种摩西球囊霉的牡丹植株叶片净光合速率、气孔导度、胞间CO2利用能力、蒸腾速率、水分利用效率和植株生长量均显著高于不接种对照,AMF能显著提高牡丹的光合作用并提高耐盐性[36]。而独脚金内酯可能通过促进多枝柽柳根系的生长来发挥作用,使得根冠比增加,具体还需进一步试验验证。
植物激素是对植物的生长发育有显著调节作用的微量有机物,在植物耐盐性方面发挥着重要作用,这些激素相互作用,调节生物化学和生理过程,转化为生长、发育、营养分配作用[37-38]。本研究发现盐处理下,丛枝菌根真菌可以促进寄主植物对IAA、GA的积累,而降低ABA的产生量。菌根植物能够合成独脚金内酯,促进根部真菌的生长,而菌根植物依靠围绕在根部的丛枝真菌从土壤中吸收氮磷钾等营养物质,丛枝真菌也从植物根部得到所需的糖类等[39]。本试验发现,盐胁迫显著降低了IAA、GA以及SLs的含量,但接种AMF后GA、IAA含量显著增加。另有研究证明,独脚金内酯能够调节其他植物激素的含量,特别是IAA和ABA,并与之相互作用[40]。本试验条件下,盐胁迫后ABA含量显著高于对应非盐胁迫处理,而接种AMF后ABA含量表现为降低的趋势,这表明在盐处理下接种AMF可以减少ABA的合成水平,促进寄主植物生长发育。因此,多枝柽柳接种AMF或者SLs处理后可通过积累IAA、GA含量,降低ABA的产生量来减缓盐胁迫造成的伤害。
4 结论
综上所述,盐胁迫下多枝柽柳生长受抑制,株高和根冠比下降,叶绿素含量以及枝条相对含水量降低,光合作用下降。而根施独脚金内酯能够促进AMF对多枝柽柳的侵染,增加泡囊数、丛枝数和侵入点位数。独脚金内酯配合AMF接种能增加叶绿素含量,通过提高叶片水分利用效率、净光合速率、蒸腾速率和气孔导度来增强植物光合作用能力,维持PSⅡ光系统中最大光化学效率、PSⅡ潜在活性以及表观电子传递速率,而减低热耗散速率,显著增加植株的株高和根冠比,维持体内激素平衡,来降低多枝柽柳的盐害指数,结论认为根施独脚金内酯配合AMF接种是提高滨海地区多枝柽柳幼苗抗盐性的有效手段。
参考文献:
[1]王文卿,刘俊伟,王良睦. 重视南方滨海地区城镇园林绿化树种盐害的研究[J]. 中国园林,2000,16(5):73-75.
[2]Xu E J,Chai L,Zhang S Q,et al. Catabolism of strigolactones by a carboxylesterase[J]. Nature Plants,2021,7:1495-1504.
[3]吴转娣,胡鑫,昝逢刚,等. 独脚金内酯的生物合成及其调控[J]. 中国农业大学学报,2020,25(12):97-110.
[4]Wang L,Wang B,Yu H,et al. Transcriptional regulation of strigolactone signalling in Arabidopsis[J]. Nature,2020,583:277-281.
[5]Sun H W,Li W Q,Burritt D J,et al. Strigolactones interact with other phytohormones to modulate plant root growth and development[J]. The Crop Journal,2022,10(6):1517-1527.
[6]沈月,陶寶杰,华夏,等. 独脚金内酯与激素互作调控根系生长的研究进展[J]. 生物技术通报,2022,38(8):24-31.
[7]朱思雅,段建维,许彬,等. 独角金内酯对盐胁迫下月季植株生长的影响[J]. 现代园艺,2022,45(1):13-15,20.
[8]王乔健. 独脚金内酯调控乌桕抗旱耐盐的分子机理研究[D]. 合肥:安徽农业大学,2019.
[9]Akiyama K,Matsuzaki K I,Hayashi H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi[J]. Nature,2005,435(7043):824-827.
[10]Müller L M,Flokova K,Schnabel E,et al. A CLE–SUNN module regulates strigolactone content and fungal colonization in arbuscular mycorrhiza[J]. Nature Plants,2019,5:933-939.
[11]谢伟,郝志鹏,张莘,等. 丛枝菌根网络介导的植物间信号交流研究进展及展望[J]. 植物生态学报,2022,46(5):493-515.
[12]Chaturvedi A,Cruz Corella J,Robbins C,et al. The methylome of the model arbuscular mycorrhizal fungus,Rhizophagus irregularis,shares characteristics with early diverging fungi and Dikarya[J]. Communications Biology,2021,4:901.
[13]雷梅,甘子莹,谭世广,等. 丛枝菌根真菌和不同形态氮对杉木幼苗根际土壤氮磷养分含量及其相关酶化学计量比的影响[J]. 林业科学研究,2023,36(1):59-67.
[14]周昕南,杨亮,许静,等. 接种AM真菌对不同盐度土壤中向日葵生长的影响[J]. 农业资源与环境学报,2020,37(5):744-752.
[15]贾婷婷,常伟,范晓旭,等. 盐胁迫下AM真菌对沙枣苗木光合与叶绿素荧光特性的影响[J]. 生态学报,2018,38(4):1337-1347.
[16]吴强盛,柳威,翟华芬,等. 盐胁迫下AM真菌对枳实生苗生长和根系抗氧化酶的影响[J]. 江西农业大学学报,2010,32(4):759-762,782.
[17]马仕林,曹鹏翔,张金池,等. 盐胁迫下AMF对榉树幼苗生长和光合特性的影响[J]. 南京林业大学学报(自然科学版),2022,46(1):122-130.
[18]崔令军,刘瑜霞,林健,等. 盐胁迫下丛枝菌根真菌对桢楠根系生长和激素的影响[J]. 南京林业大学学报(自然科学版),2020,44(4):119-124.
[19]肖生春,肖洪浪,司建华,等. 干旱区多枝柽柳的生长特性[J]. 西北植物学报,2005,25(5):1012-1016.
[20]赵峰侠,尹林克. 荒漠内陆河岸胡杨和多枝柽柳幼苗种群空间分布格局及种间关联性[J]. 生态学杂志,2007,26(7):972-977.
[21]张瑞群,马晓东,吕豪豪. 多枝柽柳幼苗生长及其根系解剖结构对水盐胁迫的响应[J]. 草业科学,2016,33(6):1164-1173.
[22]范吴蔚,潘志演,王原秀,等. 独脚金内酯对烟草腋芽生长以及抗氧化酶活性和糖含量的影响[J]. 植物生理学报,2021,57(4):910-918.
[23]刘润进,陈应龙. 菌根学[M]. 北京:科学出版社,2007.
[24]付晴晴,孙鲁龙,翟衡,等. 葡萄种间杂交砧木育种F1代植株耐盐性分析[J]. 植物学报,2017,52(6):733-743.
[25]郭娜,张玥,刘贤雍,等. 丛枝菌根真菌提高植物耐盐性生理机制研究进展[J]. 江苏农业科学,2023,51(4):16-23.
[26]陈晓楠,伊力努尔·艾力,高文礼,等. 盐胁迫下丛枝菌根真菌对疏叶骆驼刺幼苗生长和生理的影响[J]. 草业科学,2022,39(9):1763-1772.
[27]王玫,陈洪伟,王红利,等. 独脚金内酯调控植物分枝的研究进展[J]. 园艺学报,2014,41(9):1924-1934.
[28]赵玉欢,黄晓宇,关环,等. 独脚金内酯对黄芪幼苗根系发育的影响[J]. 植物生理学报,2023,59(3):505-514.
[29]贾竣淇. 外源独脚金内酯及其抑制剂对苹果根系生长发育和养分吸收的影响[D]. 泰安:山东农业大学,2022.
[30]Ha C V,Leyva-González M A,Osakabe Y,et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress[J]. Proceedings of the National Academy of Sciences of the United States of America,2014,111(2):851-856.
[31]Mostofa M G,Li W Q,Nguyen K H,et al. Strigolactones in plant adaptation to abiotic stresses:an emerging avenue of plant research[J]. Plant,Cell & Environment,2018,41(10):2227-2243.
[32]王宇超,王得祥. 鹽胁迫对木本滨藜叶绿素合成及净光合速率的影响[J]. 农业工程学报,2012,28(10):151-158.
[33]王耘,金潇潇,赵辉,等. 盐胁迫对鲜食蚕豆幼苗光合特性及抗氧化酶活性的影响[J]. 北方园艺,2017(20):25-30.
[34]王志刚,毕银丽,李强,等. 接种AM真菌对采煤沉陷地复垦植物光合作用和抗逆性的影响[J]. 南方农业学报,2017,48(5):800-805.
[35]李瑞,周玮,陆巍. 低叶绿素b水稻叶片自然衰老过程中光合作用与叶绿素荧光参数的变化[J]. 南京农业大学学报,2009,32(2):10-14.
[36]梁倩倩,王维华,郭绍霞,等. 盐胁迫下丛枝菌根对牡丹光合作用的影响[J]. 青岛农业大学学报(自然科学版),2013,30(2):79-83.
[37]杨瑞瑞,李小双,梁玉青,等. 植物激素在苔藓生长发育与逆境响应过程中的作用机制研究进展[J]. 西北植物学报,2022,42(3):527-540.
[38]白凤麟,樊雨荷,李琳. 硒、硅对盐胁迫下小麦光合生理及养分累积的影响[J]. 江苏农业科学,2023,51(3):68-75.
[39]孙金华,毕银丽,裘浪,等. 土壤中丛枝菌根真菌对宿主植物磷吸收作用机制综述[J]. 土壤通报,2016,47(2):499-504.
[40]高雨,李颖,谢寅峰,等. 独脚金内酯调控植物侧枝发育的分子机制及其与生长素交互作用的研究进展[J]. 植物资源与环境学报,2013,22(4):98-104.
