基于代谢组学解析‘翠碧一号’烟叶烘烤过程中多酚类代谢物动态变化
2024-04-24金辽刘银松林建枫张文伟顾钢魏恕文汪睿琪谢小芳张炳辉
金辽,刘银松,林建枫,张文伟,顾钢,魏恕文,汪睿琪,谢小芳,4,张炳辉
(1福建省烟草公司南平市公司,福建延平 353000;2福建农林大学生命科学学院,福州 350002;3福建省烟草专卖局烟草科学研究所,福州 350003;4福建省作物设计育种重点实验室,福州 350002)
0 引言
烘烤是烟叶感官和品质形成的关键阶段。目前烟叶生产主要以三段式烘烤为主,即变黄、定色、干筋。其中,变黄阶段主要使烟叶变黄变软,包含了一系列物质转化过程,如淀粉、蛋白质水解,可溶性糖、有机酸增多,以及氨杂味散失,烟碱含量降低等;定色阶段使烟叶干燥,固定并保持已形成的色泽和品质;干筋阶段则是使整个叶片及主脉干燥。在烘烤过程中,烟叶经历了一系列复杂的物质代谢变化,其代谢产物随烘烤温度和时间的动态变化组成了烟叶特有的内含物质,形成了特色的香气和风味。
烟草中含有各种多酚类物质及其相关化合物,包括绿原酸、香豆素类、黄酮类等[1-2]。多酚类物质在烟草的调制特性,色泽和等级,以及烟气香味等各个方面起着重要作用[3-4]。烘烤过程中的酶促棕色化反应是酚类物质转化的主要途径,适当的棕色化反应能够使酚类物质有效转换,是生产高品质烟叶产品的关键工艺。以往对酶促棕色化反应的研究多集中在PPO 活性变化[5]、单一或几种化学成分,如多酚类物质、萜类、生物碱等含量[6-8],忽视了对代谢产物的整体探究,特别是烘烤过程的棕色化反应过程中多酚代谢途径的整体变化,以及多酚类物质降解产物等方面还有众多问题尚不清楚。
代谢组学(Metabolomics)利用高通量检测与数据处理,以信息建模和系统整合为目标,研究特定时期和特定环境中生物体或某一器官组织中全部小分子的代谢产物整体变化规律和机制,包括次生代谢组,挥发性代谢组,以及糖醇类化合物、氨基酸、有机酸类、脂类等物质进行定性和定量分析[9],阐明代谢途径中关键的酶和底物与中间产物的相互作用情况,为构建新的代谢通路提供依据[10]。在烟草烘烤过程中,利用代谢组学方法分析烟叶品质形成的各类代谢产物的动态变化及其之间的相互作用关系,可以从整体上探究物质的代谢变化,揭示烟草品质形成的代谢物质基础[11]。福建省烟区特色品种翠碧一号(CB-1)烟叶具有较高的内在品质,在卷烟工业的烟叶配方中的使用性较高[12],但是其烟叶较难烘烤,常常发生过度的棕色化反应而产生较高比例的挂灰烟,影响了经济效益[13-14]。本研究拟以CB-1为材料,利用代谢组学技术方法从全局上分析烟叶烘烤过程中的代谢物变化,系统探究烘烤过程中多酚类物质代谢及其相关代谢通路,以期从整体上解析烘烤过程中多酚类代谢产物的动态变化,为揭示烟草品质形成的代谢物质基础和棕色化反应调控奠定理论基础。
1 材料和方法
1.1 试验材料
以福建省南平市延平区巨口乡栽种的烤烟品种翠碧一号(CB-1)上部叶(13~15 叶位)为实验材料。依据当地烘烤工艺,选取烘烤过程中5 个关键节点的烟叶样品,包括变黄前期(40℃)和变黄后期(41℃)、以及定色前期(43℃)、定色期(45℃)和46℃(定色小卷筒期)等,分别记为T1、T2、T3、T4和T5。每6 片烟叶作为一个样品,3个生物学重复,共15份样品,剪去叶片前后1/4部分后,沿主脉两侧打孔取样,锡纸包裹后做好标记,置于液氮中,随后放入-80℃保存备用。
1.2 样品处理
将采集的样品放置于冻干机中进行真空冷冻干燥,进一步利用研磨仪将样品研磨至粉末状,称取50 mg 的粉末,溶解于1.2 mL 70%甲醇提取液中,充分涡旋和离心后,吸取上清,用微孔滤膜(型号为0.22 μm pore size)过滤,并将样品置于进样瓶中,用于代谢物分析。
1.3 仪器分析
利用超高液相色谱串联质谱(UPLC-MS/MS)检测分析烘烤过程中5个关键节点的样品代谢物。主要仪器为超高液相色谱(UPLC,ExionLC™AD),串联质谱(Tandem mass spectrometry, MS/MS, Applied Biosystems 6500 QTRAP),色谱柱(Agilent SB-C18,1.8 μm,2.1 mm×10 mm)。试验所用的试剂均为色谱纯,主要包括甲醇、甲酸和乙腈。具体的检测由武汉迈特维尔生物科技有限公司完成(www.metware.cn)。
1.4 数据分析
基于迈特维尔生物科技有限公司的自建MWDB V2.0数据库,根据二级谱信息进行物质定性。利用三重四级杆质谱的多反应监测模式对不同烘烤温度节点样品的代谢物进行定量分析,获得各样品的代谢物质谱分析数据,进一步对所有物质色谱峰进行峰面积积分,并对其中同一代谢物在不同样本中的质谱出峰进行积分校正[15]。通过混合样本提取物制备质控样本,用于监测和校正代谢物提取和检测的重复性和可靠性。采用软件Analyst 1.6.3 处理质谱数据。利用R 语言(www.r-project.org)进行主成分分析(PCA),显示样品组内和组间的差异。以|Log2FC|≥1,VIP≥1,P<0.05为阈值筛选差异代谢物。对鉴定的差异代谢物的相对含量进行标准化处理,利用K均值进行聚类分析。利用KEGG 数据库对代谢物进行注释[16],分析样品间差异代谢物的KEGG 代谢通路(http://www.kegg.jp/kegg/pathway.html)。
2 结果与分析
2.1 样本主成分分析
15份样本和QC样本的主成分分析(PCA)显示(图1A),QC 样本点聚集在中心区域,说明这些样本分析方法稳定,数据质量良好。从15 份样本的PCA 图(图1B)发现,第1 成分(PC1)对样本的贡献率达43.98%,第2成分(PC2)对样本的贡献率为8.48%。每个处理内部的样本点分布相对集中,表明组内样本的重复性较好,代谢物较一致。此外,变黄前期(40℃)和变黄后期(41℃)的样本与其他样本的分布明显分离,但是43℃、45℃和46℃等3 个温度节点样本分布较为集中,说明从变黄前期(40℃)至定色前期(43℃),烟叶的物质代谢明显,代谢物之间存在较大差异,而定色期(43℃)之后,叶片的物质代谢较为缓慢,样本间的差异较小。
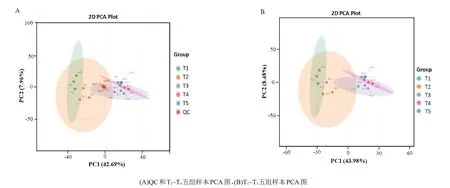
图1 样本代谢组的主成分分析
2.2 不同温度节点样本差异代谢物
为了揭示烘烤过程中物质代谢变化,对40℃、41℃、43℃、45℃和46℃(即T1~T5)等5 个温度节点的代谢物进行检测分析。在5组样本中共鉴定了1069种代谢物,主要为黄酮类、酚酸类,脂质、生物碱、氨基酸及其衍生物、有机酸、核苷酸及其衍生物、木脂素和香豆素、萜类等物质,其中黄酮类和酚酸类等多酚类物质为374 种,达总数的34.99%。通过两两比较发现(图2),烘烤过程中的T1和T2节点与其他温度节点的差异代谢物较多,并且主要以上调为主。其中,以T1为对照的4 组比较组(T1VS T2,T1VS T3,T1VS T4,T1VS T5)的差异代谢物分别为137(上调130,下调7)、331(上调298,下调33)、339(上调304,下调35)、322(上调294,下调28),以T2为对照的3 组比较(T2VS T3,T2VS T4,T2VS T5)的差异代谢物分别为262(上调246,下调16)、284(上调261,下调23)、289(上调255,下调34)。同时,T3、T4和T5等3 个温度点之间的差异代谢物很少,T3VS T4、T3VS T5和T4VS T5等比较组的差异代谢物都仅分别为13,14,19,说明烘烤定色(43℃)之后,烟叶内物质的代谢速度下降,物质含量变化幅度较小。此外,在所有的7个比较组中,多酚类差异代谢物为101种。在T1VS T2、T1VS T3、T1VS T4、T1VS T5等4个比较组多酚类的差异代谢物分别29,72,73和69(图3),其中有24 种多酚类差异代谢物为这4 个比较组(T1VS T2、T1VS T3、T1VS T4、T1VS T5)所共有,达总数的27.59%,有35 种差异代谢物为3 组比较组所共有(T1VS T3,T1VS T4,T1VS T5),达总数的40.23%。

图2 各个比较组中的差异代谢物数目

图3 各个比较组中多酚类差异代谢物Venn图
2.3 多酚类差异代谢物K均值聚类
为探究代谢物在不同样本组中的相对含量变化趋势,对多酚类差异代谢物进行了K均值(K-Means)聚类分析,结果发现,101 种差异代谢物聚为8 种类型(图4),每种模式中的代谢物具有类似的变化趋势。其中,Class 5 包含的差异代谢物最多,达39 种,其物质累积变化主要发生在T2~T3阶段,即由变黄期至定色期的过度阶段,代谢物含量呈现明显上升,随后在T3~T5阶段趋于稳定;class 4和class 7的表达模式类似,分别包含差异代谢物18和17种,这些代谢物随着烘烤时间的推移呈现持续上调趋势,随后在T4~T5阶段略微下调,在T2~T3过度阶段上调最明显;而class 6包含的代谢物位居第四,达8 种,代谢物呈现先上调再下调,随后略微上调的趋势。Class 3 包含的差异代谢物达7 种,这些代谢物从T1至T3阶段呈现明显上调趋势,在T3阶段达到最高,随后下降。此外,class 1、class 3和class 8模式的差异代谢物较少,仅分别为3,5和4。
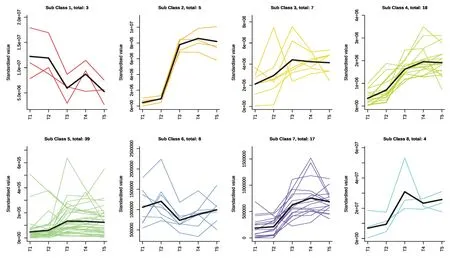
图4 多酚类差异代谢物变化模式聚类分析
2.4 多酚类差异代谢物动态变化
多酚类物质是酶促棕色化反应的主要底物,为探究酶促棕色化反应的代谢中间产物和代谢通路,进一步对鉴定的多酚类差异代谢物进行分析,结果发现,在分析的5 个烘烤节点中,酚酸类有62 种,主要为苯甲酸、苯乙酸、咖啡酸、阿魏酸、香草酸等;黄酮类38 种,主要包括二氢黄酮、黄酮,黄酮醇、查耳酮等物质,图5和图6 分别显示差异倍数|Log2FC|≥2 的31 种酚酸类物质和11种黄酮类物质。由图5可知,随着烘烤时间的增加,这31种酚酸类物质主要以上调为主。其中分支Ⅰ中的8 种代谢物在T1时期含量很低,除了3-甲氧基苯甲酸(3-Methoxybenzoic acid)和4-甲基苯甲酸(4-Methylbenzoic acid)在T3时期明显上调外,其余6 种均在T2时期明显上升,至T3时期继续上调,随后基本保持稳定;分支Ⅱ中的6 种代谢物在T1和T2时期含量相对较低,至T3明显上调,随后基本保持稳定;分支Ⅲ和Ⅳ中的代谢物在T1和T2时期就有相对较高的含量,随着烘烤时间和温度的推移表现上调趋势,在T3阶段之后,含量基本保持稳定,并且分支Ⅳ中的6种代谢物在T3、T4和T5时期含量明显高于其他代谢物。黄酮类代谢物与酚酸类呈现类似的变化趋势(图6),其代谢物含量上调主要发生在T2~T3阶段,大多数代谢物在T3之后基本保持稳定,但是有个别代谢物具有不同的变化趋势,例如 3- 邻乙酰基短叶松素(3-OAcetylpinobanksin)在T5阶段大幅下调,异鼠李素-3-O-葡萄糖苷-7-O-葡萄糖苷(Isorhamnetin-3-O-glucoside-7-O-glucoside)在T2~T4表现连续上调,T4时期达到最高,随后下调。
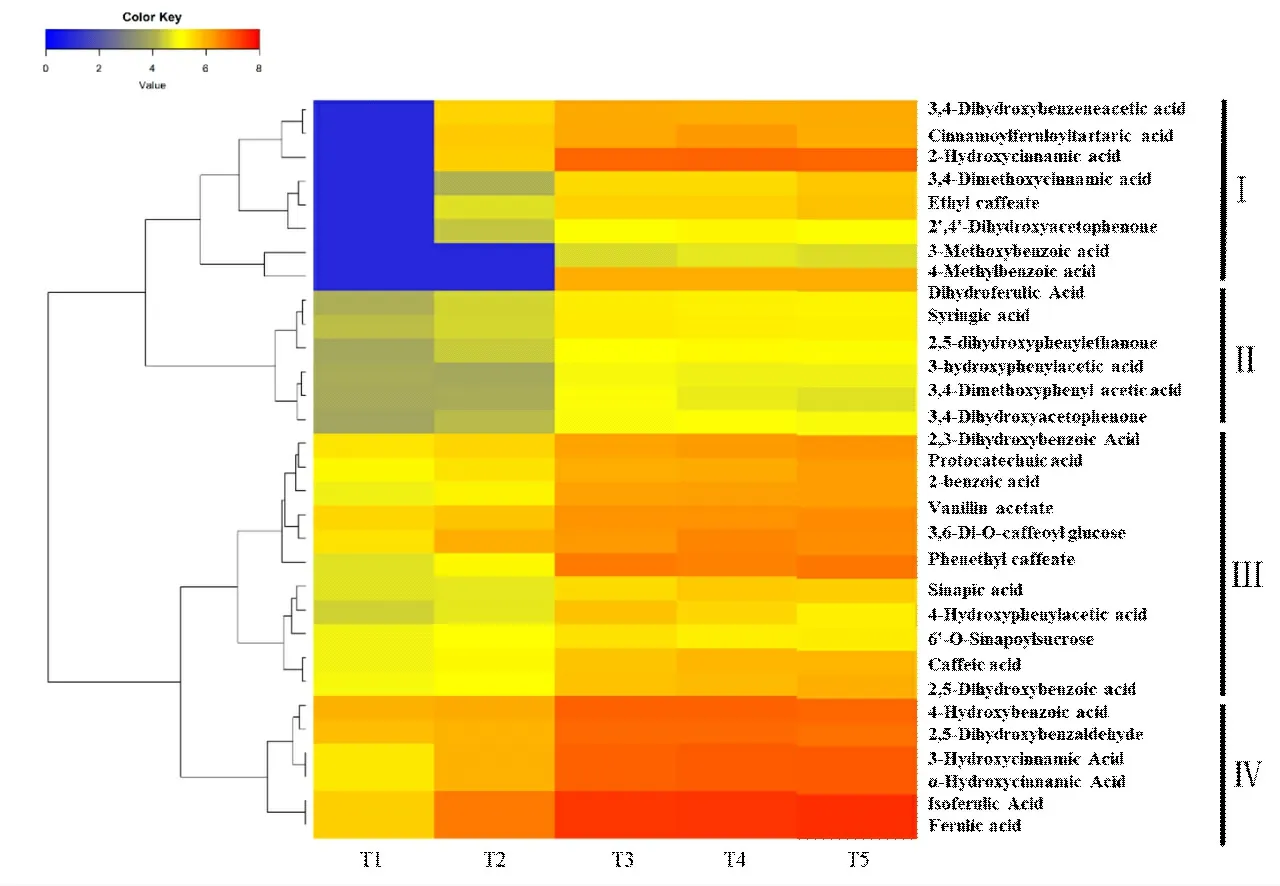
图5 不同烘烤节点烟叶酚酸类差异代谢物丰度变化
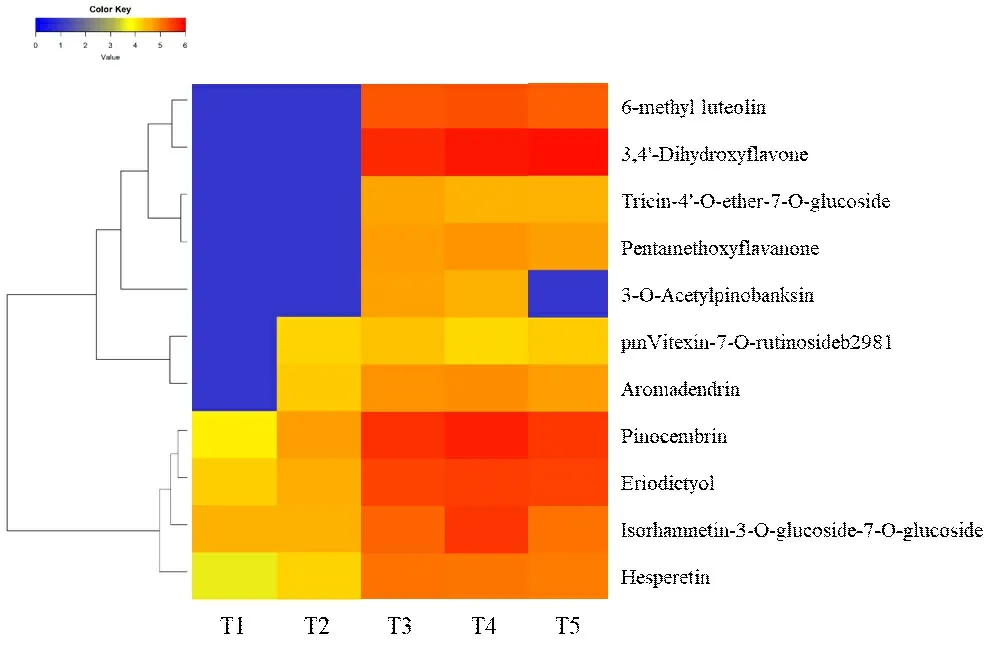
图6 黄酮类差异代谢物丰度变化
2.5 多酚类代谢物代谢通路
多酚类化合物KEGG 代谢通路分析发现,这些代谢物主要富集在4 条代谢通路中,包括酪氨酸代谢(Map 00350),苯丙氨酸代谢(Map 00360)、苯丙烷类生物合成(Map 00940)和类黄酮生物合成(Map 00941)。根据KEGG Pathway 的数据库已知的代谢通路,绘制了CB-1 上部叶烘烤过程中主要差异代谢物富集的生物合成代谢通路(图7),这些代谢通路主要涉及与烟叶风味品质相关的氢醌、绿原酸、山奈酚和芥子酸等物质的生物合成。研究发现,苯丙氨酸代谢通路和酪氨酸代谢途径均可生成2,5-二羟基苯乙酸,转化形成2,5-二羟基苯甲醛、再转化成2,5-二羟基苯甲酸甲酯,最后形成氢醌(图7A)。本研究中,合成氢醌通路上的代谢物均在T2~T3阶段发生了明显的上调。苯丙烷类生物合成通路中(图7B),苯丙氨酸合成咖啡酸,转化形成阿魏酸,在酶的作用下转化为芥子酸,这条通路上的代谢物大多在T1~T3阶段发生了上调。类黄酮生物合成通路中(图7B),差异代谢物变主要集中在三条途径,一是由苯丙氨酸合成的肉桂酰-CoA,进一步转化对香豆酸CoA,经对香豆酰奎尼酸合成绿原酸,或者经对香豆酰莽草、咖啡酰,最后合成绿原酸;另一条通路上,肉桂酰-CoA经柚皮素查尔酮、柚皮素、二氢山奈酚,形成山奈酚;此外,由肉桂酰-CoA,经松属素查尔酮、乔松素、短叶松素,最后形成短叶松素-3-乙酸酯。
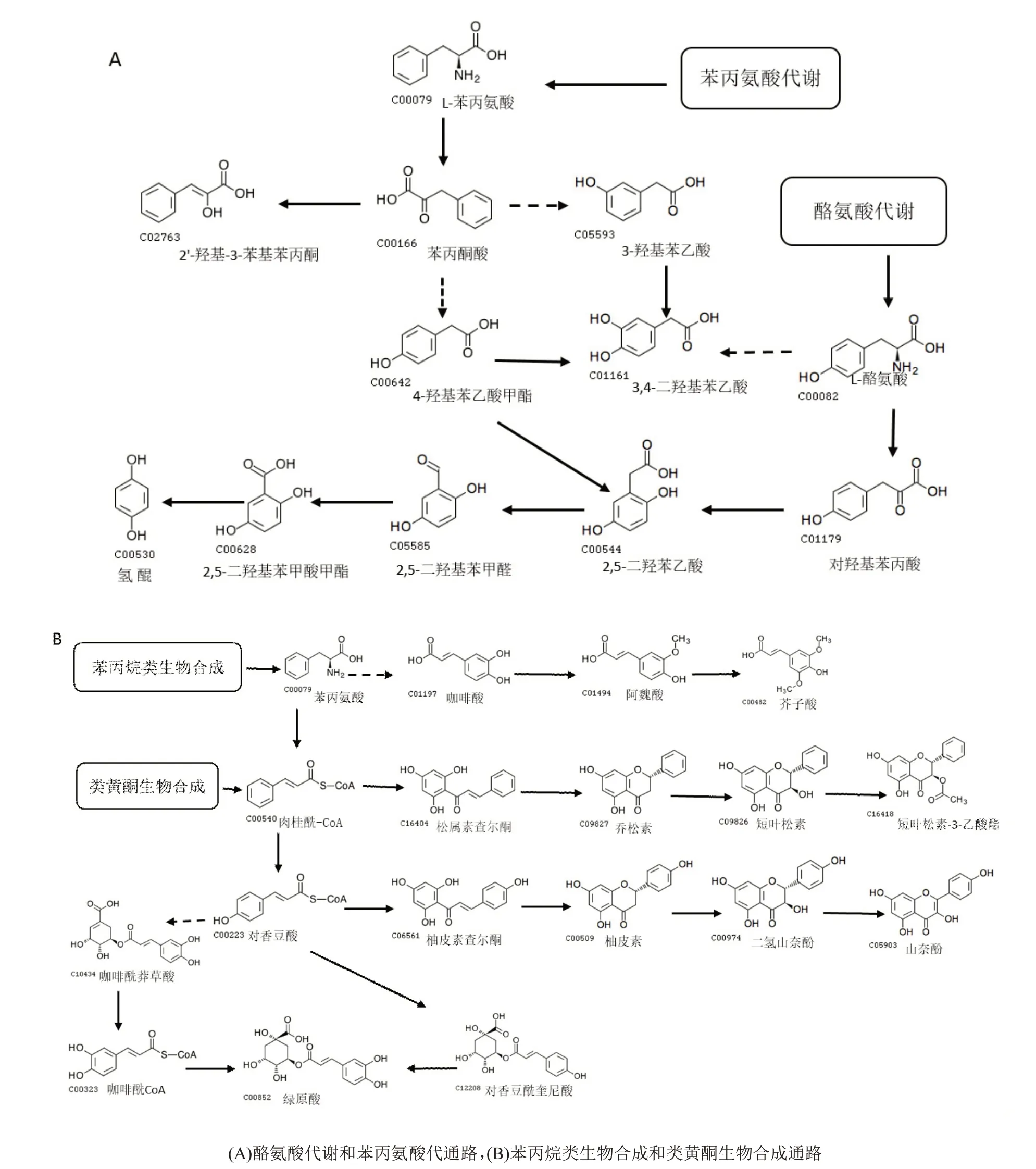
图7 多酚类差异代谢物的代谢网络分析
3 讨论与结论
酚类物质在烟叶调制特性,色泽等级和烟气香味特征形成中都具有重要作用[2]。多酚化合物不仅能够赋予烟叶某些特定的香气,在烟草燃烧时,酚类会产生酸性反应,中和烟草制品中的部分碱性,使烟叶的吃味趋于醇和。烟叶陈化过程中,烟叶中多酚类物质会发生棕色化反应,使叶片颜色明显变深,其吃味也会变得更醇。然而,在烘烤过程中,如果多酚类物质发生过度氧化,就容易产生过度棕色化反应,造成内在化学成分比例失调,从而使烟叶颜色和内在品质变差。一般而言,植物组织的褐变反应包括酚类物质、酶和氧气三大要素[17]。多酚氧化酶(PPO)是棕色化反应中最主要的酶[18],以往的研究发现,PPO 酶在43℃~46℃之间具有较高的活性[2],说明43℃~46℃区间是棕色化反应的敏感期。本研究中,从烘烤过程重要温度节点的代谢组学分析可以看出,大部分酚类物质含量在43℃之后趋于稳定(图5,图6),表明大部分酚类物质没有被氧化成醌,该烘烤工艺下不会发生过度棕色化反应。其原因可能是定色期后,烟叶水分含量较低,PPO酶缺乏适宜的反应场所。前人研究表明,烘烤阶段的变黄末期至定色初期是酶促棕色化反应敏感期,但是当含水量小于原来含水量的50%,酶促棕色化反应就较难发生[19-22]。由此可见,在定色之前,可通过降低烟叶细胞的含水量抑制PPO 活性,调控棕色化反应的速度,防止烟叶过度褐变。
烟草中的酚类物质种类众多,其中绿原酸和芸香苷是烟草中最主要的酚类物质[23]。本研究的差异代谢物富集在酪氨酸代谢,苯丙氨酸代谢、苯丙烷类生物合成和类黄酮生物合成等4 条通路中,这些通路均与绿原酸和芸香苷的合成相关。通常情况下,多酚的物质代谢起始于苯丙氨酸和酪氨酸等芳香氨基酸,在苯丙氨酸解氨酶(PAL)的催化下形成对香豆素,随后在一系列相关酶的作用下反应形成苯丙烷类和黄酮类物质。绿原酸的生物合成属于苯丙烷类代谢通路的分支,苯丙烷类代谢通路和类黄酮生物合成通路中均可形成绿原酸关键物质肉桂酰CoA(图7B),由此再合成对香豆酸-CoA,然后经对香豆酰奎尼酸合成绿原酸,或者经对香豆酰莽草、咖啡酰,最后合成绿原酸;同时,对香豆酸-CoA也可以在CHS酶的作用下合成黄酮类物质,最终合成芸香苷[24]。本研究中,从变黄期至定色期阶段,形成绿原酸通路上的代谢物大多发生了上调,然而绿原酸的含量并未发生明显变化,其原因可能是部分绿原酸发生了转化。此外,苯丙氨酸代谢通路和酪氨酸代谢通路中,氢醌形成途径上的代谢物大多出现了上调(图7A)。氢醌是抗氧化剂,但是其本身具有光敏性,当处于变黄期至定色期的烟叶从烤房中取出时,烟叶表面颜色容易变成深褐色,其原因可能是由于氢醌见光氧化变黑。值得注意的是,类黄酮代谢分支上,山奈酚和芥子酸合成通路上的部分代谢物在烘烤过程中发生了明显上调,这些物质的转化对烟叶色泽、香气和风味的形成具有重要作用。在香梨绿原酸合成途径的研究中,利用不同发育阶段的转录组和代谢组联合分析,成功鉴定了4 个香梨绿原酸发育的关键基因[25]。基于本研究鉴定的烟叶色泽、香气和风味等关键物质的合成通路,利用代谢组学和转录组学关联分析挖掘通路中的关键基因,可为将来的品种改良提供基因资源。
综上,本研究基于靶向代谢组学解析了烘烤过程中不同温度节点的烟叶代谢组差异,共鉴定出1069种代谢物,其中多酚类物质为374 个,达总数的34.99%。烘烤过程中变黄期至定色初期是多酚类差异代谢物含量变化的主要时期,差异代谢物富集在氢醌、绿原酸、山奈酚和芥子酸等与烟叶风味和品质相关物质的生物合成途径中。本研究结果为烟草品质形成的代谢物质基础剖析提供了信息,为烟叶棕色化反应调控奠定了基础。
