不同基质对红海榄幼苗生长生理特性的影响
2024-04-24贾湘璐欧阳子龙刘佳哲樊东函滕维超石景忠刘秀
贾湘璐,欧阳子龙,刘佳哲,樊东函,滕维超,石景忠,刘秀
(1广西壮族自治区林业科学研究院,南宁 530000;2广西大学林学院,南宁 530004;3钦州市林业科学研究所,广西钦州 535000;4南宁植物园,南宁 530004;5南宁青秀山风景名胜旅游开发有限责任公司,南宁 530004)
0 引言
红海榄(Rhizophorastylosa)是一种重要的红树林植物,具有较高的生态价值和环境效益,可作为海岸带防护林[1-2]。基质选择是植物幼苗培育中的关键步骤,适宜的基质有利于植物更好的生长发育和对环境的适应[3-5]。植物通过根系从基质中获取水分和养分,为枝条、花和果实的生长发育提供所需物质[6]。组分和配比也决定了基质的理化性质[7]。透气透水性、养分有效性和保温性是影响植物根系发育的重要因素。有研究表明,植物根系越发达,农艺性状指标最高[8]。代丽丽等[9]研究也表明,根系指标较高的黄栌更为健康,生长情况更好。
大量研究表明,基质能明显影响植物抗氧化酶活性,刘航江[10]发现不同基质中葎草扦插苗SOD(超氧化物歧化酶)、POD(过氧化物酶)和CAT(过氧化氢酶)活性差异显著,且影响顺序为POD>CAT>SOD。朱上游等[11]研究表明,水稻土:珍珠岩=4:1的基质配比中伴矿景天具有较高的株高、根系形态指标、色素含量及抗氧化酶活性,有利于生长。郭红等[12]进一步研究表明,不同基质对提高碧玉兰响应盐胁迫能力的效果不同,适宜的基质选择将促进产业的发展。可溶性糖和可溶性蛋白作为营养物质和渗透调节物质,在植物生长过程中起着重要的促进作用。张瑞博等[13]、宫琳琳等[14]、Yu等[15]和胡小京等[16]研究均表明,合适的基质能提高植物可溶性糖和可溶性蛋白的含量。本研究通过对不同配比基质处理下红海榄幼苗生长指标、生物量、根系形态指标及生理指标进行测定,以期筛选出较佳的育苗基质模式,进一步促进红海榄的培育。
1 材料与方法
1.1 试验地概况
试验地位于钦州市林业科学研究所苗圃(108°62′E,21°96′N),年均气温22℃,属南亚热带季风气候。年均降水量1600 mm,雨量充沛。
1.2 试验材料
试验材料为长势一致、生长状况良好且已长出一对真叶的红海榄幼苗,每15 株幼苗种植于40 cm×50 cm 的塑料框内,为1 个重复,每处理设置3 个重复。正常浇水管理,保持框内水分充足。选择钦州市滩潮淤泥和沙子作为基质材料。经测定,原始泥中氮含量为0.51±0.02 mg/kg,磷含量为0.3±0.02 mg/kg,钾含量为7.64±0.02mg/kg;沙中氮含量为0.11±0.001mg/kg,磷含量为0.11±0.003 mg/kg,钾含量为1.7±0.06 mg/kg。
1.3 试验方法
设置4种不同基质处理即:沙、沙:泥(7:3)、沙:泥(3:7)、泥的基质配比模式。2021 年11 月开始,每月进行一次数据测定,连续测定3个月,于2022年1月结束。
1.4 指标测定
株高和茎粗分别使用皮尺和游标卡尺测定,叶长、叶宽和叶面积使用叶面积仪测定。使用WinRHIZORA根系扫描仪测定各项根系形态指标。测定生物量时先将样品置入115℃的烘箱内杀青半小时,再以80℃~85℃烘至重量不变。各项色素和生理指标的测定均参考《植物生理学实验指导》[17]。
1.5 数据分析
采用DPS 7.05 软件进行方差分析和多重比较分析,SPSS 25.0 软件进行主成分分析和综合得分计算,采用Origin 2019软件作图。
2 结果与分析
2.1 不同基质处理对红海榄幼苗生长的影响
如图1所示,4种基质处理红海榄幼苗叶长、叶宽、叶面积、株高、茎粗均随时间而增加,在2022年1月最长。沙:泥(7:3)处理在3 个时期(2021 年11、12 月和2022年1月)叶长、叶宽、叶面积和株高均最大。
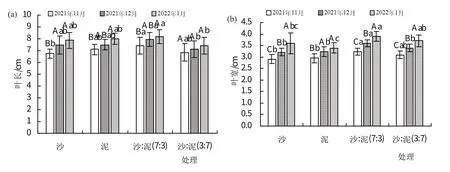
图1 不同基质对红海榄幼苗生长的影响
泥处理在3个时期茎粗均最大,分别为4.44、4.64、4.79 mm。
2.2 不同基质处理对红海榄幼苗根系的影响
由表1可知,各处理间红海榄幼苗总根长、总根表面积、总根体积、总根尖数、分枝数均差异显著,根平均直径差异不显著。总根长在沙:泥(7:3)基质处理中最长,为514.67 cm;在泥处理中最短,为243.7 cm。总根表面积在沙:泥(7:3)基质处理中最大,为32.42 cm2;在泥处理中最小,为10.41 cm2。总根体积在沙:泥(7:3)基质处理中最大,为0.32 cm3;在泥处理中最小,为0.12 cm3。总根尖数在沙处理中最多,为7234个;在泥处理中最少,为2826个。分枝数在沙:泥(7:3)基质处理中最多,为2280.5个;在泥处理中最少,为1097.75个。
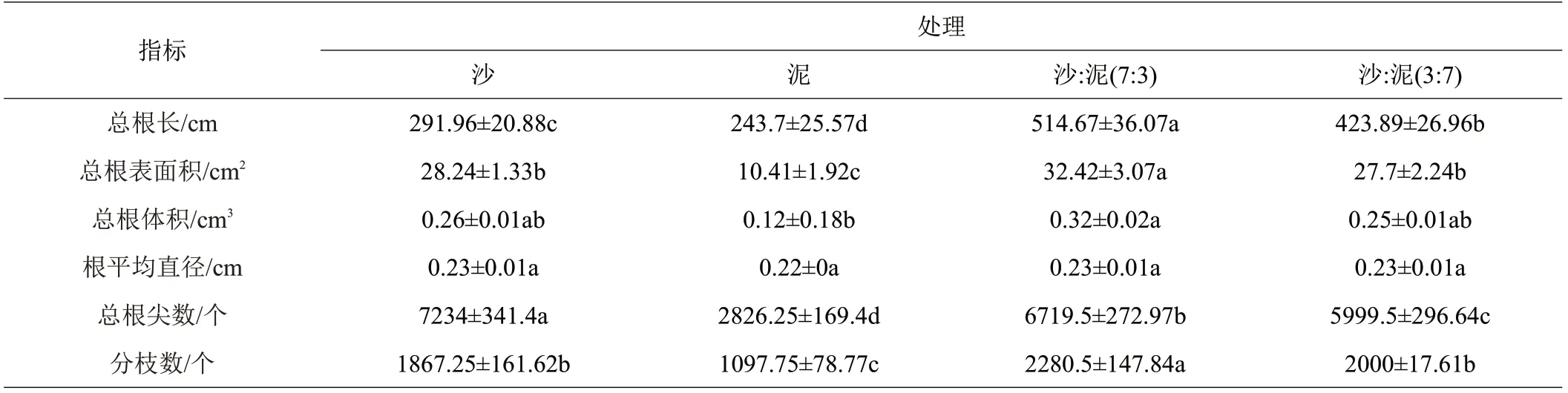
表1 不同基质对红海榄幼苗根系的影响
2.3 不同基质处理对红海榄幼苗生物量的影响
由表2 可知,各处理间红海榄幼苗根、胚轴、茎、叶、总生物量均差异显著。根生物量在沙处理中最大,为1.12 g;在泥处理中最小,为0.51 g。胚轴生物量沙处理中最大,为11.16 g;在泥处理中最小,为7.65 g。茎生物量在沙:泥(3:7)基质处理中最大,为0.76 g;在沙处理中最小,为0.43 g。叶生物量在沙:泥(3:7)基质处理中最大,为1.72 g;在泥处理中最小,为1.03 g。总生物量在沙:泥(7:3)基质处理中最大,为13.81 g;在泥处理中最小,为9.64 g。
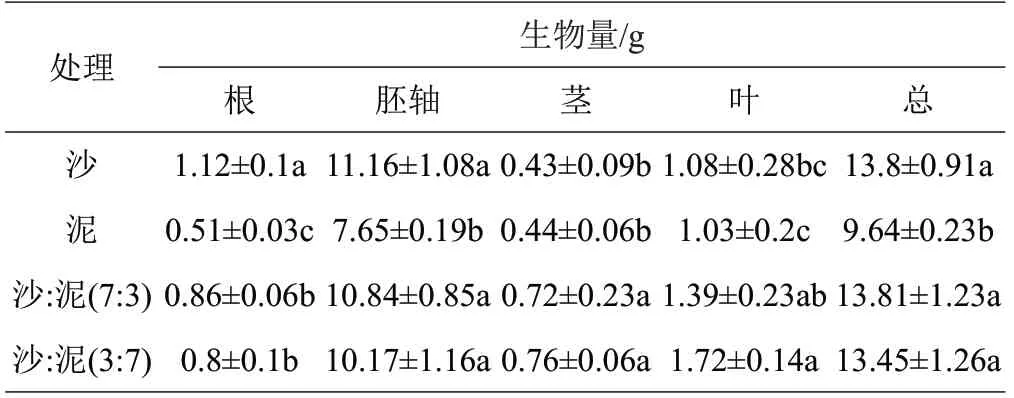
表2 不同基质对红海榄幼苗生物量的影响
2.4 不同基质处理对红海榄幼苗色素含量的影响
如图2(a)所示,随时间增加,红海榄幼苗叶绿素a(Chla)含量在沙处理和沙:泥(7:3)处理中均逐渐降低,在2022 年1 月最低,分别为0.165、0.172 mg/g;在泥处理和沙:泥(3:7)处理中均先升高再降低,在2021 年12月最高,分别为0.286、0.294 mg/g。
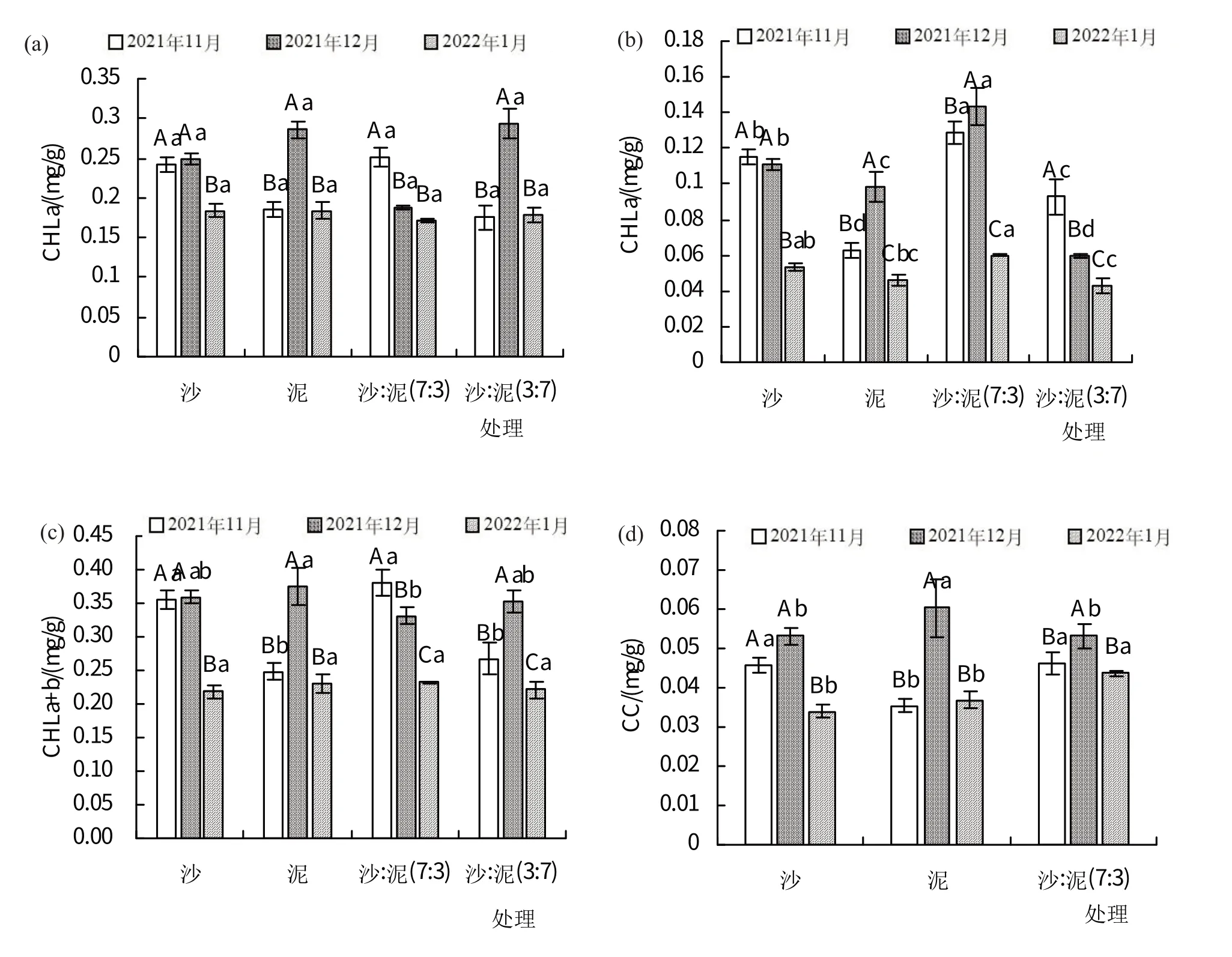
图2 不同基质对红海榄幼苗色素含量的影响
如图2(b)所示,随时间增加,红海榄幼苗叶绿素b(Chlb)含量在沙处理和沙:泥(3:7)处理中均逐渐降低,在2022 年1 月最低,分别为0.054、0.043 mg/g;在泥处理和沙:泥(7:3)处理中均先升高再降低,在2021 年12月最高,分别为0.098、0.144 mg/g。
如图2(c)所示,随时间增加,红海榄幼苗叶绿素a+b(Chla+b)含量在沙处理和沙:泥(7:3)处理中均逐渐降低,在2022年1月最低,分别为0.219、0.232 mg/g;在泥处理和沙:泥(3:7)处理中均先升高再降低,在2021 年12月最高,分别为0.375、0.353 mg/g。
如图2(d)所示,随时间增加,红海榄幼苗类胡萝卜素(CC)含量在沙处理和处理中均逐渐降低,在2022年1 月最低,分别为0.034 mg/g;在泥处理、沙:泥(3:7)处理和沙:泥(7:3)中均先升高再降低,在2021 年12 月最高,分别为0.053、0.06、0.053 mg/g。
2.5 不同基质处理对红海榄幼苗抗氧化酶和渗透调节物质的影响
如图3(a)所示,2021年12月红海榄幼苗叶片SOD活性在沙处理中最高,为807.98 U·g/min FW,在泥处理中最低,为760.57 U·g/min FW;其余两个采样时期处理间差异不显著。除泥处理外,其余3 个处理SOD酶活性均在2022 年1 月最高,分别为826.75、816.3、817.6 U·g/min FW。
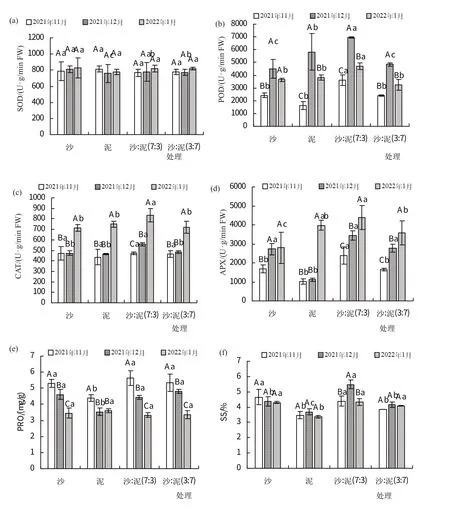
图3 不同基质对红海榄幼苗抗氧化酶和渗透调节物质的影响
如图3(b)所示,各处理中红海榄幼苗叶片POD活性随时间均呈现出先升高再降低的趋势,2021年12月最高,分别为4503.33、5814、6963.33、4848.667 U·g/min FW;2021 年11 月最低,分别为2428、1602.67、3611.33、2404 U·g/min FW。
如图3(c)所示,各处理中红海榄幼苗叶片CAT 活性随时间均呈现出逐渐升高的趋势,2022年1月最高,分别为716.33、748.67、836.67、719.33 U·g/min FW。
如图3(d)所示,各处理中红海榄幼苗叶片APX 活性随时间均呈现出逐渐升高的趋势,2022年1月最高,分别为2793.37、3980.01、4379.99、3573.33 U·g/min FW;2021 年11 月最低,分别为1686.67、1030、2380、1653.33 U·g/min FW。
如图3(e)所示,各处理中红海榄幼苗叶片PRO 含量随时间均呈现出逐渐降低的趋势,2021 年11 月最高,分别为5.3、4.39、5.64、5.36 mg/g。如图4(f)所示,沙处理中红海榄幼苗叶片SS 含量随时间呈现出逐渐降低的趋势,2021年11月最高,为4.66%,2022年1月最低,为4.3%;其余3个处理均先升高再降低,2021年12月最高,分别为3.69%、5.49%、4.16%。
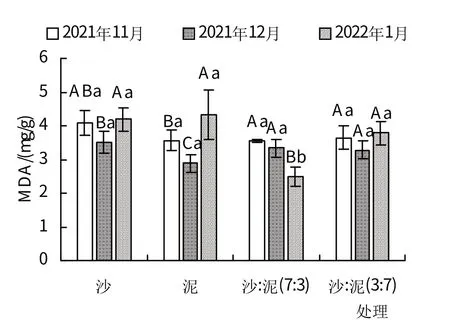
图4 不同基质对红海榄幼苗MDA含量的影响
2.6 不同基质对红海榄幼苗MDA(丙二醛)含量的影响
如图4 所示,沙:泥(7:3)处理中红海榄幼苗叶片MDA 含量随时间呈现出逐渐降低的趋势,2021 年11月最高,为3.55 mg/g,2022 年1 月最低,为2.49 mg/g;其余3 个处理均先降低再升高,2021 年12 月最低,分别为3.51、2.88、3.28 mg/g;2022 年1 月最高,分别为4.19、4.32、3.78 mg/g。
2.7 不同基质对红海榄幼苗生长生理指标的主成分分析
为探究不同基质对红海榄幼苗的促进作用,选择第3 个采样时期(2022 年1 月)各项生长生理指标(除MDA 外)进行主成分分析。如表3 所示,共有3 个主轴,LW、总根表面积、根平均直径、总根尖数、分枝数、胚轴生物量、总生物量、PRO、SS是第一轴的主要影响因子;CAT、APX、CHLa+b 是第二轴的主要影响因子;叶生物量、茎生物量是第三轴的主要影响因子。
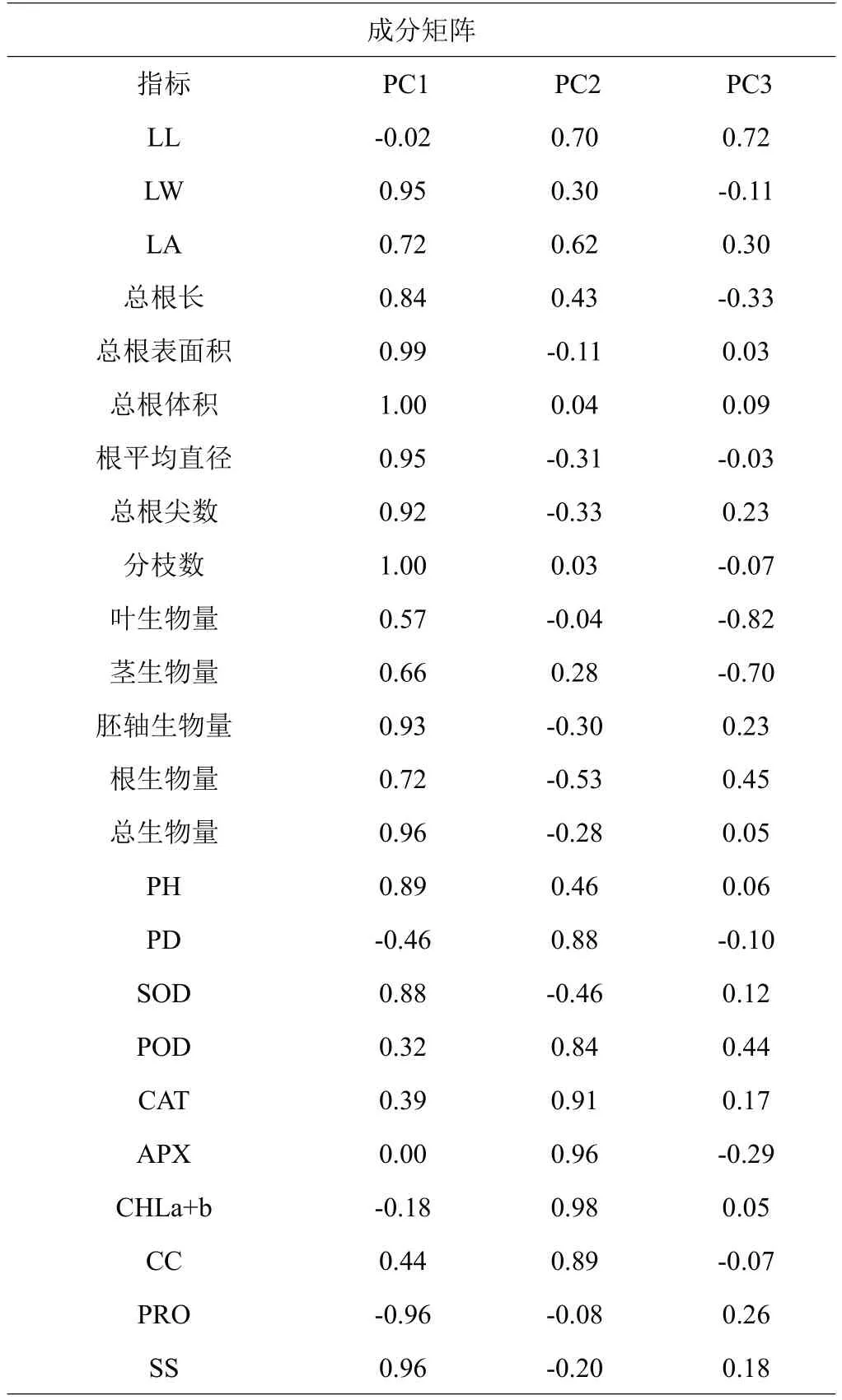
表3 红海榄幼苗生长生理指标主成分矩阵
从表4 可知,第一、二、三轴分别占58.23%、30.68%、11.09%的贡献率,累积贡献率为100%,能完好反映各生理指标的相对重要性和相互关系。由综合得分因子可知,沙、泥、沙:泥(7:3)、沙:泥(3:7)4 种处理模式综合得分分别为-0.23、-2.02、2.72、-0.47,由大到小排序为沙:泥(7:3)>沙>沙:泥(3:7)>泥,以上说明沙:泥(7:3)处理对提高红海榄幼苗生理活性、促进生长效果最佳,而泥处理模式效果不佳。
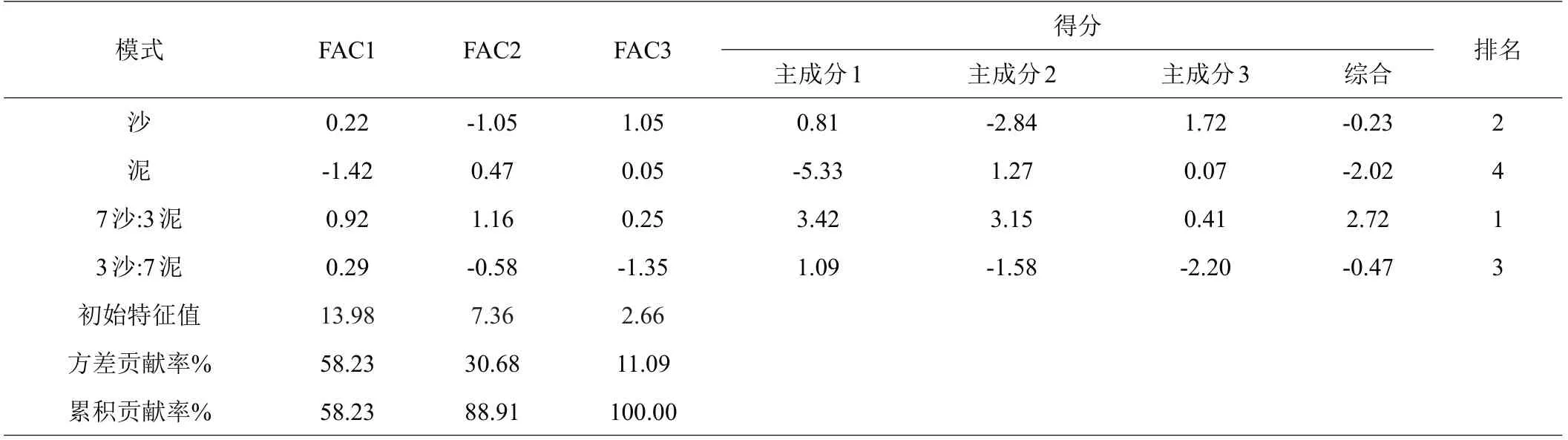
表4 红海榄幼苗生长生理指标主成分综合得分
3 讨论与结论
3.1 不同基质对红海榄幼苗生长特性的影响
基质种类和配比决定了土壤物理性质和养分特性,对于植物生长起着重要的决定作用[18]。在植物的生长过程中,基质能通过影响根系伸长来为植物提供更多的水分和养分,进而决定植物的生长状态和存活率,所以选择合适的基质对植物的生长繁殖尤为重要[19-20]。卢万义[21]对不同基质配比处理沃柑的生长指标进行测定发现,株高、地径、根系指标和生物量在不同处理间均存在显著差异,表明不同基质对沃柑生长起到了明显影响,与本研究的结果一致。陈赣等[22]研究发现不同配比的基质对西红柿生长的影响差异较大,以草炭:珍珠岩:蛭石=1:1:1 处理下生长最好、产量最高。叶长、叶宽和叶面积作为叶片功能性状指标,反映叶片形状和对光资源的利用程度,一般叶面积越大,对光的捕获能力越强,越有利于固定生长所需的碳[23-24]。本研究表明,沙:泥(7:3)基质处理的红海榄幼苗叶片功能性状指标在3 个时期均最大,而该处理的总生物量也最大,说明叶片面积增大可能提高了光合作用效率进而促进总生物量增加。陈云峰等[25]研究表明,黄心土:椰糠=1:1 基质处理下红椿生长效果较好,株高可达47.01 cm,地径达6.18 mm。本研究发现沙:泥(7:3)基质处理的红海榄幼苗株高在3 个时期均最大,而泥处理的茎粗在3个时期均最大,说明在红海榄幼苗的生长发育进程中可能存在株高和地径的平衡,沙:泥(7:3)基质处理能有效促进茎的生长使植株升高,而泥处理能使茎变粗。
根系是植物从基质环境中获取营养物质最直接的器官,决定了植物生长状态和水平[26]。及利等[27]通过不同配比基质试验发现,泥炭土:珍珠岩:木耳菌棒=3:3:4处理下核桃楸苗根系指标明显提升,根系生物量和根平均直径分别增加了6.22 g/株和76.54%。本研究中红海榄幼苗总根长、总根表面积、总根体积、根平均直径和分枝数均为沙:泥(7:3)基质处理最好,说明该处理对促进根系生长的效果较好,更有利于养分的吸收。涂远国等[28]、夏康生等[29]和Li 等[30]也得到类似的研究结果,各项根系指标的值均在同一处理中较高,说明根系发育在多个方面具有同步作用。本研究还注意到总根尖数在沙处理中最高,且在4 种基质处理中随着沙的比例降低总根尖数也下降,可能的原因是沙孔隙度较高,透水透气性较强,有利于根尖分裂。本研究中泥处理总生物量最低,可能的原因一方面是泥结构较为紧密,限制了生长初期根系的伸长,根系发育较差导致养分吸收效率不高,整体生长情况较弱;另一方面是和叶面积和叶生物量最低,光合作用能力较差。沙、沙:泥(7:3)基质和沙:泥(3:7)基质处理总生物量均较高,有利于生长发育和干物质的积累。
3.2 不同基质处理对红海榄幼苗生理特性的影响
色素是维持植物叶片生理活性的重要物质,叶绿素能促进光合作用的进行[31],提高植物对碳的积累;类胡萝卜素能维持叶绿素含量的稳定,辅助光的捕获并提升植物的光保护能力[32]。本研究中叶绿素和类胡萝卜素含量均在2021年12月最高,说明该时期下红海榄幼苗具有较强的光合作用能力,生长效率处于较高的水平;而2022年1月各色素含量降低,可能是因为该时期气温进一步降低,导致植物处于低温胁迫,影响了正常的生理过程。Odhiambo 等[33]通过对辣椒的研究也发现低温下叶片叶绿素含量出现下降的趋势,Kato等[34]则发现低温胁迫明显降低了优雅草类胡萝卜素含量。
抗氧化酶SOD、POD、CAT、APX 可以协同清除植物体内的活性氧(ROS),减少ROS 对细胞产生的伤害[35-36]。抗氧化酶活性决定了植物生理状况和对逆境环境的适应程度,是一种重要的植物逆境保护指标。本研究中沙:泥(7:3)基质处理的POD、CAT、APX 活性在各采样时期均为最高,说明该处理提高红海榄适应冬季自然低温胁迫能力的效果较好,有利于维持正常生长发育。CAT 和APX 活性均随时间逐渐升高,在2022 年1 月最高,表明红海榄幼苗能通过不断提高这两种酶的活性来响应逐渐低温。POD 活性在2021 年12 月最高,随后出现下降,可能的原因一方面在于2022 年1 月红海榄幼苗降低后的POD 活性足以抵御自然低温导致的ROS 积累,无需提高活性;另一方面的原因在于不同抗氧化酶之间存在互补效应,当某种酶活性较低时,植物能通过提高另外的酶活性维持整体抗氧化系统的稳定性。值得注意的是,2021年11月泥处理中的POD、CAT、APX活性均最低,这可能是由于生长初期泥不利于根系发育,从而降低了整体生理活性水平。本研究中SOD活性不随时间而发生变化,较为稳定,这可能是由红海榄幼苗生长过程中的生理特性决定的。
可溶性蛋白和可溶性糖为植物生长发育、生理活动提供物质和能量来源,同时可作为渗透调节物质参与对ROS 的响应当中,维持植物生理稳定性[37-38]。各处理可溶性蛋白含量均随时间而降低,说明红海榄幼苗在生长和对抗自然低温过程中消耗了可溶性蛋白。可溶性糖含量表现不一,在沙处理中随时间逐渐降低,而在其余3 种处理中先升高再降低,可能的原因是其余3 种处理短暂地促进了可溶性糖的积累,但随温度降低和植物生长需要而降低。
沙:泥(7:3)基质处理中红海榄幼苗叶片MDA含量随时间而降低,说明该处理下植株整体生理状况较好,足以应对自然低温。而其余3个处理均随时间先降再升,说明2022年1月低温造成了MDA的积累,不利于植物生理过程的稳定。
3.3 不同基质处理对红海榄幼苗的综合影响
根据主成分综合得分排序可知,沙处理得分最高,泥处理得分最低,且随沙比例降低,总的综合得分逐渐降低,说明沙可以提高红海榄幼苗整体生长状况。泥中富含有机质和营养元素,而沙的养分含量较泥低,沙处理幼苗生长初期虽效果较好,但随着生长发育的进行,基质中养分大量消耗,沙可能无法提供生长后期所需养分。钦州海滩分布有大面积的海泥,是种植红树林植物天然的基质。红树林既对海岸线生态多样性的稳定具有重大意义,又能防风固土,降低水土流失[39-40]。在钦州海岸区域推广种植红树林植物有利于整个生态系统功能的构成和完善,也有助于推动当地红树林产业的发展[41]。因此,对于红海榄幼苗培育的研究是推广的一个关键步骤,由初始阶段的沙基质转为后期富含有机质的泥基质可能更好的促进整体生长效果,缩短生长周期。本研究不足之处在于仅对红海榄幼苗生长初期指标进行测定,后期应需继续跟踪植株的生长效果以及时转变栽培措施及养分添加,以期更加完整地探索红海榄幼苗整个生长发育周期的特性。
3.4 结论
沙:泥(7:3)基质处理中红海榄幼苗生长指标、根系指标、生物量、光合色素含量和活性氧清除能力均较佳,是最佳的基质配比模式。滩潮淤泥在钦州普遍使用,但泥处理红海榄幼苗生长初期各项指标较差,未来需补充一定程度的沙以改善基质,提高红海榄幼苗的生长生理水平。
