邻苯二甲酸酯降解功能内生菌群的筛选及定殖效能
2024-03-28马俊超高彦征左翔之凌婉婷南京农业大学土壤有机污染控制与修复研究所江苏南京210095
张 帅,王 建,马俊超,高彦征,左翔之,凌婉婷(南京农业大学土壤有机污染控制与修复研究所,江苏 南京 210095)
邻苯二甲酸酯(PAEs)被广泛应用于塑料、化妆品、油漆等行业[1-3],全球年产量高达3 亿t[4].作为增塑剂,PAEs 与塑料产品之间以氢键或范德华力连接,随着时间的推移,很容易通过各种途径向环境中释放.同时作为一种持久性和疏水性有机污染物,PAEs极易在土壤中积累[5],并通过各种途径暴露于人体从而引发癌症、内分泌干扰性、生殖和神经毒性等危害,严重威胁人体健康[6].当前,美国和欧盟已将邻苯二甲酸二甲酯(DMP)、邻苯二甲酸二乙酯(DEP)、邻苯二甲酸二丁酯(DBP)、邻苯二甲酸丁基苄基酯(BBP)、邻苯二甲酸二(2-乙基)己酯(DEHP)和邻苯二甲酸二辛酯(DnOP)6 种PAEs 列为优先控制污染物[7],我国也将DMP、DBP、DnOP 定为环境优控污染物[8].
土壤中的PAEs 可被作物根系吸收并向地上部转移,造成农作物可食部位PAEs污染.长江三角洲和珠江三角洲地区的调查结果显示,我国蔬菜等农作物中普遍检出 PAEs,平均含量分别为 0.536,2.84mg/kg,主要以DBP、DEHP 和DnOP 为主,其中,珠三角地区叶菜类蔬菜中DEHP 的含量远超美国和欧盟建议的标准,严重威胁了人体健康[9-10].
植物内生细菌是栖息在植物组织内部,能够促进植物生长,提高植物抗病和抗逆能力并与其互利共生的一类细菌.许多内生细菌具备有效降解有机污染物的能力[11-12].近年来,一些PAEs 降解的内生细菌如芽孢杆菌属(Bacillus)、戈登式菌属(Gordonia)、假单胞菌属(Pseudomonas)等已经被分离出来[13-14].将分离获得的功能菌株重新定殖回作物体内,可有效减低作物体内有机污染物的积累[15].然而,实际污染区往往是多种PAEs共存,单一菌株降解PAEs 的能力有限,而不同来源的PAEs 降解菌株间的复配往往会存在拮抗作用,最终影响功能内生细菌的原位定殖效能.实际污染区作物体内存在经自然驯化且功能稳定的菌群,能否将其富集驯化并通过定殖的方式实现作物体内PAEs 的直接消减,对此,国内外相关研究较少.
本研究从PAEs 污染区生长的蔬菜中富集驯化具有PAEs 降解能力的功能内生菌群,解析菌群群落组成;验证菌群降解效能;优化菌群降解PAEs的条件;借助水培体系验证菌群对作物体内PAEs的消减效能及促生效应,旨在为消减作物体内PAEs 污染、保障污染区农业生产安全和人群健康提供参考.
1 材料与方法
1.1 试验材料
1.1.1 供试植株 空心菜(Ipomoea aquatica Forsk.)采自被PAEs 污染的土壤中.
1.1.2 主要试剂 DMP、DEP、DBP 和BBP 购自上海阿拉丁生化科技股份有限公司,纯度均为99.0%.霍格兰营养液(HNS).丙酮、正己烷、甲醇、乙腈均为色谱级.
培养基配制:LB 液体培养基(1.0L):10.0g 进口蛋白胨、5.0g 进口酵母提取物、10.0g NaCl. 无机盐液体培养基(1.0L):1.50g(NH4)2SO4、0.50g KH2PO4、0.50g NaCl 、 1.91g K2HPO4·3H2O 、 0.20g MgSO4·7H2O.涉及到的所有培养基均在高压灭菌锅中121℃条件下灭菌20min.
30%甘油:超纯水与丙三醇按7:3的体积比混合,灭菌后置于4℃冰箱保存.
1.2 内生菌群的筛选
空心菜经过表面消毒后,将其移入无菌研钵,剪碎,加入无菌水充分研磨.静置5min 后,移取适量上清液至添加PAEs(5mg/L)的无机盐液体培养基中,30℃ ,150r/min 避光振荡培养.5d后,转接上层培养液至新配制的添加PAEs(10mg/L)的无机盐液体培养基中,富集驯化培养5d,之后梯度增加PAEs 的浓度并重复以上步骤.如此共富集驯化培养20d,以获得稳定的具有高效降解PAEs 能力的功能内生菌群.最后,将获得的菌液与30%甘油按照体积比1:1 的比例均匀混合,于-80℃冰箱保存备用.
1.3 菌群生长曲线测定
于超净台内挑取-80℃条件下保存的菌种于LB液体培养基中,30℃、150r/min 避光振荡培养,定时取样测定菌液的OD600值.
1.4 菌群的降解效能测定
1.4.1 功能内生菌群菌悬液制备 于超净台内挑取-80℃条件下保存的菌种至5% LB 液体培养基中,30℃、150r/min 避光振荡培养8h,离心收集菌体,用无机盐培养液洗涤菌体2 次,再次离心后弃去上清液,收集菌体,用无机盐培养液调整菌悬液OD600为1.0,于4℃冰箱中保存备用.
1.4.2 功能内生菌群降解效能测定 将制备好的菌悬液按照5%的比例接种到添加PAEs(5mg/L)的无机盐液体培养基中,30℃,150r/min 避光振荡培养,定时取样测定剩余PAEs 含量.
1.5 菌群群落结构分析
1.5.1 样品收集 取一定量经LB 液体培养基活化后的菌群,离心,弃去上清液,收集菌体后交由美吉生物公司进行检测.
1.5.2 DNA 抽提和PCR 扩增 采用E.Z.N.A.®soil DNA kit(Omega Bio-tek,Norcross,GA,U.S.)试剂盒进行微生物群落总 DNA 抽提.使用通用引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’) 以及806R(5’-GGACTACHVGGGTWTCTAAT-3’)对16SrRNA 基因V3-V4 可变区进行PCR 扩增.扩增程序如下:95℃预变3min,27个循环(95℃变性30s,55℃退火30s,72℃延伸30s)后72℃稳定延伸10min.
1.5.3 Illumina Miseq 测序 将同一样本的PCR 产物混合后使用2%琼脂糖凝胶回收PCR 产物,利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City, CA, USA)进行回收产物纯化,2%琼脂糖凝胶电泳检测,并用Quantus™ Fluorometer(Promega,USA)对回收产物进行检测定量.
1.6 环境因子对降解的影响
取配制好的无机盐培养基,分别设置不同pH值、盐度、底物浓度的试验组,按照5%的接菌量接种菌液,30℃,150r/min 避光振荡培养,同时设置不同温度的试验组,7d 后取样测定培养液中PAEs 含量.
1.7 菌群定殖及水培试验
选取水稻为接种对象,本研究设置浸根定殖(ZR)和接种灭活菌群对照组(ZK),各接种处理的水稻暴露于含10mg/L PAEs 的Hoagland 培养液中.
接种方法如下:水稻种子经表面消毒,催苗育种7d后,选取10株长势一致的植株,菌悬液浸泡水稻根部8h,无菌水冲洗后,将植株移入添加PAEs(10mg/L)的Hoagland 培养液中,置于昼夜温度25℃ /20 ℃的光照培养箱中培养,15d 后采集植物样品进行PAEs 含量和生物量等指标的测定.
1.8 植物PAEs 含量分析
称取0.2g 制备好的植物样品于25mL 玻璃离心管中,添加15mL 丙酮-正己烷溶液(V/V, 1:1),涡旋混匀0.5~1min,水浴超声45min.随后以2000r/min 离心10min.取上清液倒入活化后(10mL 正己烷分两次淋洗)的层析柱中(层析柱上层为2g 无水硫酸钠,下层为2g 硅胶),再次添加丙酮-正己烷溶液(V/V, 1:1),同上,重复2 次.洗脱之后分3 次添加15mL 丙酮-正己烷溶液(V/V,1:9).收集混合液并借助旋转蒸发仪(40℃、100r/min 条件下)浓缩至干,移取2mL 甲醇润洗定容,随即倒入带有0.22μm 有机相滤头的玻璃注射器,过滤进2mL 棕色液相小瓶,使用HPLC 分析.
HPLC 条件:色谱柱为Ф4.6mm×250mm Inertsil ODS-P 液相色谱柱;流动相乙腈:水为60:40,初始流速为1mL/min,柱温为40 ℃;检测时间为40min,进样量为20μL;检测系统采用紫外检测器,开启波长切换模式,波长分别为205 和290nm.
1.9 数据处理与统计分析
采用Excel 2021 进行数据处理,使用Origin 2021 绘图与降解动力学数据拟合.各组之间的数据差异采用ANOVA 分析比较(SPSS 23.0 软件).
2 结果与讨论
2.1 菌群生长曲线
如图1(a)所示,根据细菌生长周期规律来划分,菌群接种到LB 培养基中后,在培养前期生长缓慢,处于延滞期,菌量增幅较小.大概经过4h 后菌群进入对数生长期,菌体数量开始快速增加,10h 时菌群达到生长高峰,之后进入稳定期,此时细菌的增殖数与死亡数基本持平.处于对数生长期的菌群生长繁殖速度最快,代谢能力强,拥有最好的降解能力[16].同时,该菌群相比某些菌群繁殖速度较快[17].
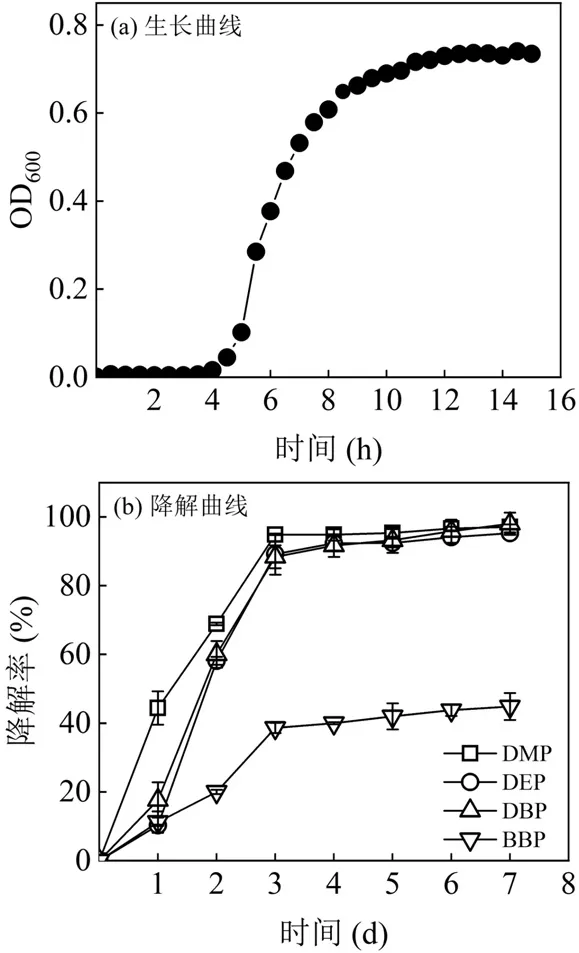
图1 菌群的生长和降解曲线Fig.1 Growth and degradation curves of bacterial consortium
2.2 菌群对PAEs 的降解效能
纯培养体系下该菌群可以有效降解 4 种USEPA 优先控制PAEs.如图1(b)所示,菌群在添加PAEs(5mg/L)的无机盐液体培养基中培养1d 时,对4种 PAEs 的降解率都比较低,分别为 44.41%、10.27%、17.65%和11.23%.第3d 时,菌群对DMP、DEP、DBP、BBP 的降解率相比第1d 有了较大提高,分别达到了95.86%、93.89%、88.37%和38.61%.随着培养时间的延长,PAEs的降解率维持平稳,第5d时对DMP、DEP、DBP、BBP 的降解率分别为96.33%、94.41%、93.20%和41.96%,第7d 时对4种PAEs 的降解率分别为97.08%、94.47%、98.02%和44.82%.
利用一级动力学方程对PAEs 降解曲线进行拟合,如图2 和表1 所示,菌群对4 种PAEs 的降解均符合一级反应动力学.
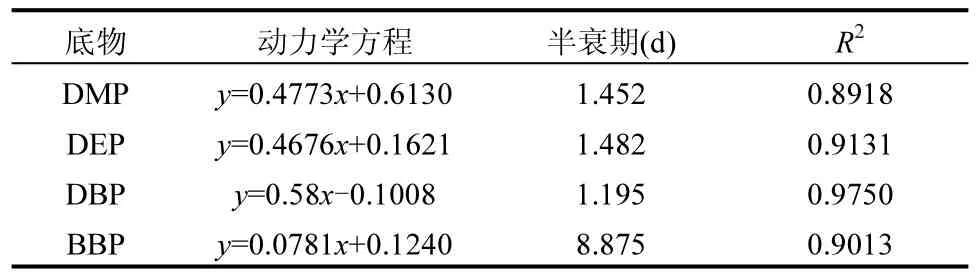
表1 PAEs 的降解动力学方程Table 1 Degradation kinetic equation of PAEs

图2 菌群降解PAEs 的准一级动力学拟合曲线Fig.2 Pseudo-first-order kinetic fitting curves for the degradation of PAEs by bacterial consortium
由于DMP、DEP、DBP 结构较简单,具有更高的生物利用率,在降解过程中更趋于被优先利用,在降解起始的第7d,3 种PAEs 基本上被完全降解.相比之下,分子量较大的BBP 降解进展比较缓慢,降解半衰期远大于其余3 种PAEs.可能是因为该菌群在PAEs 的代谢途径上不能互补,造成某些中间产物无法继续代谢,从而产生对菌群的抑制作用[18].
与单一菌株相比,菌群内部的各个菌株为降解过程提供了更多的降解基因和代谢途径,使得菌群具有更高的污染物矿化能力[19].近年来,也有许多研究者从环境中富集驯化出具有PAEs 降解能力的菌群.吕律[20]从活性污泥中富集培养得到一组菌群LV-1,该菌群对DBP 的降解率可达到95%以上;李静[21]从污泥中富集驯化的具有PAEs 降解能力的菌群SD-1 对DMP、DEP 和DBP 均具备降解能力,在72h 内的降解率分别为68.71%、68.46%和80%.相比之下本研究筛选出的菌群对PAEs 具备较高的降解广谱性和高效性,在降解环境中多种共存的PAEs时具备一定优势,应用潜力较大.
2.3 菌群的群落结构
借助16SrRNA 高通量测序技术分析了菌群的群落组成.结果(图3)显示,从门分类水平看,菌群中的最优势菌门为变形菌门(Proteobacteria,76.57%),其次为拟杆菌门(Bacteroidetes,21.04%)、放线菌门(Actinobacteria,2.37%).从属分类水平看,菌群主要由鞘氨醇杆菌属(Sphingobacterium,33.03%)、代尔夫特菌属(Delftia,40.61%)、假单胞菌属(Pseudomonas,11.70%)、无色杆菌属(Achromobacter,3.04%)和根瘤菌属(Rhizobium)(6.90%)组成.其中,鞘氨醇杆菌属(Sphingobacterium)和代尔夫特菌属(Delftia)占比较高,此两种菌属已被多次报道具备PAEs 降解的能力.Neelakanteshwar 等[22]分离出一株 Delftia sp.TBKNP-05,该菌株在培养基中经过5d 的培养后对DBP(10mmol/L)的降解率可达到100%;马永见[23]从土壤中分离出一株Sphingobacterium sp. MJ1,该菌株在 780mg/L PAEs(DMP、DBP、DEHP 各为260mg/L)的无机盐培养基中培养3d 后对DBP 和DMP 的去除率均可达到91%以上,对DEHP 的去除率也可达到82%以上.其余几种菌属同样具有PAEs降解能力,如Feng 等[24]筛选出的一株Pseudomonas sp. YJB6 经过3d 的培养可完全降解200mg/L 的DBP;Wang 等[25]筛选出一株Achromobacter sp. RX,经过96h的培养,对DEHP的降解率可达97.9%;Tang等[26]筛选出一株Rhizobium sp. LMB-1,45h 内可完全降解100mg/L 的DMP.本研究富集驯化获得的菌群中的各占比较高菌属均具备PAEs 降解能力,且各个菌属之间可能存在协同代谢作用,从而增强菌群的降解广谱性和高效性[27].值得注意的是,鞘氨醇杆菌属(Sphingobacterium)和代尔夫特菌属(Delftia)在菌群中的占比高达73.64%,推测PAEs的降解过程可能由这2 种优势菌属主导.
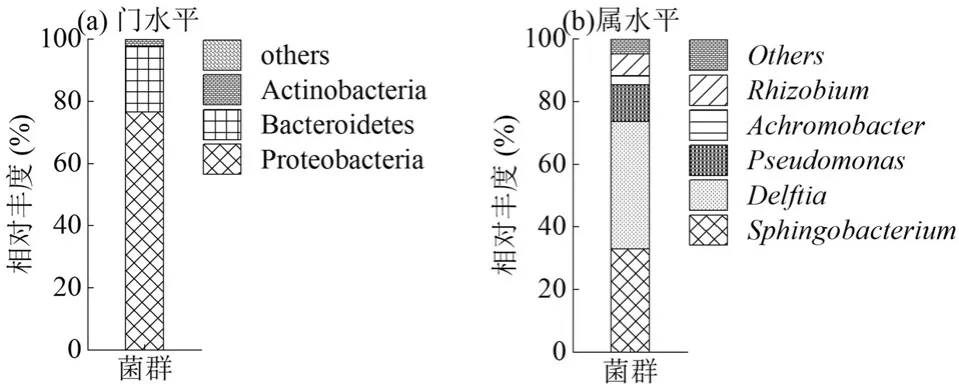
图3 菌群的群落结构Fig.3 Bacterial community structure of the bacterial consortium
2.4 环境条件对菌群降解PAEs 的影响
2.4.1 pH值 pH值是影响微生物降解能力的重要环境因素之一.过高或者过低的pH 值均会影响微生物及其相关降解酶的活性.由图4(a)可以看出,菌群对PAEs 的最大降解率主要集中在pH 7~8 的范围内,降解率分别达78.30%和76.60%,在此pH 值范围内,菌群对PAEs 的降解率无明显差异.过酸或过碱条件会改变生物细胞膜的选择透过性从而影响其对营养物质的吸收或改变相关降解酶活性.柴阳阳等[28]筛选出了具有DBP 降解能力的内生菌株HBT4,发现适合菌株培养的pH 值为6~8,pH 值为4 时菌株对DBP 的降解率不足10%.尽管本研究筛选的菌群在不同pH 值条件下对PAEs 的降解率存在一定差异,但整体来看菌群在pH 5~9 的范围内对PAEs 均具备良好的降解效果,相比于一些单菌来讲,该菌群降解谱较广的同时还具备一定的耐酸碱性.

图4 环境条件对菌群降解PAEs 的影响Fig.4 Effect of environmental conditions on the degradation by the bacterial consortium
2.4.2 温度 温度会影响微生物相关降解酶的活性从而影响菌体对污染物的降解能力.由图4(b)可以看出,菌群对PAEs 的最大降解率主要集中在温度 25~30℃的范围内,降解率分别达 85.14%和77.91%.此温度范围内,菌群对PAEs 的降解率无明显差异.当温度在20℃和35~ 40℃范围内时,PAEs降解率与25℃时相比显著降低.李方方等[29]发现当培养体系温度为20℃时,菌群LV-1 对DBP 的降解率仅在40%左右,在25~35℃范围内时,菌群对PAEs的降解率较高,可达到80%以上.可见温度过高或者过低均会对菌群的降解能力产生影响.较低的温度会使细胞膜的流动性变差,PAEs 的溶解度变低,阻碍微生物对营养物质和养分的吸收;较高的温度则会使细胞内相关酶的活性降低和核酸变性,使PAEs 降解率变低[30].相比之下,本研究筛选的菌群在较高或较低的温度条件下对PAEs 的降解率仍可以达到60%以上,在不同温度环境下降解PAEs 具备一定的优势.
2.4.3 盐度 盐度是影响菌群降解PAEs 的关键因素之一.由图4(c)可见,不同盐度条件下菌群降解PAEs 的能力差异较明显.随着盐度的增加,菌群对PAEs的降解率逐渐降低,在1%~2%范围内降解效果较好.当盐度在8%时,PAEs 降解率仅28.59%.周婷等[31]在研究盐度条件对菌群ZM 降解DMP 的影响时发现,当盐度增加到4%时,菌群对DMP 的降解率几乎为0.可能的原因是高盐度会使蛋白质发生变性,致使酶的活性受到抑制[32],同时高盐度会使微生物耗氧速率变低,导致PAEs 降解率下降[33].对比之前的研究,本研究筛选的菌群在盐度为4%时仍具备比较好的PAEs 降解能力,说明其对PAEs 降解具备较高的耐盐性和广泛的盐度范围.
2.4.4 底物浓度 底物浓度对菌群降解PAEs 的影响如图4(d)所示.菌群对PAEs 的最大降解率主要集中在 5~10mg/L,降解率最大可达 76.73%(5mg/L).当浓度在2mg/L 和15~20mg/L 时,菌群对PAEs 的降解率显著降低.徐莹[34]在研究菌株FA1对芘的降解特性时发现了类似的现象.可见底物浓度过高或者过低均会影响菌群的降解活性.这可能是由于底物浓度过低不容易引起菌群相关降解基因的表达,使其不能将底物充分利用;浓度过高则会对微生物产生毒性,影响微生物的生长和降解活性[35].
2.5 功能内生菌群定殖效能
功能内生细菌相较于其他菌株,能够更好地适应植物体内的环境,与宿主植物形成良好的共生体系,发挥良好的降解效能.本研究中富集驯化的功能内生菌群定殖后可以显著降低水稻体内的PAEs 含量(表2).相同培养周期内,ZR 组水稻体内DMP、DEP、DBP 和∑PAEs 的含量与ZK 组相比分别降低了41.09%、45.33%、63.06%和32.30%,BBP 含量无明显变化.Xu 等[36]将内生菌株Bacillus subtilis strain HB-T2 定殖到卷心菜体内,发现定殖HB-T2 的卷心菜根、茎中DBP 的残留浓度分别为未接种HB-T2卷心菜相应组织的29.3%和52.4%.

表2 水培15d 后水稻体内PAEs 的含量Table 2 Contents of PAEs in rice after 15 days of hydroponic culture
功能内生细菌定殖到植物体内后,一方面会使植物体内相关降解酶系活性发生变化进而促进植物体内有机污染物的去除.陈学斌等[37]研究发现,玉米接种菌株XB 能显著提高玉米体内超氧化物歧化酶、过氧化氢酶及多酚氧化酶活性,缓解土壤DEHP对玉米生长的胁迫,降低玉米吸收积累DEHP;另一方面,功能内生细菌可以增加植物体内降解基因丰度.Zhang 等[38]将功能内生菌群CEB 定殖到植物体内,增加了植物体内PAHs 代谢相关功能基因的拷贝数,促进了植物体内PAHs 的去除.此外,有研究指出功能内生细菌可以以植物体内的氨基酸、葡萄糖等内源性有机物为碳源和能源物质,提高自身的共代谢能力以促进植物体内污染物的去除[39].因此,定殖功能内生细菌有望降低作物体内PAEs 的积累.对比以往针对单一PAEs 的研究,本研究筛选获得的菌群定殖植物后能够有效去除植物体内3 种PAEs,在实现实际污染区作物体内PAEs 的消减方面具备一定优势,在保障PAEs 污染区农产品质量安全和人体健康方面具有较广阔的应用前景.
此外,菌群的定殖促进了水稻的生长,提高了植株生物量(表3).定殖的促生效应在根长、株高、鲜重、干重上均比较显著.定殖后根长和株高分别提高了13.01%和28.63%,鲜重和干重分别提高了14.91%和20.83%.功能内生菌群主要通过以下两个途径促进植物生长:一方面,内生细菌可以通过固氮、磷酸盐溶解和铁螯合等作用,促进植物对营养物质的吸收[40];另一方面,部分内生细菌还可以通过调节激素水平来促进植物生长[41].

表3 水培15d 后水稻的生物量Table 3 Biomass of rice after 15days of hydroponic incubation
3 结论
3.1 本研究富集驯化了一组具有降解4 种USEPA优控PAEs 能力的功能内生菌群,纯培养体系中,7d内该菌群对DMP、DEP、DBP、BBP 的降解率分别达到97.08%、94.47%、98.02%和44.82%.
3.2 菌群在门分类水平上主要由变形菌门(Proteobacteria,76.57%)、拟杆菌门(Bacteroidetes,21.04%)和放线菌门(Actinobacteria,2.37%)组成;在属分类水平上主要由鞘氨醇杆菌属(Sphingobacterium, 33.03%)、代尔夫特菌属(Delftia,40.61%)、假单胞菌属(Pseudomonas, 11.70%)、无色杆菌属(Achromobacter,3.04%) 和根瘤菌属(Rhizobium, 6.90%)组成.
3.3 菌群的最佳降解条件为:pH 7、25℃、盐度1%、底物浓度5mg/L.
3.4 水培条件下,以浸根的方式将菌群定殖到水稻体内可以有效降低植物中PAEs 的含量,15d 内DMP、DEP、DBP 和∑PAEs 的去除率分别达到41.09%、45.33%、63.06%和32.3%,同时该菌群还能够促进作物的生长,提高作物产量.
