甘蓝型油菜373S温敏不育基因在可育细胞质背景下的表达
2024-03-28王静张东锁董慧孙妍妍王珍珍郭媛胡胜武
王静 张东锁 董慧 孙妍妍 王珍珍 郭媛 胡胜武

摘 要 旨在明确甘蓝型油菜温敏不育系373S温敏不育基因表达与特定细胞质之间的关系。以甘蓝型油菜373S(微粉)为父本,以具有cam细胞质的Shaan 2B为母本,杂交、回交及自交得到BC1及F2代群体。结果表明,在上述具有cam细胞质的BC1及F2中,可育株与不育株的分离比分别符合3∶1与15∶1,表明该群体中不育性状受2对基因控制。分离群体中的不育株表现出温敏不育特征,其不育花形态与373S没有明显差别;与373S比较,该分离群体中的不育植株结实角果数及自交结实率有不同程度的降低,不育期时间率(不育期时间所占百分比)无显著差异;花冠直径、四强花药长度、二弱花药长度无明显差异,花柄长度、花瓣长度、花瓣宽度、四强雄蕊长度、二弱雄蕊长度略有增加。细胞学观察表明,该分离群体中的不育株小孢子的发育过程、染色体形态、胼胝质沉积与373S无明显差异,均在花粉母细胞时期出现异常。本研究成功地将373S温敏不育基因从pol细胞质导入cam细胞质,说明该温敏不育性状是由细胞核内温敏不育基因导致,与细胞质背景无关。
关键词 甘蓝型油菜(Brassica napus L.);温敏基因;细胞质类型;雄性不育
油菜是中国乃至世界重要的油料作物,是人类食用植物油的重要来源,也是动物饲料中蛋白质的重要来源。中国油菜常年播种面积700万hm2,总产超过1 400万t,可生产优质食用油约550万t,约占国产植物油50%;菜籽榨油后可获得高蛋白饲用菜粕约800万t。在油菜全价值链挖掘和全产业链开发中,还可以充分挖掘其菜用、蜜用、饲用、肥用、休闲观光等价值[1]。2017年全国消费植物油总量为3 565万t,其中,自产植物油为1 100万t,自给率仅为30.8%,国产油料难以满足消费需求,供需缺口与日俱增[2]。因此,大力发展油菜产业对于中国食用植物油安全供给、增加菜农收入、实现乡村振兴战略具有非常重要的意义。油菜杂种优势显著,利用杂种优势是显著提高油菜产量的重要途径之一。目前,油菜杂种优势利用的主要途径有细胞质雄性不育(Cytoplasmic male sterility,CMS)、细胞核雄性不育(Genic male sterility,GMS)、生态型雄性不育(Ecotype male sterility,EMS)、化学杂交剂诱导的雄性不育(Chemical hybridization agent (CHA)-induced male sterility,CIMS)和自交不亲和(Self-incompatibility,SI)等[3]。生态型(光温敏)雄性不育是一种特殊的不育系类型,其育性由自身基因和外界环境条件共同决定,特定生态条件下既可作为不育系进行制种,又可作为保持系自交繁殖,真正实现一系两用。生态型雄性不育具有恢复谱广、无细胞质负效应、配组自由、易选配出强优势组合等优势,在油菜杂种优势利用中受到广泛关注和研究。中国科学家在油菜光温敏雄性不育材料的发掘及利用方面做了大量先驱工作,发现了一些光温敏雄性不育材料,选育出湘杂油系列、赣两油系列、华油杂系列等两系杂交种,预示油菜两系杂交种发展前景广阔。目前,利用生态型雄性不育材料选育出一系列油菜杂交种,例如,利用湘油91S配制出的‘湘杂油5号[4-5],利用其衍生系湘油402S配制出的‘湘杂油7号[6-7];利用501-8S配制出的‘两优586[8-9],利用其衍生不育系选育出‘赣两优2号‘赣两优3号‘赣两优5号‘秀油杂610等品种[10-13]。此外,报道的光温敏雄性不育两用系还有湘油104S[14]、温敏不育系533S及其衍生系417S[15-16]、H50S[17]、160S[18]、SP2S[19]、TE5A[20]、K121S[21]等;光温敏互作不育系有H90S、N196S、遵矮S等[22-24]。
甘蓝型油菜生态型雄性不育系373S是由西北农林科技大学油菜研究中心(以下称本课题组)发现的自然突变不育株,经多年选择育成[25]。前期研究表明,373S的育性主要受温度条件影响,表现为低温可育、高温不育,育性转换的临界值在10~12 ℃;细胞学研究结果表明,373S小孢子败育起始于花粉母细胞时期;遗传研究表明,373S的不育性状受1对隐性核基因Bnmst1控制。分子鉴定结果表明,373S具有pol细胞质,其不育性状遗传机制与国内油菜杂交种育种中广泛利用的pol CMS不同[26-27]。那么,373S温敏不育基因的表达是否与pol细胞质背景有关?为回答这一科学问题,本研究以373S微量花粉为父本,以具有可育细胞质cam的甘蓝型油菜为母本,将373S的温敏不育基因通过杂交等手段,导入cam细胞质背景中,对其后代群体的不育株进行花器官形态观察、育性调查、细胞学观察以及遗传研究,旨在明确373S温敏不育基因与特定油菜细胞质之间是否存在互作关系。
1 材料与方法
1.1 试验材料
甘蓝型油菜生态型雄性不育系373S、Shaan 2B、ZS9 3个自交系。在细胞质类型分子检测结果的基础上,以具有cam细胞质类型的Shaan 2B为母本,373S为父本,进行杂交获得F1代,F1代自交獲得F2代,F1代与373S回交获得BC1代。以上材料于2020年9月播种于陕西杨凌西北农林科技大学杨凌试验基地,一般大田管理,小区 种植。
1.2 试验方法
1.2.1 细胞质类型鉴定 采取373S,以及Shaan 2B 与373S杂交衍生的具有cam细胞质背景的BC1代及F2代群体中不育株的幼嫩叶片,利用改良CTAB方法[28]提取基因组DNA。采用一步多重PCR方法鉴定所取材料的细胞质类型,所用3对PCR引物信息及试验方法见Zhao等[29]文献报道。
1.2.2 植株形态观察及育性统计 2021年盛花期,调查统计Shaan 2B×373S衍生的具有cam细胞质背景的BC1及F2群体中可育株与不育株(≥5朵不育花的单株[27])情况。上述群体中,可育株与不育株的分离比率利用χ2检验,确定该群体中不育性状的遗传行为。
在上述BC1及F2群体中,各随机选取10株不育株,每个不育株随机选取20朵花,对其花冠直径、花柄长度、花瓣长度、花瓣宽度、四强雄蕊长度、二弱雄蕊长度、四强花药长度、二弱花药长度等指标进行统计;记载其总花期时间、不育期时间、计算不育时间率(不育期时间所占百分比),不育时间率=不育期时间/总花期时间×100;不育植株套袋自交,统计自交结实率,自交结实率= 自交结实角果数/自交花朵总数;同时,对不育植株花序、花器官形态特征进行拍照记载。
1.2.3 细胞学观察 花期选取ZS9、373S、Shaan 2B×373S衍生的BC1、F2群体中不育株的主花序,利用卡诺固定液[V(无水乙醇)∶V(冰乙 酸)=3∶1] 固定12~24 h,经梯度乙醇(100%,95%,85%,75%)脱水各2 h后,置于70%乙醇中保存备用。取固定材料的雄蕊,利用1%醋酸洋红染液染色压片观察;苯胺蓝染液(DAPI染液)染色压片,进行染色体及胼胝质的观察。细胞学观察结果在OLYMPUS BX51(奥林巴斯,日本)荧光显微镜下拍照记录。
1.3 数据处理
利用Excel 2013对试驗数据进行整理,SPSS 24.0对群体育性数据进行χ2检验,且对指标数据进行统计分析,差异显著性分析使用Duncans法,P<0.05。
2 结果与分析
2.1 细胞质类型鉴定结果
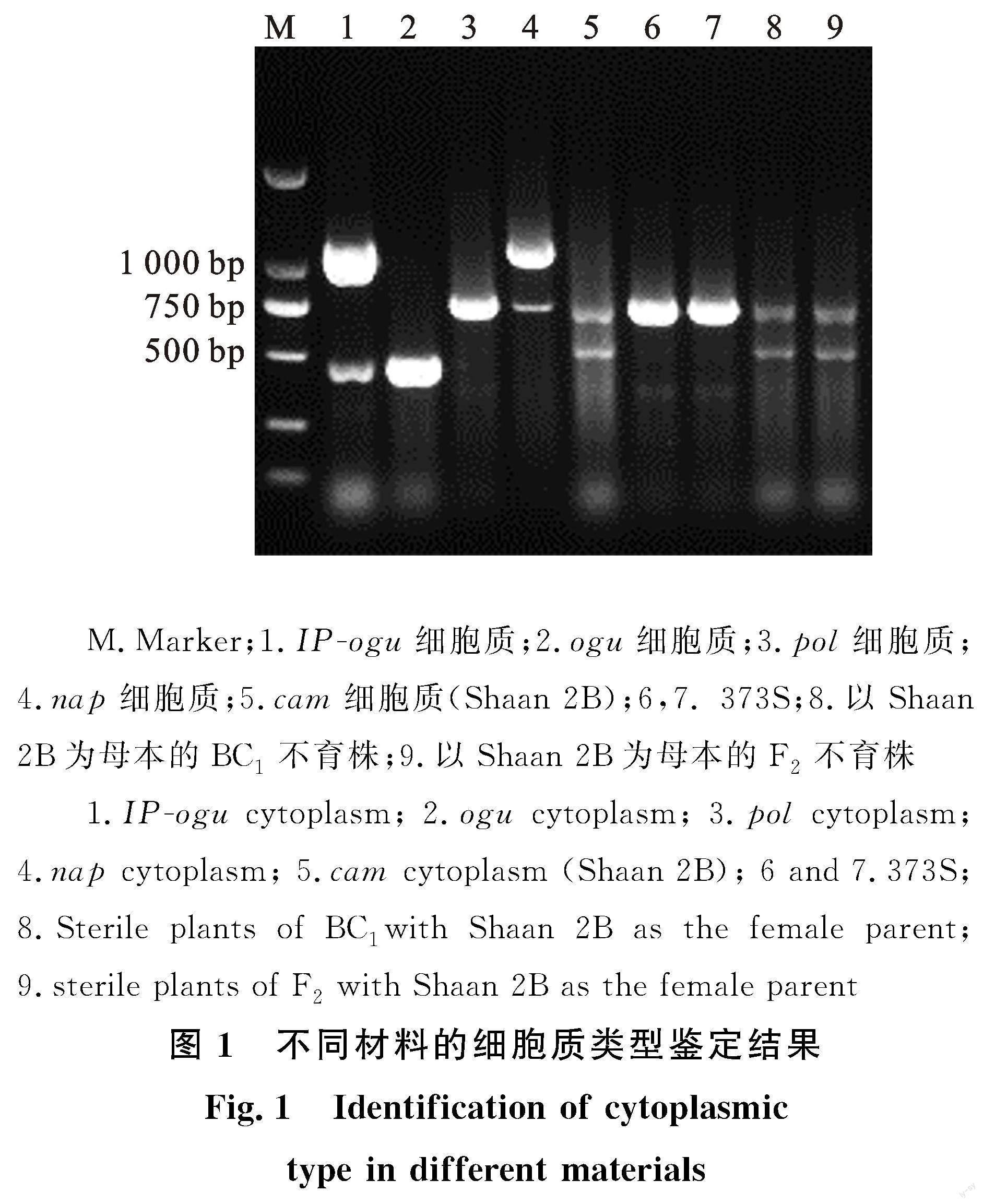
为确定待测材料的细胞质类型,利用本课题组前期研发的一步多重PCR方法[30]对参试材料的细胞质类型进行鉴定。结果表明(图1),不同的细胞质类型(IP-ogu、ogu、pol、nap、cam)扩增出特定的细胞质类型条带,与Zhao等[29]报道结果相符。其中,373S扩增出747 bp条带,为pol细胞质类型。Shaan 2B扩增500 bp和800 bp两条条带,这两条条带亮度基本一致,为cam细胞质类型。以Shaan 2B为母本,以373S不育株(微粉)为父本,杂交得到的BC1及F2群体中不育株,扩增结果与Shaan 2B带型一致,为cam细胞质类型。
2.2 育性遗传
对Shaan 2B×373S衍生的F1、BC1及F2群体进行育性调查(表1),结果显示,F1植株全部表现为可育,而在总共87株的BC1群体中,可育株63株,不育株24株,分离比符合3∶1(P> 0.05),在总共406株的F2群体中,可育株386株,不育株20株,分离比符合15∶1(P>0.05)。育性分离比结果说明,由373S衍生的cam细胞质类型的群体中,其不育性状受2对隐性基因控制。结果表明,373S的温敏基因已成功导入cam细胞质类型内,373S温敏不育基因的表达与特定细胞质背景(pol)无关。
2.3 不同细胞质背景下的不育植株形态学观察及单花性状调查
观察具有pol细胞质背景的373S植株,以及Shaan 2B×373S衍生的具有cam细胞质背景的BC1及F2群体中不育植株的育性变化、单花形态和花粉活力情况,结果发现三者初花期时,同一花序底端所开花均为不育花,雄蕊退化,无花粉或微量花粉;随着田间环境条件的转变,花序顶端新开放的花朵育性开始转变为可育状态,花药形态饱满,正常开裂,释放花粉,期间,有多级育性花并存。两种不同细胞质背景的生态不育株,植株花的育性均随田间环境条件的变化而发生转变,同一花序中出现可育花和不育花并存的现象,育性表现为环境敏感不育(图2-A1~C1)。对其不育花单花观察发现,其单花形态没有明显差异。花器官外形总体来看,萼片、花瓣、雌蕊发育均正常,但雄蕊退化,导致不育花花冠正面均看不到雄蕊,花丝长度明显变短,花药干瘪萎缩,高度不及柱头(图2-A2~C2)。分别对其不育花花粉活力进行测定,发现其花粉育性表现为彻底不育(图2- A3~C3)。
为探究其不育期时间、结实率及花器官性状之间是否存在差异,对所取不育材料进行育性统计调查及花器官性状调查。结果表明,373S、Shaan 2B×373S衍生的BC1及F2群体中不育株的不育期时间与花期总时间、不育时间率无显著差异。结实率方面,与373S相比,Shaan 2B×373S衍生的BC1及F2群体中,不育株的自交结实率有所降低(表2)。花器官性状调查发现(表3),除花冠直径、四强花药长度、二弱花药长度外,Shaan 2B×373S衍生的BC1及F2群体中不育株的其他5个指标(花柄长度、花瓣长度、花瓣宽度、四强雄蕊长度、二弱雄蕊长度)比373S显著增加。
2.4 不同细胞质背景下的不育株花药败育的细胞学观察
通过对不同细胞质背景下的油菜温敏雄性不育材料小孢子发育观察,研究其小孢子在主要细胞学发育时期是否存在差异。结果表明,与可育植株(图3-F1)相比,373S及其衍生的不育植株均在花粉母细胞时期出现异常(图3-A1~C1),可育花药的花粉母细胞周围被圆润的胼胝质层包裹,而不育花药周围胼胝质稀薄,但均能进行减数分裂且形成四分体结构。四分体时期,373S及其衍生的cam细胞质背景的不育花药周围胼胝质均明显薄于可育花药(图3-F2~C2)。随后,胼胝质解体,小孢子释放,此时可育小孢子与不育小孢子形态出现明显异常,可育小孢子花粉壁形成,外形圆润(图3-F3),而不同细胞质背景下的不育小孢子花粉外壁形状异常且不规则,出现椭圆形、多边形等异常形状(图3-A3~C3)。之后,小孢子进行有丝分裂,可育小孢子体积逐步增大,而不育小孢子的体积无明显变化(图3-F3~C3)。成熟期,可育花粉粒细胞质分布均匀,形态饱满,而不育花粉粒只剩花粉空壳,花粉败育(图3-F5~C5)。在花药发育的整个时期内,不论是pol细胞质背景的373S,还是其衍生出的cam细胞质背景的不育植株,在相同时期内,其小孢子发育形态等无明显差异。
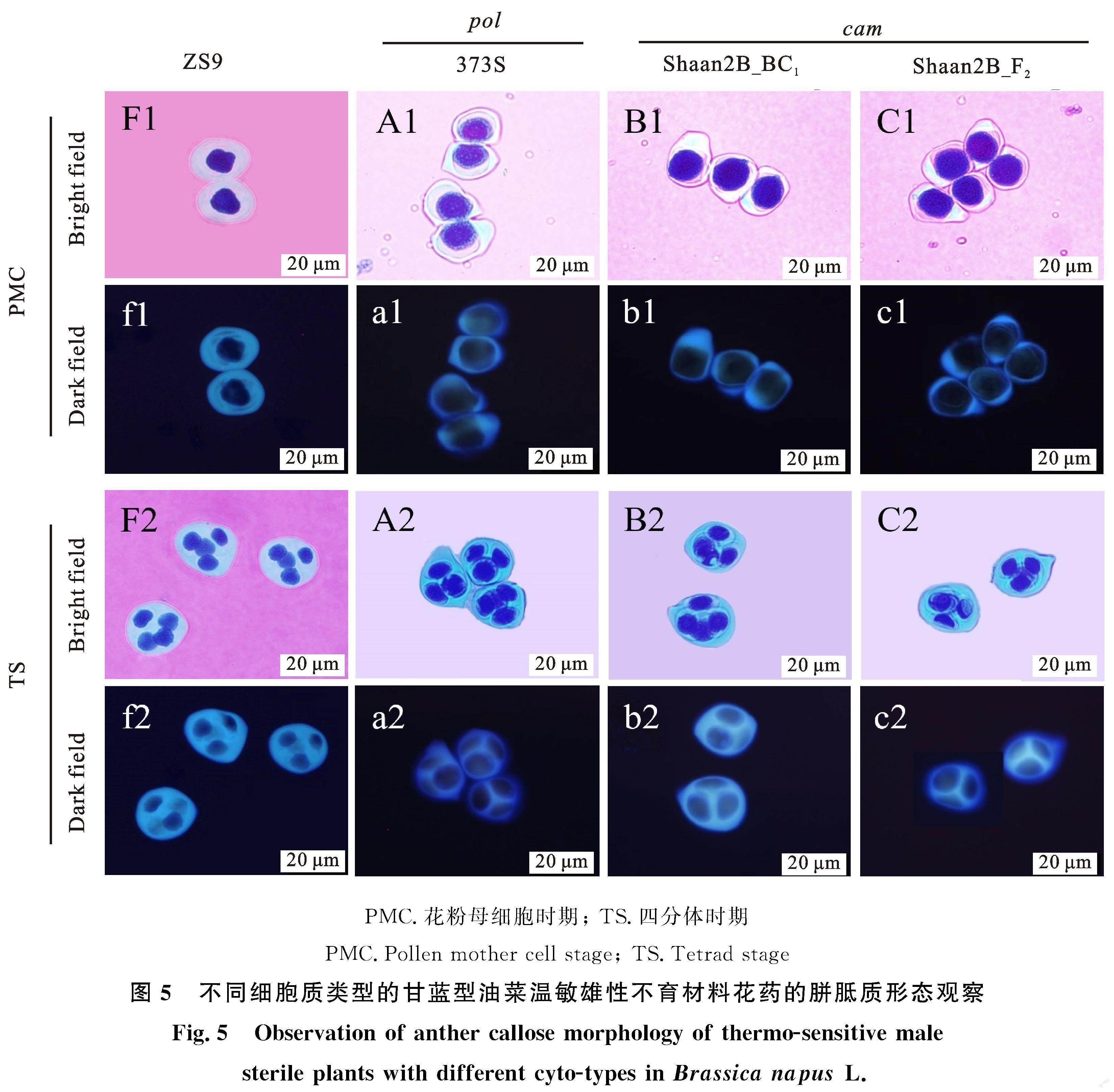
为探究花药发育期间核内染色体形态是否存在差异,对所取材料的花药进行DAPI染色观察。结果表明,与可育花药相比,两种不同细胞质背景下的不育株,在花粉母细胞减数分裂期,核内染色体出现异常,Shaan 2B×373S衍生的BC1及F2群体中不育株花药减数分裂不同步(图4-B1,4-C1),373S不育花药呈团状且黏连状态(图4-A1)。四分体时期,pol细胞质背景373S及衍生的cam细胞质背景的BC1代及F2代中不育株的不育花药其四分体荧光信号均模糊,与可育花药清晰的荧光信号成明显对比(图4-F2~C2)。成熟花粉粒时期,可育花粉粒核内染色体荧光信号强烈,而不育花粉粒均无荧光信号(图4-F3~C3)。对其胼胝质沉积模式观察发现,可育花药的胼胝质圆润清晰,不育花药其荧光信号在花粉母细胞时期便出现模糊现象(图5-F1~c1)。四分体时期,不育花药的四分体体积均小于可育花药(图5-F2~C2),其胼胝质厚度也明显薄于可育四分体,但在373S及衍生的cam细胞质背景的不育花药之间,其发育大小及胼胝质厚度无明显差异(图5-f2~c2)。
3 讨论与结论
利用杂种优势可提高作物产量品质,是作物育种的重要途径。生态型雄性不育材料的发现,促使作物杂种优势利用由“三系”向“两系”转变,因其育性受自身基因型和外界环境因素共同调控,其遗传机制也复杂多样。研究表明,油菜生态不育系501-8S[30]、104S[14]、Huiyou50S[31]、TE5A[20]等均属于单基因遗传模式;417S[32]、SP2S[19]、100S[33]其育性则由2对基因控制,属于双基因遗传模式;3基因及以上遗传模式则包括湘油91S、H90S、芥菜型油菜温敏核不育系K121S[34-36];甘蓝型油菜细胞质生态雄性不育两用系AB1[37]及其衍生系8-8112AB[38]等则属于质量-数量遗传。本课题组前期对373S的遗传研究表明,其育性受1对隐性核基因控制,且具有pol细胞质背景[27-28]。本研究以373S(微粉)为父本,通过杂交手段尝试将其温敏不育基因导入可育细胞质cam背景中,以此探究该温敏不育基因的表达与细胞质背景之间的关系。结果表明,由373S衍生的cam细胞质类型的BC1及F2群体中,均有不育株出现,且表现为温敏不育特性,表明373S温敏不育基因已成功导入cam细胞质背景中,且能够正常表达,从而使植株表现为温敏
不育性状。但对BC1群体及F2群体中,可育株与不育株的分离比的卡方检验表明,373S衍生的cam细胞质类型的后代其不育性状受2对独立基因控制,表现为隐性,这与前期研究的373S不育性状受1对隐性基因控制的结果有差异。推测出现这种现象的原因,可能是不同的油菜亲本与373S之间存在的育性基因数目有差异,在一些亲本中仅有1对基因差异,在另外一些亲本中存在2对基因差异。笔者正在对出现这种遗传行为的原因,进行经典遗传学和分子标记验证。
对生态型雄性不育材料进行细胞学观察,能够发现其败育时期及特征,有助于研究揭示其败育机理。前人研究表明,反型生态不育系533S无法分化出正常的花粉囊,属于孢原细胞败育型[39]。SP2S、TE5A由于减数分裂时期胼胝质降解滞后导致不育,属于花粉母细胞败育型,其中SP2S还出现绒毡层细胞空泡化现象[19],但TE5A的绒毡层发育正常[40]。生態型雄性不育系H50S、反型温敏不育系Huiyou50S均在单核时期出现异常现象,其中,H50S在此时期绒毡层降解缓慢,细胞核呈现解体[17],而Huiyou50S其绒毡层在单核后期迅速降解,两者均属于小孢子发育时期败育型[41]。本研究发现,373S中不育基因在不同细胞质背景下的不育株花药均在花粉母细胞时期出现异常现象。由373S衍生出的cam细胞质类型的不育株其核内染色体在花粉母细胞时期出现降解现象,减数分裂不同步、胼胝质层稀薄等;在小孢子时期,其花粉外壁不规则,体积不增大,直至败育。
本研究结果表明,调控373S育性的温敏基因在不同细胞质背景下表达,不同细胞质温敏不育材料间小孢子发育行为无明显差异,温敏基因的表达与pol细胞质背景无关。该研究结果将为分离克隆油菜373S温敏不育基因、明确其雄性不育机理及育种利用奠定基础。
参考文献 Reference:
[1] 王汉中.以新需求为导向的油菜产业发展战略[J].中国油料作物学报,2018,40(5):613-617.
WANG H ZH.New-demeand oriented oilseed rape industry developing strategy[J].Chinese Journal of Oil Crop Sciences,2018,40(5):613-617.
[2] 刘 成,冯中朝,肖唐华,等.我国油菜产业发展现状、潜力及对策[J].中国油料作物学报,2019,41(4):485-489.
LIU CH,FENG ZH ZH,XIAO T H,et al.Development,potential and adaptation of Chinese rapeseed industry[J].Chinese Journal of Oil Crop Sciences,2019,41(4):485-489.
[3] 傅廷栋,沈金雄,易 斌.杂交油菜的遗传与育种[M].武汉:湖北科学技术出版社:2019.
FU T D,SHEN J X,YI B.Breeding and Utilization of Rapeseed[M].Wuhan:Hubei Science & Technology Press,2019.
[4] 席代汶,陈卫江,宁祖良.甘蓝型油菜温敏核不育系‘湘91S的选育[J].湖南农业科学,1994(4):17-18.
XI D W,CHEN W J,NING Z L,et al.Breeding of thermo-sensitive genic male-sterile line ‘Xiang91S in Brassica napus[J].Hunan Agricultural Sciences,1994(4):17-18.
[5] 席代汶,邬贤梦,宁祖良,等.优质两系杂交油菜湘杂油5号的选育[J].中国油料作物学报,2005,27(1):23-25.
XI D W,WU X M,NING Z L,et al.Breeding of a two-linehybrid variety Xiangzayou 5 with double-low quality in Brassica napus L.[J].Chinese Journal of Oil Crop Sciences,2005,27(1);23-25.
[6] 邬贤梦,席代汶,宁祖良,等.甘蓝型油菜温敏核不育系湘油402S的选育[J].中国油料作物学报,2005,27(3):74-76.
WU X M,XI D W,NING Z L,et al.Breeding of thermo-sensitive genic male-sterile line Xiangyou 402S in Brassica napus[J].Chinese Journal of Oil Crop Sciences,2005, 27(3):74-76.
[7] 邬贤梦,宁祖良,黄虎兰,等.优质杂交油菜新品种湘杂油7号的选育[J].湖南农业科学,2005(5):37-38.
WU X M,NING Z L,HUANG H L,et al.Breeding of high quality rape variety Xiangzayou No.7[J].Hunan Agricultural Sciences,2005(5):37-38.
[8] 刘尊文,彭芝兰,周小平,等.甘蓝型油菜光温敏雄性不育两用系501-8S的选育[J].中国油料作物学报,1998,20(1):13-16.
LIU Z W,PENG ZH L,ZHOU X P,et al.Breeding of GMS two-line system 501-8S sensitive to temperature and photoperiod in Brassica napus[J].Chinese Journal of Oil Crop Sciences,1998,20(1):13-16.
[9] 刘尊文,袁卫红,李文信,等.甘蓝型两系杂交油菜两优586的选育[J].中国油料作物学报,2000,22(2):5-7.
LIU Z W,YUAN W H,LI W X,et al.Breeding of two-line system hybrid Liangyou 586 in Brassica napus[J].Chinese Journal of Oil Crop Sciences,2000,22(2):5-7.
[10] [ZK(#]刘尊文,吴 平,张迁西,等.优质两系杂交油菜“赣两优二号”的选育[J].江西农业学报,2007,19(11):10-11.
LIU Z W,WU P,ZHANG Q X,et al.Breeding of high quality two-line hybrid rape variety Ganliangyou2[J].Acta Agriculturae Jiangxi,2007,19(11):10-11.
[11] 高海軍,刘尊文,袁卫红,等.甘蓝型两系杂交油菜赣两优3号的选育[J].湖北农业科学,2010,49(10):2370-2371.
GAO H J,LIU Z W,YUAN W H,et al.Breeding of two-line hybrid rape variety Ganliangyou No.3[J].Hubei Agricultural Sciences,2010,49(10):2370-2371.
[12] 刘 宁,吴 平,袁卫红,等.优质两系杂交油菜‘赣两优5号的选育[J].江西农业学报,2013,25(2):25-26.
LIU N,WU P,YUAN W H,et al.Breeding of high-quality two-line hybrid rape variety ‘Ganliangyou No.5[J].Acta Agriculturae Jiangxi,2013,25(2):25-26.
[13] 王小武,袁卫红,吴 平,等.甘蓝型早熟两系杂交油菜‘秀油杂610的选育[J].湖南农业科学,2016(4):14-15,18.
WANG X W,YUAN W H,WU P,et al.Breeding of precocious two-line hybrid rape variety ‘Xiuyouza 610 in Brassica napus[J].Hunan Agricultural Sciences,2016(4):14-15,18.
[14] 邬贤梦,肖 钢,官春云.甘蓝型油菜温敏核不育材料104S的发现及遗传特性分析[J].湖南农业大学学报(自然科学版),2009,35(6):597-601.
WU X M,XIAO G,GUAN CH Y.Discovery and genetic analysis of a thermo-sensitive genic ms material 104S in Brassica napus[J].Journal of Hunan Agricultural University (Natural Sciences),2009,35(6):597-601.
[15] 董军刚,董振生,刘创社,等.甘蓝型油菜生态型雄性不育两用系533st的选育[J].陕西农业科学,2001(7):8-9.
DONG J G,DONG ZH SH,LIU CH SH,et al.Breeding of GMS two-line system 533st in Brassica napus[J].Shaanxi Agricultural Sciences,2001(7):8-9.
[16] 董军刚,董振生,刘创社,等.甘蓝型油菜温敏不育系417S选育与鉴定[J].中国油料作物学报,2008,30(3):306-311.
DONG J G,DONG ZH SH,LIU CH SH,et al.Selection and identification of thermo-sensitive male sterile line 417S in Brassica napus L.[J].Chinese Journal of Oil Crop Sciences,2008,30(3):306-311.
[17] 孙晓敏,胡胜武,于澄宇.油菜生态不育系H50S花药败育的细胞学观察[J].西北农业学报,2009,18(5):153-158.
SUN X M,HU SH W,YU CH Y.Cytological observation of anther development of an ecological male sterile line H50S in Brassica napus L.[J].Acta Agriculturae Boreali-occidentalis Sinica,2009,18(5):153-158.
[18] 张 涛,沈亮余,王瑞雪,等.甘蓝型油菜雄性不育系160S育性转换与利用[J].西北植物学报,2012,32(1):35-41.
ZHANG T,SHEN L Y,WANG R X,et al.Fertility alteration and utilization of male-sterile line 160S in Brassica napus[J].Acta Botany Boreali-occidentalis Sinica,2012,32(1):35-41.
[19] YU C Y,GUO Y F,GE J,et al. Characterization of a new temperature-sensitive male sterile line SP2S in rapeseed (Brassica napusL.)[J].Euphytica,2015,206(2):473-485.
[20] ZENG X H,LI W P,WU Y H,et al. Fine mapping of a dominant thermo-sensitive genic male sterility gene (BntsMs) in rapeseed (Brassica napus) with AFLP- and Brassica rapa-derived PCR markers[J].Theoretical and Applied Genetics,2014,127(8):1733-1740.
[21] 李石開,刘其宁,吴学英,等.芥菜型油菜光温敏核不育系K121S的选育[J].中国油料作物学报,2002,24(3):1-5.
LI SH K,LIU Q N,WU X Y,et al.Breeding of photo-thermosensitive GMS line K121S in mustard(Brassica juncea)[J].Chinese Journal of Oil Crop Sciences,2002,24(3): 1-5.
[22] 王 华,赵继献,汤晓华.甘蓝型油菜新型细胞核雄性不育材料的选育[J].中国油料,1997,9(1):8-11.
WANG H,ZHAO J X,TANG X H.Selection and breeding of a new genetic male sterility type in B.napus L.[J].Chinese Oil,1997,9(1):8-11.
[23] 刘尊文,吴 平,袁卫红,等.甘蓝型油菜光温敏核不育两用系N196S的选育[J].江西农业大学学报,2006,28(5):654-667.
LIU Z W,WU P,YUAN W H,et al.Breeding of photo and temperature sensitive genic male sterile doub-use line N196S in Brassica napus[J].Acta Agriculturae Universitatis Jiangxiensis,2006,28(5):654-667.
[24] 苏振喜,邱怀珊,李石开,等.芥菜型油菜生态型雄性不育两用系研究[J].中国油料作物学报,1999,21(2):5-7.
SU ZH X,QIU H SH,LI SH K,et al.Preliminary studies on GMS two-line system in B.juncea[J].Chinese Journal of Oil Crop Sciences,1999,21(2):5-7.
[25] 于澄宇,李 玮,常建军,等.油菜温敏雄性不育系373S的选育[J].中国农学通报,2007,23(7):245-248.
YU CH Y,LI W,CHANG J J,et al.Development of a thermo-sensitive male-sterile line 373S in Brassica napus L.[J].Chinese Agricultural Science Bulletin,2007, 23(7):24-248.
[26] 孙妍妍.甘蓝型油菜生态不育系373S的温光特性、细胞学特征及遗传调控网络研究[D].陕西杨凌:西北农林科技大学,2020.
SUN Y Y.Thermo- and photoperiod- characterization,cytological characterization and genetic regulatory betwork of the ecotype male sterile line 373S in Brassica napus L.[D].Yangling Shaanxi:Northwest Agriculture & Forestry University,2020.
[27] SUN Y Y,ZHANG D S,WANG Z Z,et al. Cytological observation of anther structure and genetic investigation of a thermo-sensitive genic male sterile line 373S in Brassica napus L[J].BMC Plant Biology,2020,20(8):1-14.
[28] LI J L,WANG S,YU J,et al. A modified CTAB protocol for plant DNA extraction[J].Chinese Bulletin of Botany,2013,48(1):72-78.
[29] ZHAO H X,LI Z J,HU S W,et al. Identification of cytoplasm types in rapeseed (Brassica napus L.) accessions by a multiplex PCR assay[J].Theoretical and Applied Genetics,2010,121(4):643-650.
[30] 徐 亮,彭小松,贺浩华,等.甘蓝型油菜生态型细胞质雄性不育系育性的初步研究[J].西北农业学报,2006, 15(1):20-23.
XU L,PENG X S,HE H H,et al. Preliminary study on the fertility of the ecotypical CMS for Brassica napus L.[J].Acta Agriculturae Boreali-occidentalis Sinica,2006,15(1):20-23.
[31] 徐献锋,胡玉梅,于澄宇,等.甘蓝型油菜反型温敏核不育Huiyou50S的生理特征及遗传分析[J].华北农学报,2014,29(3):147-152.
XU X F,HU Y M,YU CH Y,et al. Physiological characterization and genetic analysis of reverse thermo-sensitive genic male-sterile line Huiyou50S in Brassica napus[J].Acta Agriculturae Boreali-Sinica,2014,29(3):147-152.
[32] 董军刚.甘蓝型油菜温敏细胞质雄性不育系417S选育与鉴定[D].陕西杨凌:西北农林科技大学,2010.
DONG J G.Development and indentification of thermos-sensitive CMS 417S in Brassica napus L.[D].Yangling Shaanxi:Northwest A&F University,2010.
[33] 陶芬芳,杨学乐,彭 烨,等.甘藍型油菜温敏核不育系100S的遗传特性分析[J].湖南农业大学学报(自然科学版),2016,42(2):125-128.
TAO F F,YANG X L,PENG Y,et al. Analysis of the inherited character of a thermo-sensitive genic male sterile line 100S in Brassica napus[J].Journal of Hunan Agricultural University (Natural Sciences),2016,42(2):125-128.
[34] 李 莓,陈卫江,席代汶.甘蓝型油菜温敏核不育系湘油91S的研究与应用前景[J].作物研究,1997(4):23-26.
LI M,CHEN W J,XI D W. Research and application prospect of thermosensitive male sterile line Xiangyou 91S in Brassica napus L[J].Crop Research,1997(4):23-26.
[35] 王 华,汤晓华,赵继献.甘蓝型油菜胞核雄性不育材料H90S的遗传研究[J].中国油料作物学报,2001,23(4):11-15.
WANG H,TANG X H,ZHAO J X. Genetic study on ecotype genetic male sterile of H90S in Brassica napus L[J].Chinese Journal of Oil Crops,2001,23(4):11-15.
[36] 李石開,苏振喜,吴学英.油菜‘K121S温敏核不育的遗传初探[J].西南农业学报,2004,17(增刊):49-54.
LI SH K,SU ZH X,WU X Y.Preliminary study on inheritance of Brassica juncea thermosensitive genicmale sterile fine ‘K121S[J].Southwest China Journal of Agricultural Science,2004,17(Suppl):49-54.
[37] 杨光圣,傅廷栋,杨小牛,等.甘蓝型油菜生态雄性不育两用系的研究Ⅰ.雄性不育两用系的遗传[J].作物学报,1995,21(2):129-135.
YANG G SH,FU T D,YANG X N,et al.Studies on ecotypical male sterile line of Brassica napus L. I. Inheritance of the ecotypical male sterile line[J].Acta Agronomica Sinica,1995,21(2):129-135.
[38] 袁 美,杨光圣,傅廷栋,等.甘蓝型油菜生态型细胞质雄性不育两用系的研究Ⅲ.8-8112AB的温度敏感性及其遗传[J].作物学报,2003,29(3):330-335.
YUAN M,YANG G SH,FU T D,et al.Studies on the ecotypical male sterile-fertile line of Brassica napus L. III. Sensitivity to temperature of 8-8112AB and its inheritance[J].Acta Agronomica Sinica,2003,29(3):330-335.
[39] 董军刚,董振生,刘绚霞,等.甘蓝型油菜生态雄性不育系533S花药发育的细胞学研究[J].西北农林科技大学学报(自然科学版),2004,32(7):61-66.
DONG J G,DONG ZH SH,LIU X X,et al.Cytological studies on anther development of ecological male sterile line 533S in Brassica napus L.[J].Journal of Northwest A&F University(Natural Science Edition),2004,32(7):61-66.
[40] 李可琪,曾新华,袁 荣,等.甘蓝型油菜温敏细胞核雄性不育系TE5A花药发育的细胞学研究[J].中国农业科学,2016,49(12):2408-2417.
LI K Q,ZENG X H,YUAN R,et al.Cytological researches on the anther development of a thermo-sensitive genic male sterile line TE5A in Brassica napus[J].Scientia Agricultura Sinica,2016,49(12):2408-2417.
[41] 葛 娟,郭英芬,于澄宇,等.甘蓝型油菜光、温敏雄性不育系Huiyou50S花粉败育的细胞学观察[J].作物学报,2012,38(3):541-548.
Expression of Thermo-sensitive Male Sterility Gene in Normal Cytoplasmic Type Derived from TGMS Line 373S (Brassica napus)
Abstract This study investigated the relationship between the thermo-sensitive male sterile gene and specific cytoplasmic type in thermo-sensitive genic male sterile (TGMS) line 373S in Brassica napus L.,with the aim of advancing the understanding of its male sterility mechanism and utilization in two-line hybrid breeding.This study used the female parent line Shaan 2B with normal cytoplasmic type cam and the TGMS line 373S as the male parent,in a cross,resulting in the separation ratios of fertile and sterile plants at 3∶1 and 15∶1 in BC1 and F2 population,respectively,suggesting that the male sterility trait was controlled by two pairs of recessive genes in this population. The sterile plants in the BC1 and F2 populations exhibited thermo-sensitive sterility.Compared with the line 373S,the seed-setting rate of male sterile plants slightly decreased,and the male sterile period had no significant difference in the segregated populations. Additionally,there were no significant differences in the corolla diameter,length of four stronger stamens anther,and length of two weaker stamens anther,while a slightly increase was observed in the length of stalk,length of petal,width of petal,length of four stronger stamens and length of two weaker stamens in the segregated populations. Cytological observation revealed abnormal microspores at pollen mother cell stage in male sterile plants in the segregated populations in terms of anther development stage,chromosome morphology and callose deposition,with no differences compared with the line 373S. These findings suggest that the TGMS gene(s) in the line 373S is successfully introduced from pol to cam cytoplasm,and the TGMS trait of 373S is caused by the TGMS gene(s) in the nucleus,independent of the cytoplasmic background.
Key words Brassica napus;Thermo-sensitive sterility gene;Cytoplasmic types;Male sterile
