植物花青素合成的环境调控研究进展
2024-03-19王凤华吴正景张菊平
王凤华,郭 佳,吴正景,张菊平
(1.河南科技大学 园艺与植物保护学院,河南 洛阳 471000;2.南京国环有机产品认证中心有限公司,江苏 南京 210042)
花青素是一种重要的天然植物色素,自然条件下,大多以糖苷的形式存在,称为花青苷。花青素和花青苷广泛存在于植物的组织和器官中,使植物呈现五彩缤纷的颜色。花青素也是影响园艺产品外观品质的重要因素,一般花青素含量高的产品,市场价格也较高。花青素还具有抗肿瘤、抗氧化、抗生物和非生物胁迫等作用。植物以苯丙氨酸为前体,按照图1 的途径合成花青素[1]。苯丙氨酸裂解酶(Phenylalanine ammonialyase,PAL)、查尔酮合成酶(Chalcone synthase,CHS)、查耳酮异构酶(Chalcone isomerase,CHI)、黄烷酮-3- 羟化酶(Flavanone 3-hydroxylase,F3H)、二氢黄酮醇还原酶(Dihydroflavonol 4-reductase,DFR)、花青素合成酶(Anthocyanidin synthase,ANS)、类黄酮-葡萄糖基转移酶(UDP-glucose flavonoid 3-O-glucosyl transferase,UFGT)等是催化花青素合成的主要酶类。编码这些酶的基因称为结构基因,在花青素生物合成中起主导作用。结构基因的表达受调节基因即转录因子的影响,bHLH(basic helix-loop-helix)、MYB和WD40等转录因子参与了植物花青素的合成。结构基因与转录因子共同形成了植物花青素合成代谢的分子调控网络。
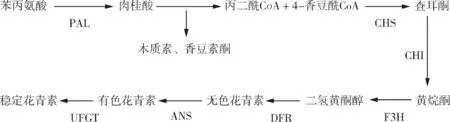
图1 植物花青素合成途径Fig. 1 Synthesis of plant anthocyanin
在植物花青素生物合成代谢中,环境因素也发挥着重要调控作用。例如,同株月季(Rosa chinen-sisJacq.)花经常随年份、月份呈现不同的颜色,这主要就是受环境的影响。环境因素通过影响结构基因和转录因子的表达而影响植物花青素的合成,并导致花青素含量、分布、稳定性等发生改变,从而颜色也发生相应改变[2]。
1 光照对花青素代谢的调控
1.1 光照对花青素结构基因表达的影响
花青素积累受光照强度的影响,随光照强度增加而增加[3]。调控植物花青素合成的酶类大都是光调节酶,光照可以提高这些酶的活性,促进花青素合成[4]。强光上调花青素合成相关基因CHS、CHI、DFR等的表达[5]。光质是影响植物花青素合成的另一重要因子,紫外光能诱导芜青(Brassica rapaL.)花青素合成[6]。不同部位对光质的反应存在差异,芜青中蓝光主要在下胚轴上部,紫外光UV-B 在中部和上部,UV-A 在下部到上部[7]。在一定范围内,光照时间越长,花青素积累越多[8]。叶片是光信号的感受器官,将离体矮牵牛(Petunia×hybridahort. ex Vilm.)置于黑暗中生长,最终花冠长度、花青素含量、CHS基因的表达被显著抑制。然而,将其花冠嫁接到其他植株上,遮住花冠,再进行照光,结果对花冠生长和CHS基因表达的影响甚微[9]。将月季花冠遮光,花瓣能正常显色,但将叶片遮光,显色受到抑制[10]。目前研究已经分离鉴定了一大批光受体,如光敏色素(Phytochrome,PHY):PHYA、PHYB、PHYC、PHYD、PHYE(吸收红光和远红光);隐花色素(Cryptochrome,CRY):CRY1、CRY2、CRY3(吸收蓝光和UV-A);向光素(Phototropic pigment,PHOT):PHOT1、PHOT2 及UVR8(UV-B 敏感光受体)等[11]。
1.2 光照对花青素合成转录因子的影响
光照对花青素的调控主要是影响其转录因子。光诱导转录因子MYB10、WD40和HY5的表达,会激活花青素生物合成途径中关键基因,促进花青素的合成和积累。HY5是诱导花青素生物合成的关键调控因子,MYB10与HY5相互作用,通过与G-box 基序结合,正向调节植物花青素生物合成[12]。光照增强果皮颜色,上调PyMYB10和PybHLH基因表达。MYB-bHLH-WD40转录复合体在光照下形成,参与调控果皮花青素积累。PyMYB10、PybHLH和Py-WD40转录因子组成的三元复合物调控云南红梨[Pyrus pyrifolia(Burm. F.)Nakai.]花青素的合成和积累[13]。光照诱导梨果实和愈伤组织中转录因子PpbHLH64的表达和花青素的积累[14]。
照光后,植物光受体将光信号传递给转录因子,转录因子接受光信号后与结构基因中的顺式作用元件相互作用调控花青素合成[15]。拟南芥(Arabidopsis thalianaL. Heynh.)CHS、FLS、F3H基因的启动子中均含有MYB识别元件[16]。HY5转录因子也是COP1 的靶基因,光照条件下,COP1 从细胞核中转移出来,与HY5结合,启动相关结构基因表达,促进花青素合成;黑暗条件下,COP1 在细胞核中与SPA 形成复合物COP1/SPA,使HY5泛素化,HY5降解,最终无法启动结构基因表达,花青素合成受抑[11]。拟南芥中,HY5通过影响转录因子MYB75/PAP1的活性而影响花青素合成[17]。MdCOP1与MdMYB1互作调控苹果花青素合成[18]。Ni 等[19]将突变体“红早酥”梨暴露在蓝光下,显著诱导花青素积累,同时Pp4ERF24和Pp12ERF96基因表达上调,说明光照可能引起乙烯表达量变化从而导致花青素积累。并不是所有植物的花青素合成都受光调控,例如葡萄(Vitis viniferaL.)的某些品种,光照对其花青素合成并没有显著影响[20]。
2 温度对花青素合成的调控
温度也是影响花青素积累的主要因素。低温促进花青素积累,高温抑制花青素合成。牡丹(Paeonia × suffruticosaAndr.)花青素合成相关基因受低温促进[21]。低温下,柑橘(Citrus reticulataBlanco.)PAL、CHS、DFR等基因的表达上调,果肉花青素含量增加[22]。低温更多是通过影响转录因子的表达而影响花青素的合成。低温下苹果(Malus pumilaMill.)果皮中的转录因子MdMYBA表达量明显增加,花青素含量增加[23];MdMYB73介导了植物的耐寒性,过量表达MdMYB23的转基因苹果愈伤组织和拟南芥花青素含量增加,耐寒性增强[24]。相反,高温则抑制植物花青素合成。拟南芥转录因子TT8、TTG1及EGL3等的表达受高温抑制,花青素积累减少[25]。参与花青素生物合成的AmCHS1、Am-CHI、AmF3H、AmDFR、AmANS和AmUFGT转录本水平在高温下明显降低[26]。高温诱导了HY5降解,抑制了MYBL2的表达,花青素减少,在cop1 和hy5突变植株中,花青素的生物合成和积累不受温度变化的影响[27]。温度还通过影响相关酶的稳定性而影响花青素积累,紫菀(Aster tataricusvL. f.)PAL、CHI 酶的活性受温度影响,温度高活性低。29℃/21℃(昼/夜)条件下,花瓣中的花青素含量只有17℃/ 9℃(昼/夜)时的一半。镁处理紫菀植物,在高温下,花青素浓度提高了80%,可以部分防止颜色褪色[28]。温度还影响花青素的稳定性,当环境温度大于30℃时,花青素不稳定,菊花花瓣较难着色或褪色[29]。
3 糖对花青素合成的调控
糖是构成花青素碳骨架的基础,也是一种信号分子,研究证明糖参与了植物花青素合成。萝卜(Raphanus sativusL.)可溶性糖含量与根肉色呈显著相关[30]。紫薯(Solanum tuberosumL.)花青素合成也受糖调控[31]。拟南芥CHS、CHI、DFR、F3H、PAL、DFR、UFGT和MYB75等基因的表达受蔗糖调控。蔗糖对拟南芥花青素合成相关基因的调控力度存在差异,一般来说对结构基因的影响最大,其次是正调控因子GL3、TT8和PAP1等,最后是负调控因子,如MYBL2[32]。蔗糖处理增加了金鱼草(Antirrhinum majusL.)切花花青素的积累,提高了花瓣在高温下的糖含量,AmCHS1、AmF3H、AmDFR、AmANS和AmUFGT转录水平也提高[26]。糖的种类不同其调控效果也不同。蔗糖增加花青素积累,果糖和葡萄糖效果甚微[33]。糖对花青素合成代谢的影响可能与糖的磷酸化作用相关,可以被磷酸化的糖能显著促进花青素积累,不能被磷酸化的则不能诱导花青素积累[34]。研究发现,在牵牛花和拟南芥的CHS基因5’端有个“蔗糖盒”[35],这似乎解释了蔗糖诱导花青素合成的原因。
4 激素对花青素合成的调控
激素主要通过2 种方式调控植物花青素合成,即正调控和负调控。
4.1 激素对花青素合成的正调控
生长素(Indole-3-acetic acid,IAA)、细胞分裂素(Cytokinin,CTK)、脱落酸(Abscisic acid,ABA)、茉莉酸(Jasmonate,JA)等能激活花青素合成。用IAA 处理拟南芥幼苗后,CHS、CHI、F3H、TTG1、PAP1、MYB12等基因表达上调[36]。CTK 受体AHK2、AHK3和AHK4(组蛋白激酶)以及ARR1、ARR10和ARR12(B 型响应调节因子)等参与了植物花青素合成,其中AHK2、AHK3和AHK4是正调控因子,CTK 显著抑制拟南芥突变体ahk2/3 和ahk3/4的花青素合成[37],这也证明CTK 正调控拟南芥花青素合成。拟南芥中有7 种B 型响应调节因子ARRs参与了CTK 信号的传递[38]。CTK 对植物花青素合成的调控可能还需要光的参与[11],其调控作用通过光电子传递信号途径进行[39]。ABA 诱导葡萄花青素合成[40],但是ABA对花青素合成的调控可能需要糖,因为单独采用ABA 处理拟南芥植株,只有PAP2和ATT的表达受影响,对其它相关基因的影响不大;但采用糖和ABA同时处理,则促进了大部分相关基因的表达[41]。ABA 的调控作用可能通过转录因子介导,在玉米中存在一个MYB转录因子,其活性受ABA 调控[42]。JA 也对植物花青素合成起正调控作用。茉莉酸甲酯(Methyl jasmonate,MeJA)通过上调相关基因的转录促进花青素的积累。MeJA 处理萝卜可以显著增加萝卜花青素含量[3]。Wang 等[43]发现转录因子MdMYB24L与JA 信号因子(Md-JAZ8,MdJAZ11和MdMYC2)存在相互作用,Md-MYC2 蛋白也是MdJAZ8和MdJAZ11的靶向蛋白,它们在MeJA 处理下迅速降解。MdMYB24L过表达导致转基因苹果愈伤组织中花青素含量高于对照,MdUFGT和MdDFR的表达上调。与ABA 类似,JA发挥作用也需要糖。因为仅用JA处理拟南芥,花青素合成不受影响;但是JA 和糖同时存在时,却显著促进了CHI、PAP1、PAP2基因的表达[11]。COI1 介导了JA 调控的植物花青素合成。JAZ 是茉莉酸信号途径的一类调控蛋白,JAZ 通过调控转录因子bHLHs和R2R3-MYBs而影响植物花青素合成[44]。
4.2 激素对花青素合成的负调控
赤霉素(Gibberellic acid,GA)和乙烯(Ethylene receptor,ETR)对花青素合成起负调控作用。外源GA3 处理显著降低萝卜花青素含量[45]。GA3 显著降低了结构基因DFR和转录因子PAP1和PAP2的表达水平[41]。缺乏赤霉素应答的拟南芥突变植株ga1,GA 前体无法生成,内源赤霉素含量较低,抑制作用降低,进而触发了参与花青素合成途径基因PAP1等表达上调,植株花青素含量增加[41]。ETR也是植物花青素合成的负调控因子,过量表达突变基因ETR1H69A(ETR 受体基因)的转基因烟草(Nicotiana tabacumL.)中,花青素含量显著增加[42]。植物体中存在乙烯受体,这些受体也影响花青素合成。已知的乙烯受体有ETR1、ETR2、ERS1、ERS2、EIN4等,其中EIN2、EIN3/EILs起正调控作用,ETRs和ERSs 起负调控作用[11]。在拟南芥乙烯突变体etr1-1、ein2-1、ein3和eil1中,花青素含量显著增加[13,39]。关于ETR 的作用机制,Kim 等[40]认为ETR通过下调bHLHs和MYB以及上调MYBL2的表达而抑制玉米(Zea maysL.)花青素合成。ETR抑制植物花青素合成可能与ETR三重反应有关[32]。
5 其他环境因子
盐、干旱、低氮胁迫等也对植物花青素合成存在显著影响。NaCl 胁迫下萝卜花青素含量显著增加[3]。转入花青素合成相关基因可以显著提高植物的抗盐能力,在转Del基因的烟草中,NtCHS、Nt-CHI、NtF3H、NtDFR和NtANS表达上调,花青素含量显著增加,抗盐能力显著提高[46]。过量表达AtDFR的甘蓝型油菜(Brassica napusL.)DFR基因的转录水平显著升高,花青素含量提高,抗盐力提高[47];而缺失花青素合成相关基因抗盐能力则降低[48]。干旱胁迫也影响植物花青素合成。干旱胁迫下,花青素合成相关的结构基因和调节基因表达上调,花青素含量上升[49-50]。转Del基因和R2R3-MYB基因的植物,花青素含量提高,抗旱能力提高[46,49]。缺氮条件下,植物花青素含量升高,这也许是植物适应逆境胁迫的一种方式,转基因苹果花青素积累量增多,抗低氮胁迫的能力提高[51-52]。在低氮胁迫下,拟南芥tt3突变体(DFR缺失),DFR合成受阻,抗低氮胁迫能力减弱[52]。海拔、pH值等也与植物花青素合成有关[53],这些因素主要通过影响光、温度、水等的再分配影响花青素的合成,植物花青素的合成可能还与Ca2+信号途径有关[54]。
5 问题与展望
环境因子主要通过调控结构基因和转录因子的表达影响植物花青素合成,这也是植物为了生存自我调节适应逆境的一种应激反应。尽管已经从拟南芥、苹果和矮牵牛等植物中获得了参与花青素合成代谢的转录因子,但是环境因子对这些转录因子的作用机制以及转录因子与结构基因的相互作用机制仍不清楚。实际生产中,环境因子不会单独存在,常常是多因素协同作用。例如,蓝光诱导拟南芥CHS基因的表达受蔗糖的影响;低温诱导拟南芥花青素合成需要光的参与;蔗糖诱导花青素合成,但会被赤霉素抑制;光呼吸和温度共同调控桃花青素代谢[55],那么各因素之间是如何协同作用共同调控植物花青素代谢的呢?相关机制有待阐明。另外,植物花青素的抗氧化功能,不仅表现在非生物胁迫中,也表现在生物胁迫中。
今后的研究可以着力解决以下问题:环境因子与转录因子的作用机制,转录因子与结构基因的相互作用机制,多因子如何相互作用,各因子调控的共同点和不同点等。由于花青素的抗氧化功能,花青苷已经成为一个重要的植物育种性状,如何将理论研究用于植物育种中也是广大育种学家面临的重要问题。相信随着分子生物学相关技术的发展,这些问题会逐步得到阐明。
