大黄根腐病与土壤营养及微生物群落组成的相关性研究
2024-03-18林亚蔡瑜胡凡王佳瑜江虹霖周双双张庆伟
林亚,蔡瑜,胡凡,王佳瑜,江虹霖,周双双,张庆伟
1.西南大学 生命科学学院/三峡库区生态环境教育部重点实验室,重庆 400715;2.重庆市南川区道地中药材研究院,重庆 南川 408407;3.重庆灏天生态农业科技有限公司,重庆 南川 408407
大黄是一种重要的中药材,来源于掌叶大黄(RheumpalmatumL.)、唐古特大黄(RheumtanguticumMaxim ex Balf)和药用大黄(RheumofficinaleBaill.)的干燥根及根茎.大黄在西南地区的种植范围包括贵州、四川、重庆等地,其中重庆石柱、奉节、南川等地大黄种植规模较大,具有重要的产业价值.大黄在种植过程中会发生多种病害,如轮纹病、炭疽病、霜霉病、根腐病,以及黑粉病等[1-2],其中危害较大的是根腐病.该病主要危害根部,一经发生,危害严重,治理困难[3].染病后根部先腐烂,然后叶片渐渐变黄,直至整株枯死.目前已有研究表明镰刀菌(Fusarium)、丝核菌(Rhizoctonia)、小核菌(Sclerotium)和链格孢(Alternaria)等属微生物会引起植物根腐病,其中半知菌亚门镰刀属菌种所占比例最大.它是最早发现的根腐病病原菌,具有高度的寄生性,引起根腐病发生的概率较高,并且致病力也较强[4-5].根腐病病原菌存在于土壤中,土壤理化性质与其生长及侵染植物有着直接的关系[6],同时土壤微生物种类繁多,数量庞大,不同类型的微生物之间相互促进或相互制约,也是根腐病病原菌繁殖与侵染植物的重要条件.针对影响根腐病病原菌生长繁殖的相关环境条件,本研究通过分析不同发病率大黄生长环境的理化性质,并结合微生物群落分析,探究大黄根腐病发生与土壤理化性质之间的关系,以期为大黄的种植和根腐病的治理提供理论基础.
1 材料与方法
1.1 研究区域
前期已了解到大黄根腐病在重庆石柱地区发生较为普遍,因此选择有近350 hm2种植规模的石柱土家族自治县冷水镇(108.27°E,29.92°N)多个大黄种植基地作为调查点.该镇属亚热带季风性湿润气候,海拔高度约为1 550 m,年降雨量约1 400 mm,年平均气温10.4 ℃.当地具有悠久的大黄种植传统,以农业合作社为单位进行大黄的种植和加工,种植技术和管理方式较为一致.
1.2 土壤样品采集
2021年9月15日,根据当地种植户提供的初步信息(不同地块发病率情况),选取30块样地,包括平地(坡度小于 15°)和坡地(在15°~30°之间).结合大黄根腐病的发病特征(症状主要发生在根的中上部和根茎部,发病初期根茎形成淡褐色和黑褐色不规则形病斑,并有局部腐烂,发病后期根茎组织全部腐烂,产生黑色胶状物,并伴随有叶片焦枯症状[1]),进行根腐病发病率(样地发病株数/样地总株数)的统计.对于症状不明确的植株,采集根茎组织带回实验室进行病原菌鉴定后确认.病原菌鉴定方法主要为使用显微镜观察发病部位是否有大量的镰刀菌存在,镰刀菌形态鉴定参照相关文献[7-8]进行.初步计算样地根腐病的发病率后,采用五点取样法进行土壤样品的采集[9],即以样地对角线的中点为中心采样点,再从对角线上选择4个等距中心点作为采样点.在每个采样点先用环刀取土,然后再从土壤表层下5~20 cm取样.采集获得的土样通过除去杂物并充分均匀后分为两部分:一部分用于理化指标分析,另一部分进行冷冻保存以用于后续的DNA提取以及16S和ITS测序.
1.3 土壤理化性质分析
理化性质指标包括土壤pH值,有机碳,碱解N,速效K,有效K,缓效K,全N,全K,全P,Cu,Zn,Fe,Mn等元素的质量分数,具体测定方法参考《土壤农业化学分析方法》[10]进行.取用于测定理化性质的土样,在室内自然干燥后进行研磨,之后分两步筛土,第一步将这些土样通过2 mm土壤筛,过筛后的土样一部分用于土壤pH值,碱解N,速效K,有效K,缓效K质量分数的测定,另一部分将过2 mm土壤筛的土样再通过0.25 mm土壤筛用于有机碳,全N,全K,全P,Cu,Zn,Fe,Mn质量分数的测定.各指标具体测定方法如表1.

表1 土壤理化指标的具体测定方法
1.4 土壤生物学性质分析
由北京诺禾致源科技股份有限公司完成对大黄根际土壤DNA的提取、扩增和测序.
1.5 数据统计方法
利用 Excel 2019对所得数据进行初步整理,采用SPSS 22.0进行数据分析,具体采用单因素(One-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05),并利用皮尔逊(Pearson)方法对大黄根腐病发病率与土壤理化性质、微生物多样性之间的相关性进行研究.
2 结果与分析
2.1 样地大黄根腐病发病率
通过实地调查统计结合采集样品进行病原菌的鉴定,最终计算出每个样地根腐病的发病率,并根据发病率的范围将样地分为4种类型,即轻微、轻度、中度和严重(表2).实际调查中发现大黄根腐病发病率普遍较高,这与种植环境有直接的关系.一般坡地发病率相对较低,低洼地块发病率偏高,而重茬地的发病率最高,部分地块接近100%.由于是根据发病率进行样地选择,调查样地数量有限,本研究并未分析不同地块的地形和耕作方式与根腐病发生之间的关系.

表2 采样地概况及发病率
2.2 不同发病程度地块土壤的理化性质
2.2.1 土壤pH值与有机碳质量分数差异
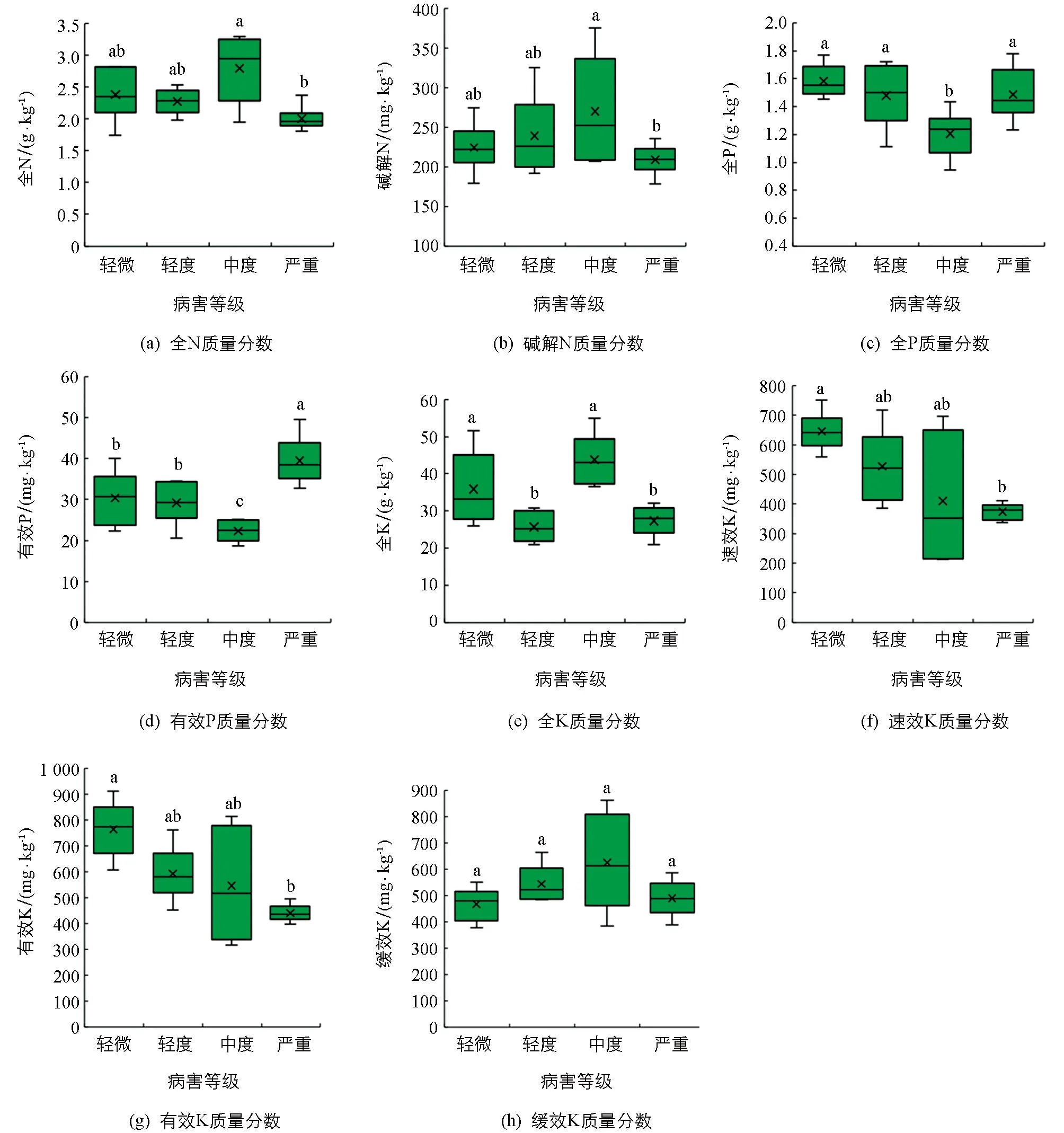
通过分析发现所取样地的大黄种植地块土壤pH值大都接近中性,不同发病程度(病害等级)地块的土壤pH值之间差异无统计学意义(图1a).在土壤有机碳质量分数方面,不同发病程度地块之间存在差异,中度发病程度的地块土壤有机碳质量分数高于其他发病程度的地块,且差异达到统计学意义(图1b).

小写字母不同表示p<0.05,差异有统计学意义.
2.2.2 土壤大量元素质量分数差异
通过分析发现不同发病程度地块土壤样品中N,P和K相关指标存在差异(图2).具体表现为:1) 发病程度为中度的地块土壤全N和碱解N质量分数高于其他发病程度地块土壤,且显著高于严重发病地块土壤; 2) 除中度外,不同发病程度地块土壤全P质量分数之间差异无统计学意义,但有效P质量分数差异较大,中度发病程度地块土壤有效P质量分数低于轻微发病和轻度发病地块土壤,三者都低于严重发病地块土壤,且差异有统计学意义; 3) 在土壤全K质量分数上,轻度和严重发病地块土壤低于轻微和中度发病地块土壤,差异有统计学意义,而在速效K和有效K质量分数方面,随着发病程度的加深,其质量分数呈现下降趋势,且轻微发病地块土壤高于严重发病地块土壤,差异有统计学意义.
小写字母不同表示p<0.05,差异有统计学意义.
2.2.3 土壤微量元素质量分数差异
为探索土壤微量元素与大黄根腐病发生的关系,本研究分析了常见微量元素Fe,Mn,Cu和Zn质量分数,结果表明不同发病程度地块之间土壤微量元素质量分数存在一定的差异(图3).随着病害程度的加深,土壤Fe质量分数呈现下降趋势,但不同病害程度地块之间差异无统计学意义.与之相反,土壤Mn质量分数呈现上升趋势,而且严重发病地块高于其余发病地块且差异有统计学意义.在土壤Cu质量分数方面,不同发病程度地块之间差异无统计学意义.此外,不同发病程度地块土壤Zn质量分数之间存在差异,中度和严重发病地块低于轻微和轻度发病地块,其中轻度发病地块高于中度发病地块,中度发病地块高于严重发病地块,差异有统计学意义.

小写字母不同表示p<0.05,差异有统计学意义.
2.3 发病程度与土壤营养元素质量分数之间的相关性
为从众多土壤理化指标中筛选与大黄根腐病发生率关系更为密切的指标,本研究进行了相关性分析,结果如表3.由于严重发病地块多数为连作(重茬)地块,具有特殊性,本研究排除连作地块后对获得的数据进行相关分析,结果表明发病率与土壤速效K,有效K,Fe,Mn和Zn质量分数有显著的相关性,其中土壤Mn质量分数与发病率呈正相关,其余与发病率呈负相关.从相关系数和显著性上来看,土壤速效K,有效K及Zn质量分数与发病率之间相关系数较高,差异有统计学意义,说明三者与根腐病的发生更为相关.
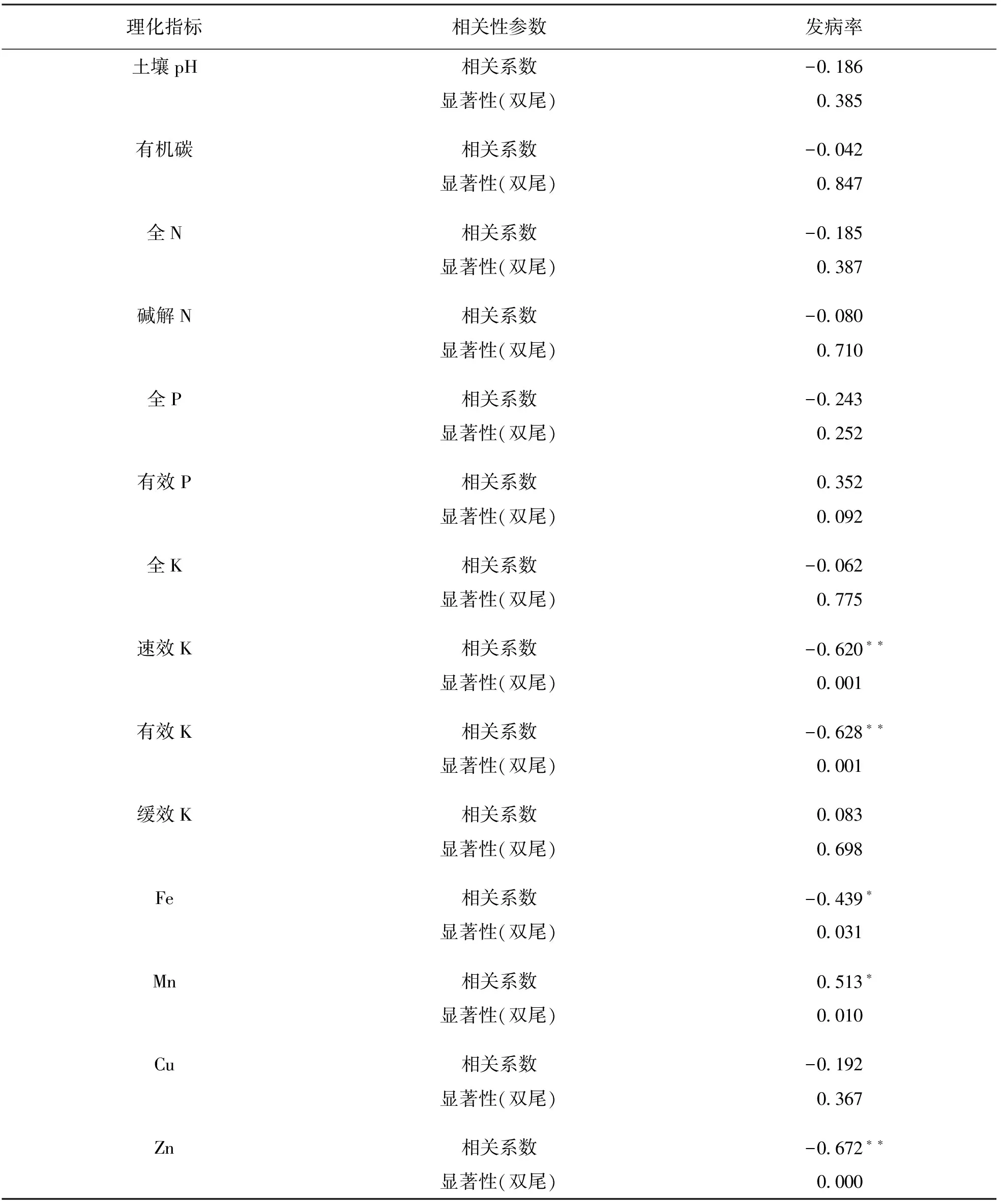
表3 大黄根腐病发病率与土壤理化指标之间的相关性分析结果
2.4 不同病害等级地块土壤微生物群落差异
2.4.1 不同病害等级地块土壤细菌群落组成
16S和ITS扩增子测序结果表明不同发病程度地块土壤细菌和真菌的群落组成具有一定的差异.图4显示,尽管从不同发病程度地块土壤样品中获得的细菌和真菌OTU数量之间没有明显差异,但在土壤微生群落组成上,不同发病程度地块土壤中细菌和真菌的群落组成差别较大(图5).对细菌而言,在门的分类水平上,不同发病程度地块土壤之间存在差异的细菌群落主要分布在厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)和浮霉菌门(Planctomycetes); 而在目的分类水平上,不同发病程度地块土壤之间存在差异的细菌群落主要分布在黄单胞菌目(Xanthomonadales),γ-变形菌纲(Gammaproteobacteria)下的某些种类和噬几丁质菌目(Chitinophagales)等.对于真菌而言,在门的分类水平上,出现差异的真菌主要分布在子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和被孢霉门(Mortierellomycota); 在目的分类水平上,不同发病程度地块土壤之间存在差异的真菌群落主要涉及粪壳菌目(Sordariales),囊丝担子菌目(Cystofilobasidiales)、尾囊黑粉菌目(Urocystidales)、格孢腔菌目(Pleosporales)、被孢霉目(Mortierellales)和鸡油菌目(Cantharellales).

图4 不同病害等级地块土壤细菌和真菌OTU数量

uf表示未能具体鉴定到目.
2.4.2 不同病害等级地块土壤微生物群落丰富度和多样性指数
根据16S和ITS扩增子测序结果,对不同发病程度地块土壤样品中细菌和真菌群落进行丰富度和多样性分析,结果表明不同病害等级的大黄根周围土壤细菌群落的丰富度指数(Chao1,ACE)、均匀度指数(Simpson)以及多样性指数(Shannon)差异均无统计学意义(表4).从土壤细菌群落丰富度来看,病害严重土样(中度、重度)的细菌群落最丰富; 从土壤细菌群落均匀度来看,随着病害严重性的增加,细菌群落的均匀度也随之增加; 从土壤细菌群落多样性来看,随着病害严重性的增加,细菌群落的多样性也随之增加(表4).对于不同发病地块土壤的真菌群落而言,除轻微发病地块土壤外,其余土壤样品中,随着病害程度的增加,真菌群落的丰富度、均匀度以及多样性都表现出一定的增高趋势,但不同发病程度地块多个指标之间差异无统计学意义(表5).

表4 不同病害等级地块土壤细菌群落丰富度、均匀度和多样性指数

表5 不同病害等级地块土壤真菌群落丰富度、均匀度和多样性指数
为探究细菌和真菌群落丰富度及多样性指数是否与病害的发生有关,本研究进行了相关性分析(表6).结果发现:1) 对于细菌群落,发病率与Chao1,ACE以及Simpson之间没有明显的相关性,而与Shannon呈现出显著正相关; 2) 对于真菌群落,发病率与Chao1,ACE,Simpson以及Shannon之间都没有明显的相关性.这些结果表明大黄根腐病的发生与细菌群落有一定的相关性,且随着大黄根腐病发病率的增加,细菌群落的多样性也随之增加.

表6 土壤细菌与真菌群落丰富度和多样性指数与发病率之间的相关性
2.4.3 不同病害等级地块土壤微生物优势物种差异
对土壤微生物优势物种(相对丰度排名前20的属)进行分析,发现不同病害等级地块土壤细菌和真菌的优势物种差异较大(图6).在细菌群落方面,发病程度较轻(轻微、轻度)的地块土壤中罗河杆菌属(Rhodanobacter)、伯克霍尔德氏菌科(uf_Burkholderiaceae)某些属和代尔夫特菌属(Delftia)丰度较高; 而发病程度较高(中度、严重)的地块土壤中γ-变形杆菌纲(uf_Gammaproteobacteria)某些属、酸杆菌门(uf_Acidobacteria)某些属、亚硝化单胞菌科(uf_Nitrosomonadaceae)某些属和溶杆菌属(Lysobacter)细菌丰度较高.对于土壤中真菌优势种群,尽管发病程度不同,但所有样品都检测到了潜在能引起根茎腐烂的病原菌属,即黑粉病菌属(Thecaphora)、亚隔孢壳属(Didymella)和镰刀菌属(Fusarium),而且它们在发病程度较高的地块土壤中丰度相对较高.此外,通过比较可以发现发病程度较轻的地块土壤中腐质霉属(Humicola)、被孢霉属(Mortierella)等丰度较高,而发病程度较高的地块土壤中枝顶孢属(Acremonium)、四枝孢属(Tetracladium)和枝鼻菌属(Cladorrhinum)等属真菌丰度变化较大.

图中数字表示细菌或真菌的相对丰度(%); uf表示未能具体鉴定到属.
3 讨论
根腐病是一种土传病害,由生活在土壤中的病原菌引起,在合适的条件下,病原菌侵入根系,造成根腐病的发生[11].病原菌的生长繁殖与土壤性质和土壤微生物组成关系密切,本研究也发现根腐病发病率较高的地块在多种营养元素的质量分数上与低发病率地块有所不同,发病率较高的土壤样品pH值相对较低.众多研究表明土传病害的发生与土壤酸碱度有一定关系,如高鑫媛等[12]发现患根腐病北苍术植株根际土壤的pH值显著低于健康北苍术植株根际土壤.另外,本研究还发现中度发病地块土壤有机碳质量分数显著高于轻微发病和轻度发病地块以及严重发病地块,尽管这个结果不能说明土壤有机碳质量分数影响了大黄根腐病的发病率,但类似的研究表明土壤有机碳质量分数与多种根茎类植物根腐病的发生有着紧密的联系,如唐涛等[13]通过对比健康和染病黄连根际土壤养分时发现染病植株根际土壤有机碳质量分数低于健康植株.
K元素对于植物生长发育至关重要,有助于增强植物对于多种病原菌的抗性[14],其中就包括引起根腐病的镰刀菌[15].本研究发现,发病率较高的土壤样品在速效K、有效K等指标上显著低于发病率较低的土壤样品,这暗示土壤K元素被植物吸收后在应对根腐病相关病原菌的过程中发挥了作用.该结果与赵林艳等[16]针对白及根腐病的相关研究结果相一致,即与健康植株相比,白及根腐病患病植株根际土壤有效K质量分数较低.
除此之外,本研究还发现根腐病发病率较高的地块土壤Mn质量分数显著高于发病率较低的地块土壤,与之相反,发病率较高的地块土壤Zn质量分数显著低于发病率较低的地块土壤.虽然Mn和Zn等微量元素对于植物生长发育具有重要的作用,但关于土壤微量元素质量分数与根腐病等植物病害发生之间的关系,目前还没有确切的证据.曾有人提出“Zn,Ni等元素在植物中的超量积累能够提高植物对于病害的抗性”的假说[17],而且还有一些证据支持这种假说[18].本研究中发病率较低地块土壤中Zn质量分数相对较高,但不等同于植物中的超量积累,所以无法用上述假说去解释这一现象.
将多种土壤营养指标进行综合分析,根腐病发病程度较高土样和根腐病发病程度较低土样之间在有机碳、有效K、速效K以及微量元素Mn和Zn质量分数上存在差异,而且这些差异的规律并不一致.这说明大黄根腐病的发生与多种土壤营养元素之间关系复杂,单一的土壤营养指标只能在一定程度上影响根腐病的发生.
土壤微生物在农业生产中占有举足轻重的地位,当土壤病害发生时,健株和病株根际土壤中的微生物多样性和种群之间的动态平衡往往存在较大的差异[19].本研究对不同病害程度地块土壤微生物的多样性和丰富度构成进行对比分析,结果表明,大黄根际土壤细菌和真菌群落的Chao1,ACE,Shannon和Simpson在不同病害等级上差异无统计学意义,表明不同病害等级的大黄植株根际土壤细菌和真菌群落的丰富度、多样性和均匀度差异不大.利用皮尔逊法对发病率与细菌群落多样性进行相关性分析,发现发病率与细菌群落多样性指数Shannon显著正相关,表明根际土壤中细菌群落的多样性与大黄根腐病的发生密切相关.这与闫欢等[20]发现黄芪根腐病的发病率与土壤细菌群落呈正相关的实验结果一致.关于根腐病的发生对于土壤微生物多样性的影响,目前众多研究仍没有形成明确的结论,例如健康枸杞和北苍术根际土壤的真菌群落丰富度、多样性及均匀度指数均高于患病株[12,21],而在黄连和黄精相关的研究中则发现患病株根际土壤细菌和真菌丰富度及多样性都要高于健康植株[13,22].这些都说明虽然普遍认为根腐病主要由镰刀菌属等真菌引起,但其对于土壤微生物多样性的影响仍然会因植物种类的不同而有所差别.
通过分析不同发病程度地块土壤样品的细菌群落结构,发现4种土壤样品的优势细菌群落在门水平上相对一致,优势门主要有变形菌门、拟杆菌门,酸杆菌门等,不同发病程度地块土壤之间存在差异的细菌群落主要分布在厚壁菌门、拟杆菌门和浮霉菌门; 而在目的分类水平上,不同发病程度地块土壤之间存在差异的细菌群落主要分布在黄单胞菌目,γ-变形菌纲下的某些种类和噬几丁质菌目等.现有研究表明以黄单胞菌为代表的黄单胞菌目细菌是一类能够侵染水稻、小麦、番茄以及十字花科等多种植物的病原细菌[23],其是否与大黄根腐病的发生有关,有待深入研究.在属的水平上,土壤细菌中相对丰度较高的前5个属是罗河杆菌属、伯克霍尔德氏菌科某些属、γ-变形杆菌纲下某个属、酸杆菌门某些属和代尔夫特菌属.染病和重病植株根际土壤中γ-变形杆菌纲下某些属、酸杆菌门某些属丰度更高,尤其是γ-变形杆菌纲下某个属在染病样品中普遍富集,说明γ-变形杆菌纲细菌的大量存在可能是导致大黄根腐病发生的原因之一.罗河杆菌属显著富集于轻微发病程度地块土壤样品中,在中度和严重病害的土壤样品中丰度较低,表明罗河杆菌属细菌可能有抑制根腐病相关病原菌发生的作用.Shi等[24]发现通过施用生物有机肥使三七根际罗河杆菌属和其他属细菌丰度提高,进而提高三七的产量和抗病性.另外,在Zhang等[25]的研究中也发现,施用某些生物试剂可以提高罗河杆菌属细菌的丰度,从而抑制导致辣椒根腐病的镰刀菌的丰度.在发病程度较轻的地块土壤中存在丰度较高的某些伯克霍尔德氏菌科的细菌,其可能也是大黄根腐病发病率较低的原因之一,因为相关证据表明在根际环境中伯克霍尔德氏菌是一类重要的植物促生菌,其群落结构变化能够影响植物的生长和发育[26].
对土壤样品的真菌群落组成进行分析可以发现,不同发病程度地块土壤中的优势真菌群落差异主要分布在粪壳菌目(Sordariales)、囊丝担子菌目(Cystofilobasidiales)、尾囊黑粉菌目(Urocystidales)、格孢腔菌目(Pleosporales)、被孢霉目(Mortierellales)和鸡油菌目(Cantharellales)中,而具体到属,可以发现黑粉病菌属、亚隔孢壳属、镰刀菌属和枝顶孢属等.这些与根腐病关系更为密切的真菌存在于不同发病程度的地块土壤中,而且随着根腐病发病程度增加,大多属丰度也有升高的趋势.这一现象与本研究进行取样的地区有关,该地区存在较长时间的大黄种植历史,根腐病相关的病原菌在土壤中广泛存在.此外,发病较轻的地块土壤中腐质霉属、被孢霉属丰度较高,而发病程度较高的地块土壤中四枝孢属、枝鼻菌属等属真菌丰度较高.相关研究表明腐质霉属、被孢霉属真菌是土壤碳及养分转化的关键微生物成员[27],其是否能够抑制根腐病相关病原菌的发展,目前尚未有清楚的答案.在关于人参的研究中发现,连作会导致土壤中有益真菌被孢霉属的丰度降低,而潜在的致病真菌四枝孢属的丰度上升[28].另外,如上所述多种病原菌都随着发病程度的加深而丰度增加,在一定程度上也暗示引起根腐病的病原菌不是一种或者一类微生物.
调查中发现多数发病严重的地块属于连作地块.连作土壤中往往存在极为严重的土传病害,且伴随着植物正常生理活动受抑、化感自毒物质积累、土壤酸化、养分失衡、微生物群落结构失衡等现象,最终造成植物生长发育不良[29].连作土壤的pH值变化明显,如连作后的西洋参土壤pH值显著降低,致使植株无法存活[30].土壤pH值下降影响土壤的电导率与氧化还原状态,同时影响土壤有机质的转化速率,进而影响土壤有机质质量分数[28].在不同连作土壤中N,P,K养分质量分数存在不同的变化趋势,例如Liu等[31]研究表明,土壤总N,P,K质量分数与有效态N,P,K质量分数均随连作时间的延长呈上升趋势,而部分有效态养分如有效P,速效K质量分数却随连作时间的延长呈现下降的趋势[32].另外,连作会导致原有根际土壤微生物群落动态平衡受到破坏.如连作3年的西洋参(PanaxquinquefoliusL.)土壤中细菌群落的多样性指数降低,而真菌群落的多样性指数则显著增加[32]; 但在白术的连作土壤中,真菌群落多样性表现出相反的变化趋势[33].在本研究中也发现连作后根腐病发病较高的地块土壤中细菌和真菌群落丰富度和多样性指数偏高.因此连作对于微生物群落多样性的影响机制还有待更深入的研究.
4 结论与展望
4.1 结论
通过对重庆石柱地区大黄根腐病的发生与土壤营养元素质量分数和微生物群落组成之间关系的研究,综合分析得出以下结论:
1) 土壤营养元素中,速效K、有效K质量分数与根腐病发病率之间存在极显著的相关性,应该是影响根腐病发生的重要因素; 土壤Mn,Zn,Fe元素质量分数与大黄根腐病的发生也有一定的相关性.
2) 通过对比不同病害等级地块土壤的细菌组成,发现在属水平上,差异细菌群落主要分布在罗河杆菌属、伯克霍尔德氏菌科下某些属等; 其中,较低发病率土壤样品中罗河杆菌属、伯克霍尔德氏菌科下某些属丰度较高,暗示其有助于抑制根腐病相关病原菌的发生.
3) 通过对于土壤真菌群落组成进行分析,发现多种致病病原菌(黑粉病菌属、亚隔孢壳属、镰刀菌属和枝顶孢属)在不同病害程度地块土壤中普遍存在,随着发病程度增加其丰度变化较大.此外,不同发病程度地块土壤中差异细菌群落主要分布在腐质霉属、被孢霉属、四枝孢属和枝鼻菌属等中,前二者有益于植物的生长发育,在发病较轻地块土壤中丰度较高,而后二者可能与根腐病的发生关系密切,在发病程度较高的地块土壤中丰度较高.
4) 不同病害程度地块土壤细菌和真菌在多样性和丰富度上存在一定的差异,但差异并无统计学意义.相关分析发现,大黄根腐病的发生与反映细菌群落多样性指数的Shannon显著相关,说明随着大黄根腐病发病率增加,细菌群落的多样性也随之增加.
4.2 展望
本研究初步分析了重庆石柱地区大黄根腐病与土壤理化性质和微生物群落的关系.未来研究可以从以下3方面深入:1) 通过精确实验验证土壤理化性质与根腐病的关系; 2) 利用宏基因测序等技术更详尽地分析根际土壤细菌群落结构; 3) 关注病原菌侵害植物和植物抗性的变化,为抗病品种选育提供依据.
