草地植被生产力模拟及其影响因素
2024-03-14赵泽钰黄娅兰秦荣竹张开平
赵泽钰,杨 阳,黄娅兰,汪 悦,秦荣竹,张开平,张 峰
(兰州大学生态学院, 甘肃 兰州 730000)
草地是全球重要的陆地生态系统类型之一,在世界上分布广泛,全球陆地总面积的20%左右都是草地,大约为32 亿hm2[1]。草地具有提供自然资源和生态屏障的重要作用,如防风固沙、水土保持以及生物多样性保护等功能。同时,草地也为发展草原畜牧业提供物质基础[2],在生态和经济方面都具有重要的意义。由于人类活动对生态系统的干扰和全球温室效应日益显著,草地生态系统对全球气候变化和碳氮循环的响应成为当前研究热点之一。
植被净初级生产力(net primary productivity, NPP)是绿色植物通过光合作用固定的碳总量和自养呼吸作用消耗的碳在单位面积、单位时间内的累积量[3],可以反映生态系统的生产状况,全球气温升高、降水格局改变、碳排放变化等因素都会引起草地生产力的波动。使用模型估算植被净初级生产力并且通过模型研究气候变化、管理方式、土壤条件等因素对草地生物量的影响,对于理解草地碳动态对全球气候变化的响应有重要意义。经过多年来国内外学者的努力和探索,植被生产力模型在草地生态系统的研究方面取得了很大进展,经历了从简单的统计模型,到结合遥感卫星数据的光能利用率模型,再到引入植被生长的物理化学过程的生态系统过程模型的发展过程。
因此,本研究就有关草地植被生产力的影响因素和估算模型作一综述,分析了不同研究条件下草地植被生产力对各影响因子的响应情况,详细介绍统计模型、光能利用率模型和生态系统过程模型在草地生态系统中的应用,探讨不同模型在研究中的优势以及局限性,以期为未来相关领域的深入研究提供一些参考。
1 影响草地生产力的因素
1.1 人类活动
人类活动对草地的影响主要有两个方面,一个是放牧的过程中牲畜的采食以及粪便分解等过程对植被的影响,另一个是农业耕地管理和居民地以及交通区域的人类活动范围扩张对草地的影响。
首先,大量放牧通常会导致草地上的植被覆盖度降低和凋落物积累减少,以及土壤中的水分蒸发加速和水分储存的改变,进而促进风对土壤的侵蚀作用,导致表层土壤进一步沙化,并且会加剧土壤有机碳和氮的损失[4]。其次,牲畜的采食会影响草地上的植物种类组成及物种多样性,而动物的排泄过程会影响土壤碳氮循环和土壤养分的有效性等[5]。另外,不同的放牧强度对土壤和植被的影响也有所不同。大量研究表明,过度放牧是干旱地区半干旱地区荒漠化的主要原因,在内蒙古奈曼地区的一项研究表明,放牧不仅直接改变草地生态系统的植被覆盖度和物种组成,还通过改变地表粗糙度间接影响放牧试验区的风态,例如放牧会引起植被的冠层结构变化,如植被覆盖度和冠层高度的降低,导致风可以直接作用于沙质草地表面,加剧沙漠化[6]。
放牧对草地生产力的影响大多是负面的,但也有一些研究表明放牧可能会提高草原的生产力,另外不同的放牧强度对植被和土壤的影响也随生长季节而变化。研究表明[6],轻度放牧的草地上没有发生荒漠化,中度放牧的草地上冬季比夏季更易出现荒漠化,这是由于在冬季植被处于休眠状态,降水较少,地区强风多发。此外,放牧对氮矿化速率的促进或抑制也会对草地生产力产生影响,总的来说,在草地生长早期阶段,放牧强度在中度以上是可以提高土壤氮的矿化速率的,但在生长季和非生长季阶段的中度放牧则会降低土壤氮的矿化速率,这不利于草地生产力积累[7]。但是,有研究表明,适度放牧由于提高了土壤氮的可利用性[8],有利于维持草地物种多样性和草地的生产力水平,可能还会产生补偿性生产。其次,放牧还可以通过减少竞争优势物种的丰度进而提高植物多样性[9],例如在乌拉圭草原的一项研究表明放牧会导致优势物种的转变(冷季的草丛减少而暖季的草丛有所增加),且放牧区的杂草数量有所增加[8],尤其是不同放牧牲畜的混牧更有利于草地植物多样性的维持。但过度放牧,会对草地生产力、牧草质量、凋落物累积量、地表抗旱性及生态系统恢复性等方面造成影响[10]。因此,对于健康草地而言,适度放牧优于禁牧,这既可以获得由畜产品生产而得的经济效益,又可以维持甚至提高草地生态系统的各项生态功能。
另外,人类活动如农业耕垦、放牧、采矿、居民地和交通区域的扩张等也会对草地产生影响,当草地距离人类居住地越近,其生产力受到人类活动影响越大,不仅会影响草地面积,还会造成草地生产力下降。
1.2 气候变化
1.2.1 温 度和降水
气候变化会对草地生态系统的影响非常大,草地的群落结构及组成、草地生物量和空间格局都会因此发生变化,因此研究气候变化和草地之间的相互作用及影响是目前全球变化研究的热点之一。温度和降水是与植物生长最密切相关的气象因素,因此关于温度和降水对植被生产力的影响的研究有很多,总的来说,由于研究区域的植被类型、地理位置和气象条件不同,温度和降水对于草地生产力的影响大小也不同[11]。
水分在植被的生长过程中发挥了很重要的作用,例如降水量的增加可以通过影响土壤含水量来促进植物光合作用速率,进而提高植被生产力。而温度通常可以决定草地空间分布,并且可以影响植被在进行光合作用时所需要的酶的活性。
来自高纬度和高海拔地区的植物群落对于气候变暖尤为敏感,有研究表明寒冷生物群落在气候变暖的影响下,草本植物和灌木会向更北的地区转移,植被的生产力有所提高[12],这可能是由于在高纬度的草原生态系统中,植物的生长主要受到氮的可用性限制,而温度升高导致高纬度地区的氮的可用性更高[11]。一项位于内蒙古半干旱草原生态系统的研究认为,温度升高对根生产力的影响大于对地上植被生产力的影响[13],虽然在气候变暖的条件下,提高了植被的生产力,但同时也导致生态系统中植物物种丰富度有所下降,引起了物种损失[14]。
也有研究认为,温度升高对草地生态系统中多年生物种的长期影响取决于温度对土壤湿度的影响,而土壤湿度取决于降水模式[15]。也就是说,温度对植被生产力的影响通常也与降水有关。一项在青藏高原海北草原的研究认为高寒草甸草原的地上净初级生产力与温度没有显著的相关性,而是与生长季降水量极显著相关(P< 0.01)[16]。原因是,在植被的生长季时期,草地处于迅速增长的阶段,需要土壤为植被的生长提供大量水分来进行光合作用,进而提高生产力。有研究认为非生长季的降水量也显著影响的植被的生产力(P< 0.05),这是由于草原进入冬季后,降水的主要形式为降雪,积雪的厚度和覆盖面积会影响植物越冬芽的存活,进而影响下一年的植被生产力[17]。这与李英年等[18]的研究结果一致,即水分是限制草地生产力的主要因素。同样的,在新疆地区开展的一项草地净初级生产力对气候变化的响应的研究结果表明新疆地区草地的生产力主要受夏秋两个季节的降水影响,与温度的相关性较低[19]。穆少杰等[20]关于内蒙古草地生态系统的研究认为,降水量是内蒙古草地植被生产力的主要影响因素,分布在半湿润和半干旱过渡气候带的草甸草原生产力和温度有较高的相关性,低温是植被生长的主要限制因素。分布在干旱少雨地区的典型草原和荒漠草原的草地植被生长主要受降水的影响,其中荒漠草地生产力与降水量的相关性更强。
总的来说,在全球气候变暖、降水格局改变的背景下,区域植被生产力变化对气候变化的响应存在着区域差异。一些研究的草地生产力和温度与降水呈正相关关系[21]。相反,有一些区域的研究表明草地生产力的温度呈负相关关系[22]。在气候变化的研究领域得出两种相反的观点的原因可能是,不同研究区域的季节性气候与年度气候变化之间存在着很大的差异,而不同区域植被的生产力对不同时期的气候变化的响应也有所不同,例如春季温度的升高会导致一些受温度限制的研究区域的冻土提前融化,加速土壤有机质的分解并降低土壤肥力,对植被生产力产生负面影响;而在另一些区域,春季气温下降和降水增加容易造成春季霜冻,在生长季节影响植物的生长。一项位于中国干旱和半干旱地区研究表明,冬季气温和夏季降水是整个研究区域植被生产力的限制因素,春季气温和秋季降水在西部影响不大,但在东部两者却至关重要[22]。因此,季节性气候变量对植被生长的影响对于研究植被生产力非常重要,在未来的研究中,季节性气候变量对植被生长的影响应纳入考虑。
另外,大多数探索植被生产力对温度升高的响应的研究没有考虑动物的影响,即放牧与增温的相互作用对草地生态系统的生产力、物种多样性以及土壤理化性质的影响。因此,在放牧条件下,气候变暖对不同区域的草地生产力及多样性等方面还存在很大的不确定性。
1.2.2 二 氧化碳
二氧化碳(carbon dioxide,CO2)主要是从土壤的水分利用效率、植物生理生态过程等方面影响植被生产力的。首先,大气中CO2浓度的升高会提高植物的水分利用效率,从而导致土壤水分利用率增加并减少植物的水分胁迫,因此CO2浓度升高会通过对土壤水分水平的影响来提高幼苗成活的成功率[15]。其次,有研究发现CO2浓度升高可以促进植被的光合作用,提高土壤中的微生物活性和呼吸[23],并减少蒸腾作用和植被的气孔导度[24],从而提高土壤水分利用率,促进植被产量的积累。同时,CO2浓度升高会提高植物的氮需求,从而改变凋落物的碳氮比和分解速率,继而影响生态系统的生产力[25]。研究表明,空气中CO2浓度的升高可以增加干旱条件下作物的生物量累积,尤其是地下生物量有显著增加(P< 0.05)[26],但对于水分充足条件下的作物影响不大[27]。
目前有关CO2对草地生态系统中生产力、土壤养分、温室气体循环等方面的研究大多为田间试验或模型预测,而田间试验的时间一般为3~8 年,这对于需要长期监测过程的试验来说相对较短,例如植物群落物种的变化、土壤碳氮的变化等,这些方面对于CO2的响应可能会随着时间产生变化并得到不同的结果,因此有关CO2在较长的时间尺度上如何影响草地生态系统的生产力等方面仍需进一步研究。
1.2.3 氮 沉降
氮沉降是陆地生态系统中除气候变化和大气CO2浓度升高之外对草地植被生产力影响较大的因素之一,近年来,大气中氮沉降量明显增加,氮也成为草地植被生长和生产力的主要限制因素。
大部分研究表明,氮沉降可以显著提高草原生态系统的生物量和净初级生产力(P< 0.05)[28],但存在氮临界负荷,即当氮沉降的量到达一定量后,植被的生产力将会有所降低[29]。
氮添加也会对植物和土壤群落产生负面影响,有研究定量分析了氮沉降对于全球草地生态系统的影响,结果表明虽然氮沉降提高了草地的生物量和生产力,却会造成植物多样性降低。这是由于外部氮素的添加会扰乱生态系统的平衡状态[30],因此造成物种对于生态系统中的营养物质的不平衡竞争,一些物种可能会逐渐被另一种物种所取代,进而导致物种丰富度的降低[31]。同样有研究表明,在高水平施氮时,氮添加可能会利于一年生物种生长或外来物种的入侵导致原生物种的损失[32]。
具有不同管理制度的地区和不同气候条件下的生态系统可能以不同的方式响应额外的氮,在湿润的草原环境下,植被的生物量和生产力受氮沉降影响更为显著,这说明气候条件适宜的情况下,外部添加的氮素更易被植物所利用[32]。
生态系统中输入的氮素分为自然界大气氮沉降和放牧、施肥等人类活动输入的氮,这些氮沉降的成分复杂,大致可以分为无机氮和有机氮。不同配比的两种氮沉降可能会对草原生态系统的生产力和生物量产生不同的影响。有研究表明,不同配比的无机氮和有机氮均可使草地的地上净初级生产力增加,而不同配比处理之间的无显著差异,但无机氮的含量越多,对于物种丰富度的影响越大[33]。
气候变化和人类活动并不是作为单一因子对草地生态系统的生产力和生物量产生影响,目前的研究结果表明,在自然降水条件下,氮和水为共同的限制因素,因此无论是氮添加或是降水增加都会使植被初级生产力和生物量增加。而放牧和气候变化也会相互作用影响氮动态。降水、温度、CO2的变化以及氮沉降等影响因子的交互作用对草地的影响仍值得探究。
2 草地生产力的估算方法
2.1 气候相关统计模型
有关用于模拟植被生产力的模型的研究开始于传统的统计模型,气候相关的统计模型只考虑一些主要的气象因子,如温度和降水等,建立气候因子和植被生产力之间的简单统计回归模型,使用大范围尺度上的气候数据和植被统计数据,如Miami 模型、Thornthwaite Memorial 模型、Chikugo 模型、分类指数模型等(表1)。气候相关的统计模型在研究较大区域尺度的植被分布规律方面有很大优势。然而,气候相关的统计模型主要以简单的气候因子为理论基础,不涉及植物的生理生态学过程,而不同地区的植被生产力对于气候因子的响应有所不同。因此仅使用气候因子作为输入参数的统计模型估算植被生产力可能会产生较大的误差,目前常使用气候相关的统计模型来估算植被的潜在生产力。
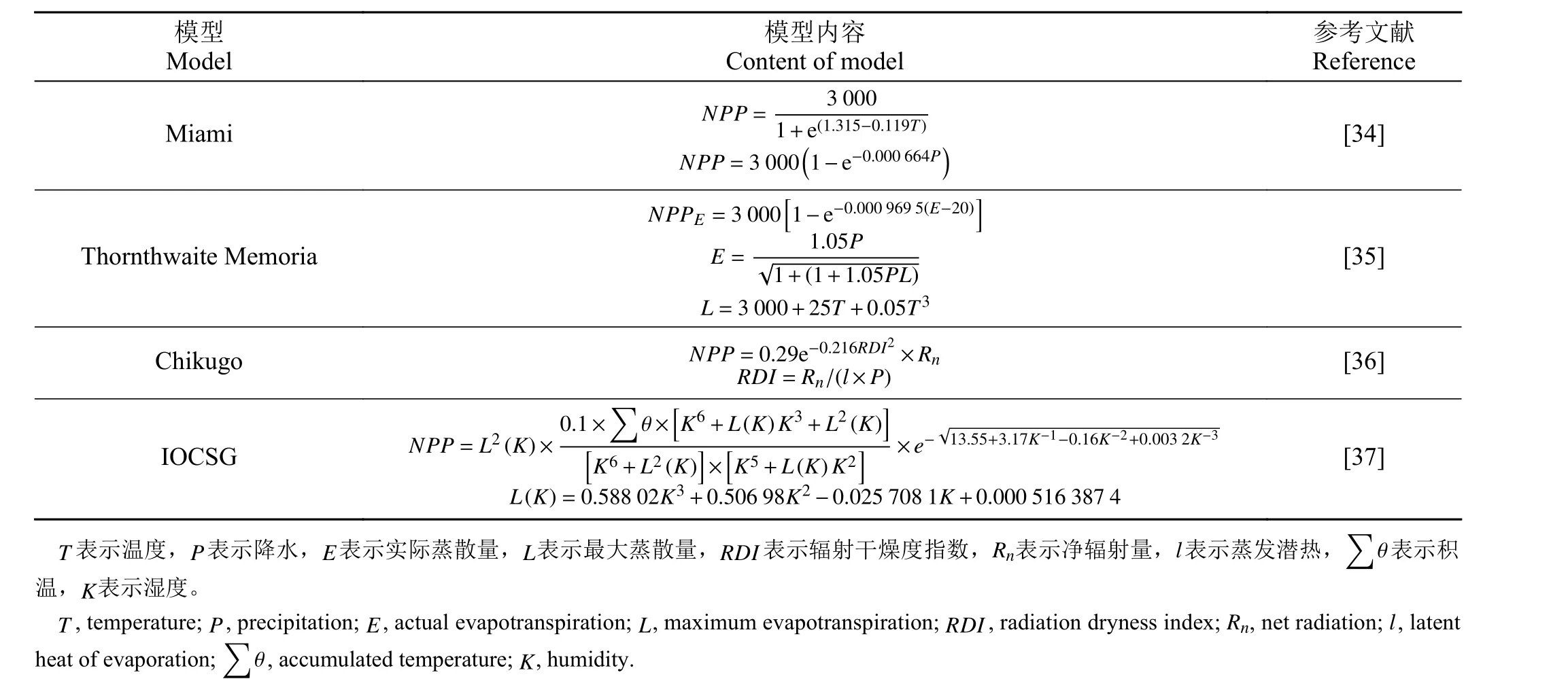
表1 不同统计模型的比较Table 1 Comparison of different statistical models
2.1.1 Miami 模型和Thornthwaite Memorial 模型
Miami 模型是最早用来估算生态系统植被生产力的经验模型,它是一种使用环境变量的数学模型,Miami 模型以年平均温度(T)和年降水量(P)这两个最常用的气候变量作为输入参数,使用最小二乘法建立年均温和年降水与植被净初级生产力之间的公式,使用时通常分别计算由上述两个环境变量得到的生产力,再根据Liebig 最小因子定律取较低的值为输出结果[34]。
Miami 模型使用的是单因子公式,仅考虑气候因素中的温度和降水,而植被生产力除了受温度和降雨的影响之外,还受其他气候因素影响。因此,Lieth 和Box 在Miami 模型的基础上,引入了蒸散量因子,提出Thornthwaite Memorial 模型,其输入参数包括年实际蒸散量、最大蒸散量、年降水量和年平均气温,其中E表示实际蒸散量[35]。
Miami 模 型 和Thornthwaite Memorial 模 型 都 是基于大范围实测数据建立主要气候因素和植被生产力之间的统计模型,估算方法简单,所需参数少,在有关估算植被生产力的研究初期,这两个模型被广泛应用于较大区域尺度的植被生产力的估算。国内有学者基于长期的气候数据,使用Miami 和Thornthwaite Memorial 模型估算内蒙古农牧交错带的草地生产潜力对不同气候变化情景的响应,分析草地生产潜力随气候要素的变化规律[38]。
2.1.2 Chikugo 模型
Chikugo 模型基于植物生理生态学基础,引入了辐射干燥度、净辐射通量、蒸发潜热等参数,使用国际生物学计划(International Biological Program,IBP)研究期间的全球植被生产力数据和气候数据进行统计分析,建立了根据净辐射量计算植被生产力的半经验半理论模型[36]。该模型的优点是考虑了几个主要气候因子的综合影响,但Chikugo 模型是基于土壤水分充足、植被生长茂盛的条件通过确定植被水分利用效率来计算植被的生产力,对全球大多数地区不具有普适性。因此Chikugo 模型在估算干旱、半干旱地区的草地生产力时会产生较大的误差[39],我国的学者根据多年气候数据,使用多种气候生产力模型对西北地区草地的生产力进行估算,结果表明Chikugo 模型的估算结果明显偏大(P< 0.05)[3]。
2.1.3 IOCSG 模型
草原综合有序分类系统(Integrated Orderly Classification System of Grassland,IOCSG)是基于定量化分类的草地分类系统(Comprehensive and Sequential Classification System of Grassland,CSCS)的 数 学 模型,也称分类指数模型[37]。该模型不需要使用遥感植被指数,根据年积温和年降水量指标可以将研究区域分为不同的类别,根据草地的位置和类型计算出相对应的草地生产力,目前有很多学者使用分类指数模型研究草地生产力及生态系统碳排放[40],有助于了解气候和植被之间的动态变化关系。但由于根据CSCS 方法划分的草地类型,是假设在当前的气候条件下生态系统不受人类活动干扰的潜在植被类型。因此,在验证模型时,存在如何分析和评价现实草地类型与潜在草地类型之间的关系的问题,例如植被受人类活动干扰的强度等。另外,将该分类指数模型推广至全球,以及探索进一步将草地类别细化的分类体系是未来草原综合有序分类系统研究的重要内容。
2.2 光能利用率模型
随着遥感和计算机技术的发展,利用遥感技术获取植被、气候等参数为估算草地生产力提供了重要手段,光能利用率模型,也称参数模型,由于卫星技术可以提供大范围尺度的植被、气候数据,使用光合利用率模型可以较为准确地估算区域以及全球尺度的植被生产力。该类模型是基于植物的光合作用过程及相关影响因子来估算植被生产力,目前常用的参数模型有CASA 模型、C-FIX 模型等(表2)。参数模型具有数据获取便捷、空间范围广和输入参数少的优点,相关植被参数无需测量和试验过程,通过遥感信息中的植被指数的计算即可获得植被生长过程中的许多参数。光合有效辐射(photosynthetically active radiation,PAR)是使用光能利用率模型估算植被生产力的关键参数。植物的光合有效辐射(absorbed photosynthetically active radiation,APAR)主要是指波长在0.4~0.7 范围内,可以被绿色植物用来进行光合作用的太阳辐射能。

表2 不同光合辐射模型的比较Table 2 Comparison of different models of photosynthetic radiation
但使用遥感数据进行植被生产力估算仍存在以下不足:第一遥感数据主要针对大尺度范围的区域,对于站点数据不够精确,同时,遥感信息在获取的过程中受到很多因素的干扰,受限于获取影像时的气候状况和传感器精度;第二是由于遥感卫星开始应用的时间,大部分数据获取持续的时间较短,对于长时间序列的研究无法提供充足的数据支撑;第三是光能利用率模型无法从植物生理生态学方面解释植被生产力随气候变化的机制。
2.2.1 CASA 模型
CASA 模型使用的参数主要是光合有效辐射(APAR)和光能利用率ε,以此来计算植被生产力,

CASA 模型主要应用于区域和大尺度范围内的植被生产力以及生态系统碳循环等方面的研究。张美玲等[47]将CSCS 的湿润度指标和 > 0 ℃的年积温引入CASA 模型,使用改进后的CASA 模型模拟并分析了2004-2008 年我国草地的生产力的时空变化,研究在实现草地分类与生产力模拟的关联方面取得了进展。研究结果表明水热条件是决定草地生产力的关键因素,不同草地类型的生产力存在较大差异,且对温度和降水的变化响应不同。这与杨红飞等[48]使用CASA 模型模拟新疆地区和内蒙古地区的植被生产力的研究结果一致,即草地生产力的空间分布特征主要受到水热条件的制约。
2.2.2 GLO-PEM 模型
GLO-PEM 模型是考虑了植物的光合作用和呼吸作用等植物生理过程的光能利用率模型,几乎完全由遥感数据所驱动。
式中:st和 为Nt分别表示t时刻的入射光合有效辐射和被植物所吸收的光合有效辐射比例, ε表示光能利用率,R为植物的自养呼吸[43]。GLO-PEM 模型大多用来模拟森林生态系统以及区域尺度的植被净初级生产力。
2.2.3 VPM 模 型
VPM 模型使用遥感数据和涡度观测碳通量数据,并考虑植物的光合有效辐射来估算植被总初级力。
式中: ε为最大光能利用率,f(T)、f(w)、f(p)分别模拟了温度、水和物候对植被生产力的影响,PAR为光合有效辐射,FPAR是植物吸收的光合有效辐射的比例[42]。郑艺[49]使用遥感和地面观测数据,基于光能利用率模型评估在植被生产力的估算过程中的参数不确定性,改进了该模型中的f(w)的计算,使VPM模型在草地和农田生态系统中的估算精度显著提高(P< 0.05)。张馨元[50]在草地和农田生态系统GPP 模拟的研究结果表示,经验参数(如植物光合的最高最低最适温度)不是影响VPM 模型模拟精度的主要因素,而对遥感数据进行去噪等处理可以提高模型的精度。VPM 模型在使用时通常需要对其中的参数重新调整,使其适用于不同植被类型的研究区域。
2.3 生态系统过程模型
生态系统过程模型以生态学理论为基础,根据植物基本的生理、生态学过程,模拟植被的生命活动,如通过光合作用、呼吸作用、蒸腾作用,以及有机物分解和循环等过程。并且,结合气候和土壤数据等来估算植被生产力,有利于揭示草地生态系统与气候变化之间的相互作用关系,不仅用于估算生态系统生产力,还常用于模拟生态系统碳氮排放等过程,目前常用的生态系统过程模型有DNDC、CENTURY、Biome-BGC、BEPS 等(表3)。但由于生态系统过程模型考虑比较复杂的环境因子和植被参数,导致模型需要输入和计算的参数较多,模拟过程复杂,导致该类模型在获取定量化参数方面有些困难,因此在进行大尺度区域和全球范围内的模拟时有一定的局限性。

表3 不同过程模型的比较Table 3 Comparison of different process models
2.3.1 CENTURY 模型
CENTURY 是用于模拟植被生产力和土壤有机质变化的生物地球化学模型之一[52]。模型包括3 个子模型,分别是植物生长子模型、土壤有机质及养分循环子模型、温度和水分收支子模型[17](图1),模型的输入参数有气象数据、研究区域的土壤理化性质和控制参数、植被生理生态参数等。该模型参数在不同区域的草地生态系统的普适性较高,CENTURY 模型可以用来模拟土壤中各种有机质变化[56]和草地生态系统的生产力,也可以模拟环境因子和管理措施(如放牧、施肥和刈割等)的变化对生态系统造成的影响。CENTUEY 模型的时间分辨率为月步长,使用的参数大多为月平均数据,该模型不仅可以用来模拟草地生态系统的生产力,还被广泛应用到农田、森林生态系统中。

图1 CENTURY 模型结构Figure 1 Structure of the CENTURY model
郭燕云等[2]基于新疆各气象试验站站点的多年草地观测数据,通过比较生物量的模拟值和实测值,对CENTURY 模型中会对植被生产力有较大影响的关键参数进行训练和验证,研究结果表明CENTURY 模型在天山山区有较好的适用性。耿元波等[16]基于多年气象数据,使用CENTURY 模型模拟青海省海北高寒草甸地上净初级生产力的年际动态变化,研究结果表明,CENTURY 模型适用于高寒草甸有关植被生产力的模拟。王松等[17]以内蒙古典型草原为研究对象,用过去多年历史气候数据和土壤数据,应用CENTURY 模型模拟该区域植被生产力的变化,认为影响模型精度的参数主要是研究地点的土壤质地信息、植被生长中的过程参数以及适宜植物生长的最适和最高温。另外,CENTURY模型中缺少厌氧环境的模拟过程,因此不适用于湿地生态系统, 但也有研究通过调整模型中的代表厌氧条件的参数使其成功应用于湿地生态系统的碳库动态变化模拟[57]。
CENTURY 模型的局限性在于,首先CENTURY模型的时间步长是以月为单位,模型的输出结果是月平均值,而使用的实测数据是以天为单位的地上部分生物量,其次模型采用月时间尺度,没有考虑极端天气事件对草地生产力的影响,除此之外模型没有考虑除气候因素外其他影响草地生长的因素,如病虫害等,没有综合考虑生态系统中的生物因子与气候因子的相互作用对草地生态系统的影响。
2.3.2 Biome-BGC 模型
Biome-BGC 模型最初开发为森林动力学模型,经过多位学者多年的发展和补充,将其发展为可以模拟不同生态系统植被和碳氮储量变化的过程模型[51],可以通过模拟生态系统中的碳、氮、水等循环过程来估算植被生产力,常用于森林生态系统模拟。Biome-BGC 模型包括了冠层辐射、光合作用、蒸腾作用、和呼吸作用等模块[58](图2),输入参数包括研究区域的基本信息,如经纬度、海拔高度、土壤质地、植被类型,和每日最高最低气温、降水、太阳辐射等气象数据,以及植被碳氮比、气孔导度等植物生理生态参数。

图2 Biome-BGC 模型结构Figure 2 Structure of the Biome-BGC model
李传华等[58]使用参数本地化后的Biome-BGC模型,模拟青藏高原五道梁地区的草地净初级生产力,研究结果表明在该地区,温度是植被生产力的主要限制因子,温度和降水交互影响该地区的生产力。有研究通过改进Biome-BGC 模型中的冻融循环过程,使其能更好地应用于模拟高寒地区植被生产 力,显 著 提 高 了 模 型 的 估 算 精 度(P< 0.05)[59]。Biome-BGC 模型一般用于模拟天然植被的碳氮循环过程,模型中没有设置适用于农作物的生理生态参数,也不考虑一些放牧、收割等人为管理措施,因此在使用Biome-BGC 模型模拟农田作物的碳循环和生产力时需要对草地的生理参数进行修改。所以有学者将遥感数据引入Biome-BGC 模型,用来代替模型模拟的叶面积指数,并修改模型中的草地参数使其适用于农田生态系统的模拟[60]。
2.3.3 DNDC 模型
DNDC 模型即反硝化、分解模型,是一种生物地球化学模型,它通过计算反硝化和有机质分解过程来模拟生态系统中的碳氮循环过程[53]。DNDC 主要由两部分组成(图3),第一部分包括土壤气候、植物生长和有机质分解3 个子模型,在这个部分中,模型通过输入的气候、土壤、植被等参数模拟生态系统中环境因素的变化;第二部分包括硝化、反硝化和发酵3 个子模型,在这部分的主要作用是分析第一部分中的环境变化,进而模拟生态系统中的各类生物化学反应[61]。经过长期发展及多位学者的扩展和补充,DNDC 模型目前可以用来计算不同生态系统中(如农田、草地、森林、湿地等)的温室气体排放、作物产量以及土壤有机碳含量等[62-64]。DNDC模型在点位尺度和区域尺度方面的应用以及验证的研究都有很多。
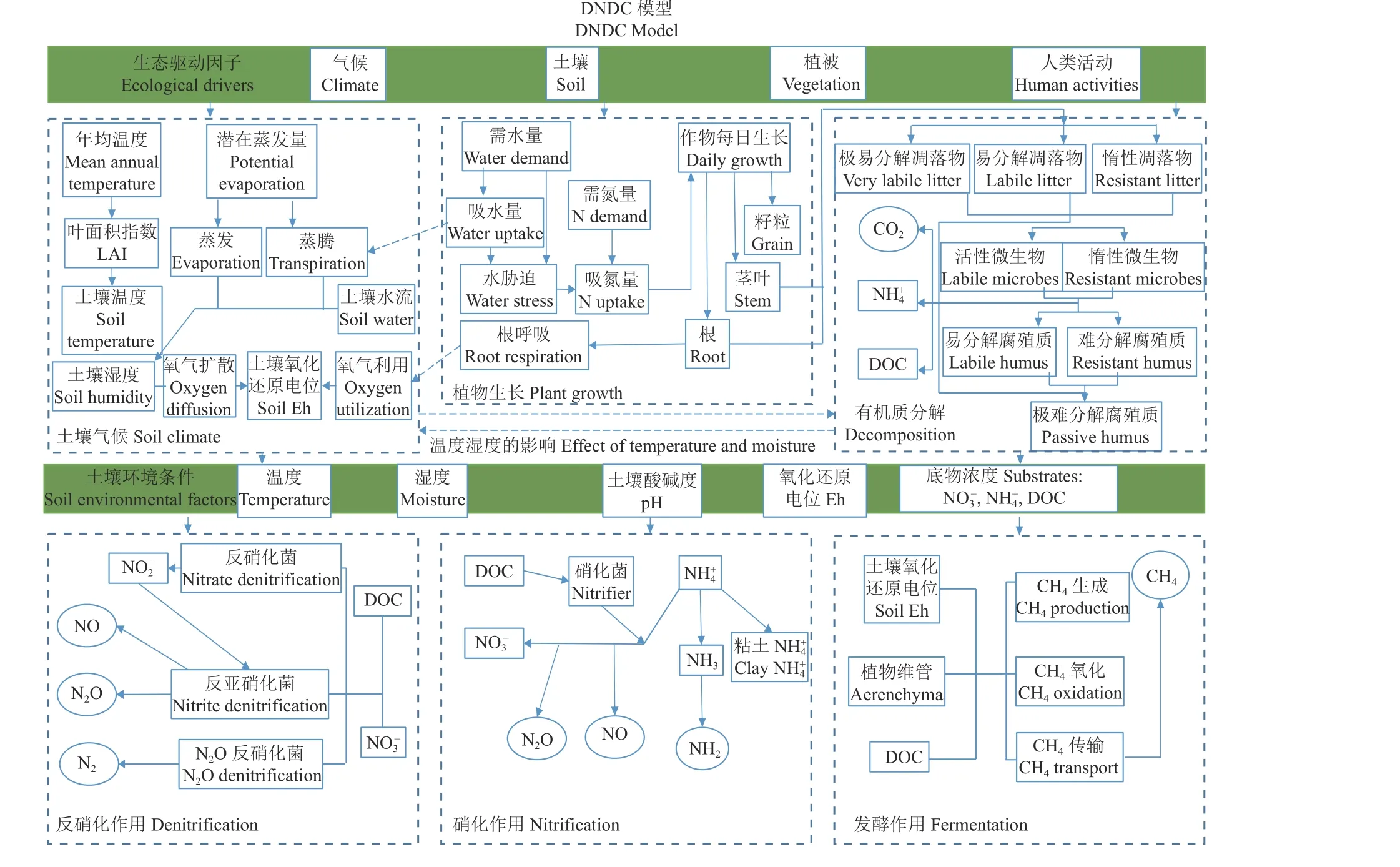
图3 DNDC 模型结构Figure 3 Structure of the DNDC model
有学者对DNDC 模型雨雪分配、土壤温度、土壤冻融等模块进行改进,将其应用于加拿大TGASMAN 站点,使其更好地预测寒冷地区土壤温度和温室气体排放[65]。王多斌[66]采用增温-放牧试验方法,使用DNDC 模型分析气候变化和放牧对青藏高原草甸土壤有机碳含量的影响,研究认为气温升高和放牧会对SOC 产生负面影响,而气候变化是导致该生态系统中土壤有机碳变化的主要因素。有学者基于时间序列的遥感数据量化放牧强度,将其结合DNDC 模型,应用于我国若尔盖草原探索放牧对草地生产力的影响,结果表明考虑放牧的生产力与实测生产力数据显著相关(P< 0.05)[67]。
DNDC 模型的优势在于,可以反映生态系统中土壤-气候-作物之间的相互作用以及碳、氮循环过程,尤其可以较为精确地模拟产量、气体排放等状况,该模型在点模拟和区域模拟方面都发挥了重要作用。然而模型虽然考虑到了环境因子和植被的生长过程,但在模拟各因素之间的复杂关系和相互作用方面还不够完善,不能很好地掌握不同类型作物的生长状况。
3 讨论与结论
基于气候相关的统计模型在研究初期由于其模型方法简单、输入参数少因而得到了广泛的应用,由于其仅考虑研究区域气候因素而不考虑植物自身生长的生态生理过程,目前常被用来估算大范围区域内的潜在植被生产力。随着遥感和计算机技术的发展,利用遥感获取植被、气候等参数为估算草地生产力提供了重要手段。光能利用率模型主要考虑植被生产力和植物所吸收的光合有效辐射之间的关系,其中的输入参数可以直接从卫星数据中获取,省略了许多耗费人力和成本的取样和试验过程,但光能利用率模型中的输入参数通常是以区域形式输入,难以获取高分辨率下的站点数据,所以在点验证方面有一定的局限性,且该模型对于遥感数据获取和预处理的精确度要求较高,卫星的传感器性能、天气状态等都有可能造成较大的误差。过程模型不仅考虑了环境中的气候、土壤因素,还纳入了作物生理及物候学参数,不仅可以用来估算植被生产力,还可以模拟生态系统中的碳氮循环过程以及土壤呼吸和土壤有机碳的动态变化过程,从而揭示生态系统中植被和环境的相互作用关系,但过程模型原理复杂,需要输入的参数很多,各因素之间的相互作用及其参数的不确定性可能会使模型的估算结果有误差。
有关估算草地生产力的研究经过多年的发展和改进已经取得了很多进步,国内外的学者根据研究区域气候特征、植被类型、土壤环境等不同条件也开发和改进了许多适用各类草地生态系统的估算模型。除了本研究中提到的统计模型、光能利用率模型、生态系统过程模型等,还有很多学者将不同模型进行耦合,使其同时兼具两种模型的优点,避免了使用单一模型进行模拟的局限性,使得模型估算的精度有了很大的提高。例如过程模型由于需要模拟植被的生长状况以及环境条件,需要获取复杂且大量的参数作为模型的输入参数,但受到空间和时间尺度的制约,气候参数、植被参数和土壤参数等在获取时受到技术以及时间和人力的制约,而遥感技术的发展为数据获取提供了技术支持,很多难以通过直接取样得到的参数可以通过卫星观测数据进行计算,例如叶面积指数、植被覆盖度、NDVI等。因此,将遥感手段与过程模型相结合,使用卫星数据代替难以获取的实地采样数据,并将模拟扩展到区域和全球尺度上,将是未来有关草地生态系统研究的发展趋势。例如BEPS 模型,基于FOREST-BGC机理模型,将输入参数中的叶面积指数使用遥感数据反演得到,实现遥感和过程模型的耦合[68],以及GLOPEM-CEVSA 模型,是将过程模型CEVSA 和光能利用率模型GLO-PEM 进行耦合,以叶面积指数作为中间参数简化了模型中的生理生态过程,使其兼具两者的优势[69]。另外,如何根据不同特征的研究区域选择合适的模型和参数也是未来草地植被生产力研究的重要内容之一。
但是,在有关植被生产力方面的研究迅速发展的同时,仍存在一些问题值得思考和改进。首先是人类活动和气候变化如何影响草地生态系统的植被生产力以及气体排放这一课题中,大多是温度和降水的双因子交互控制试验,多因子(如温度、降水、氮沉降、放牧等)之间的交互作用方面的研究还比较匮乏。
此外,模型精度的验证方面也有一定的困难,区域甚至更大尺度的草地生产力的精度如何验证一直是模型估算研究中的难点之一。目前的研究中,模型验证的主要方法是与野外实测数据或站点数据对比,但由于部分地区环境恶劣难以进行人工采样,以及当研究区域范围较大时,难以获取大量的实测数据进行验证。
