高寒草甸功能群组成及营养品质对不同放牧强度的响应
2024-03-14张正义张振豪苗百岭侯扶江程云湘
张正义,胡 逸,张振豪,苗百岭,侯扶江,程云湘, 4
(1.内蒙古大学生态与环境学院, 内蒙古 呼和浩特 010021;2.内蒙古自治区气象科学研究所, 内蒙古 呼和浩特 010000;3.兰州大学草地农业科技学院 / 兰州大学草地农业生态系统国家重点实验室, 甘肃 兰州 730020;4.蒙古高原生态学与资源利用教育部重点实验室, 内蒙古 呼和浩特 010021)
青藏高原的高寒草甸承载了高原畜牧生产主要资源供给,也是全国生态系统类型和植物多样性保留相对完好的一块净土[1-2]。温室效应、人类活动等因素诱发了草地沙化、斑块化、盐渍化以及外来种入侵等问题,这些问题会影响草地生态系统的生物多样性以及生态系统服务与功能,导致部分高寒草甸地区退化[3]。因此,探究合理的牦牛放牧强度对恢复草地生产力和物种多样性,以及群落功能群组成和牧草营养品质随牦牛放牧的变化规律,进而揭示放牧草地的退化机制具有重要意义。
适度放牧对草地植物生长和群落物种多样性有一定的促进作用,使得草地群落处于较稳定状态,确保草地生态系统可持续发展[4-5]。而过度放牧造成地上生物量显著下降,群落高度及物种多样性下降,物种结构趋简单化,使群落主要植物种功能群原本稳态的优势地位被杂类草替代,使草地种群的生境遭到破坏[6-7]。秦洁等[8]发现,在内蒙古不同草地类型中羊草(Leymus chinensis)的生物量会随着放牧强度的增加而减少。卫智军等[9]的研究指出重度放牧会导致草地质量降低。段敏杰等[10]对西藏那曲地区紫花针茅(Stipa purpurea)草原的研究也证明了高强度的放牧行为会降低群落盖度及地上生物量。但从放牧对植物营养影响的角度考虑,Bai 等[11]和Schönbachp 等[12]的研究却发现放牧活动能够使牧草的部分营养价值增加。
不同的研究发现,相比于物种多样性,功能多样性可以更好地揭示生态系统的多功能性[13-14]。将植物按照相似的属种关系和生长特性划分为不同的功能群类型,划分后的功能群将更有利于从大尺度上揭示放牧对草地群落结构的影响。同一功能群的物种在植物生长阶段和所含营养成分具有高度的相似性,而由于草地生产力和结构的变化最终会体现在营养成分和各功能群占比的变化上,这同样将有助于揭示放牧活动中家畜不同的采食方式和偏食性对草地群落结构和草地生产力的影响[15]。本研究主要集中在高寒草甸植物群落特征,功能群成分变化,牧草营养品质及其在不同强度放牧干扰条件下的相互关系,以反映群落在放牧干扰下的变化过程,分析放牧活动对植物群落的影响,深化对放牧干扰下的草地功能群演替过程的理解。
1 材料与方法
1.1 研究区概括
该研究在甘南藏族自治州玛曲县阿孜畜牧科技示范园区进行,平均海拔3 500 m,年平均气温1.2 ℃,年均降水量620 mm (图1)。植被类型为典型高寒草甸和高寒灌丛草甸,植被于每年5 月开始返青,8 月达生长期旺季,9 月下旬逐渐枯黄。土壤类型为亚高山草甸土。主要以青藏苔草(Carex moorcroftii)、棘豆(Oxytropis)、高山嵩草(Kobresia pygmaea)、莓叶委陵菜(Potentilla fragarioides)和垂穗披碱草(Elymus nutans)为主。

图1 2016-2017 年研究区月均温和降水量Figure 1 Average monthly moderate precipitation in the study area from 2016 to 2017
1.2 试验设计
2016 年6 月,在研究区附近选取利用情况相似、植被较为均一的3 户牧场作为试验样地。以每家牦牛数量、牧场放牧面积,结合牦牛在放牧时行走范围距离棚圈的远近,畜圈为中心向外辐射大约250、500、1 000 m,将每家牧场划分出3 个不同的放牧强度区域,牧场围栏为边界,分别定义为重度放牧牧区、中度放牧区、轻度放牧区[16](表1)。相对放牧强度 = 牦牛存栏数/不同放牧强度区域面积,按照1 牦牛 = 5 个标准羊单位进行换算。以围封4 年的禁牧区作为对照。于2016 年和2017 年的6 月-9 月进行植被调查及土壤样品收集。在每个放牧区及对照区分别随机放置4 个0.5 m × 0.5 m 的样方进行群落调查。将样地内数量占比高的物种按照通用的草地植物群落功能群和经典的林奈生物学分类法[17]划分为7 个主要功能群:禾本科、莎草科、豆科、菊科、蔷薇科、毛茛科和其他杂类草。
1.3 群落调查和样品采集
记录每个样方内出现物种名称、物种数量、物种盖度、植株高度和生长期信息等。在植物调查结束后刈割收获地上生物量编号并带回实验室。在刈割后的样地随机选取4 个点,用直径5 cm 的土钻采集0-20 cm 表层土样测土壤鲜重,过筛分选出植物根系并去除砾石,对筛选的植物根系水洗、烘干,称量地下生物量数据。收获的植物地上部分经过烘干粉碎后用于植物营养指标的测定。
1.4 测定指标与方法
采用全自动凯氏定氮法测定粗蛋白(crude protein,CP)含量;采用AnkomXT 15i 全自动脂肪仪测定粗脂肪(ether extract,EE)含量;采用Ankom A200i 全自动纤维分析仪测定中性洗涤纤维(neutral detergentfiber,NDF)和酸性洗涤纤维(aciddetergent fibre,ADF)含量;采用灼烧灰化法(550 ℃马弗炉)测定粗灰分(Ash)含量[18]。
根据物种数目、物种的重要值,用以下公式计算群落多样性。
Margalef 丰富度指数:Ma=(S-1)/lnS。
式中:S为每个样方出现的总物种数,N为样方内的全部植株数量总和。Pi为样方内的第i种类群占整个群落的相对重要值。
1.5 数据分析
使用Excel 2010、SPSS 24.0 分析软件对不同放牧强度下的3 家牧户的地上生物量(aboveground biomass,AGB)、地 下 生 物 量(belowground biomass,BGB)、多样性指数、均匀度指数、丰富度指数及各牧草营养指标进行单因素ANOVA 模型分析;并对牧户Ⅰ、牧户Ⅱ、牧户Ⅲ内不同放牧强度下的植被情况,采用LSD 检验方法进行各处理间的多重比较。使用Origin 2021、BioRender 等软件绘制相关图表。
2 结果与分析
2.1 牦牛放牧强度对草地生产力的影响
牦牛放牧强度不同时,高寒草甸植被生物量变化有显著影响(图2)。结果表明,2016 年、2017 年两年间的变化趋势相对一致,牦牛放牧干扰对草地生产力(总生物量)有显著影响(P< 0.05)。随放牧强度增大,地上生物量呈现显著降低的变化趋势,其中可食部分显著递减,不可食部分也呈递减趋势,除轻牧和中牧处理间无显著差异外,对照、轻牧及重牧间不可食部分差异显著。2016 年对照区地上生物量显著高于其他放牧处理组,约404.98 g·m-2,可食部分约284.45 g·m-2,不可食部分约120.53 g·m-2。地下生物量随放牧强度增加呈单峰变化,轻牧处理最高,显著高于对照组和重牧组。
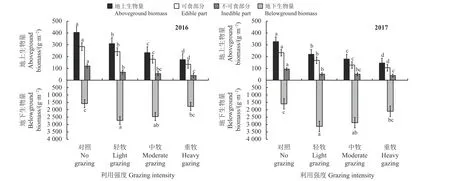
图2 2016 和2017 年牦牛不同活动强度对群落生物量的影响Figure 2 Effects of yak activity intensity on community biomass in 2016 and 2017
2.2 放牧强度对植物群落多样性的影响
放牧强度与物种多样性之间存在较强的相关性(表2)。2016 年群落多样性指数、均匀度指数和丰富度指数随着放牧强度的增大呈现先上升后下降的变化趋势,即3 组放牧处理下,中牧的多样性指数、均匀度指数和丰富度指数均高于轻牧和重牧;2017 年对照区域3 种指数显著高于3 组放牧处理。在放牧干扰下,2016 年与2017 年两年之间的植被多样性变化趋势也相对一致,并且与植被生物量的变化趋势略有不同,均以中牧处理的多样性最高。这可能说明中等放牧强度更利于维持群落较高的物种多样性和丰富度,而长期或过度的放牧干扰将会对物种多样性产生消极影响。
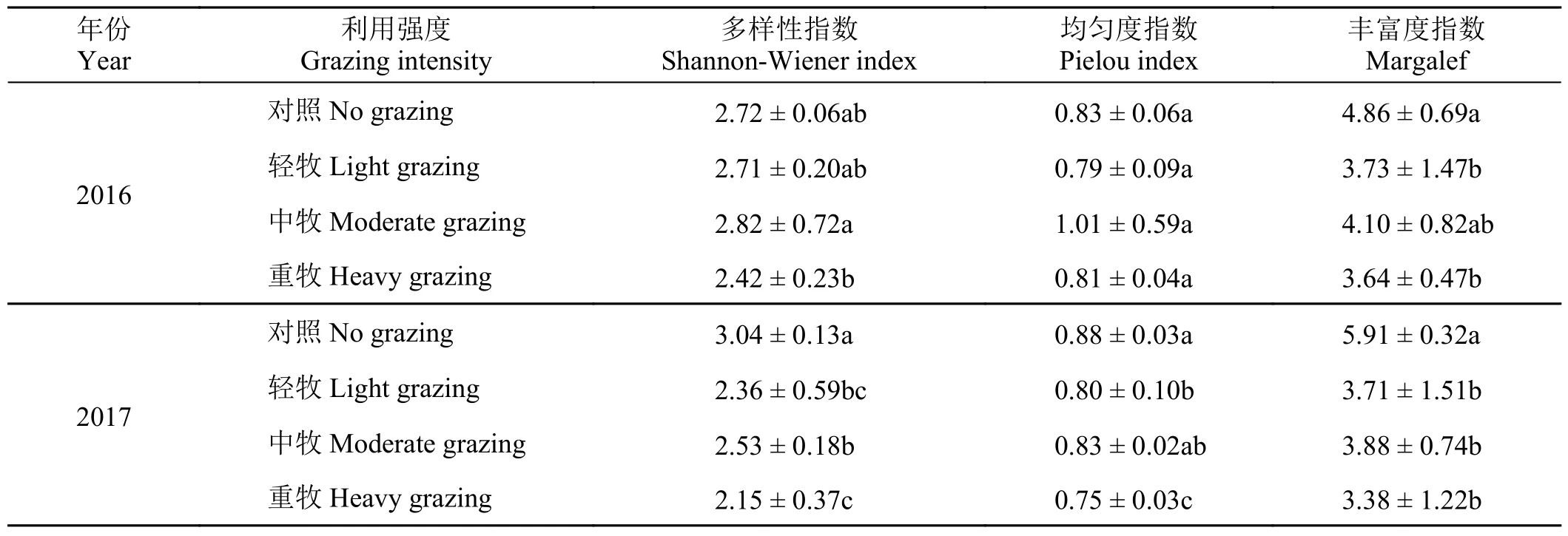
表2 2016 年和 2017 年牦牛活动强度对植物多样性影响Table 2 Effects of grazing intensity on plant diversity in 2016 and 2017
2.3 放牧强度对牧草营养品质的影响
放牧干扰与牧草营养品质之间有显著相关关系,但不同牧草营养指标对放牧干扰的响应情况不同(图3)。不同放牧强度下高寒草甸植被群落的粗蛋白、粗灰分含量存在差异,两者均随放牧强度的增加有增长的趋势;粗脂肪含量各放牧强度间无显著差异。中性洗涤纤维在2016 年随放牧强度增加呈先增长后下降的趋势, 而在2017 年则呈连续下降趋势;酸性洗涤纤维含量在2016 年在不同放牧强度间差异不显著,但在2017 年各放牧强度显著低于对照。

图3 2016 和 2017 放牧强度对牧草营养品质的影响Figure 3 Effects of grazing intensity on nutrient quality of herbage in 2016 and 2017
2.4 放牧强度对高寒草甸群落功能群比例影响
植被功能群组成的分析结果可知,在放牧条件下,3 家牧场的禾本科植物比例总体均低于对照组(图4),但莎草科植物比例总体高于对照组,并且整体上禾本科植物与莎草科植物在两年内均呈现此消彼长的状态,且变化幅度与放牧强度的变化存在明显的关联;豆科植物比例占群落体量较小,且在放牧干扰下豆科比例总体呈下降趋势,占比均低于对照组;除2017 年牧场Ⅲ以外,菊科植物的比例在3 家牧场两年的放牧梯度上都随放牧强度的增加而呈现先增长后降低的趋势;此外,3 家牧场的蔷薇科植物在放牧干扰下的比例均高于对照组,并随着放牧强度的增加,蔷薇科植物比例也有所增加;在牧场Ⅰ、牧场Ⅱ样地内,毛茛科植物比例均高于对照组,且随放牧强度的增加呈下降趋势。
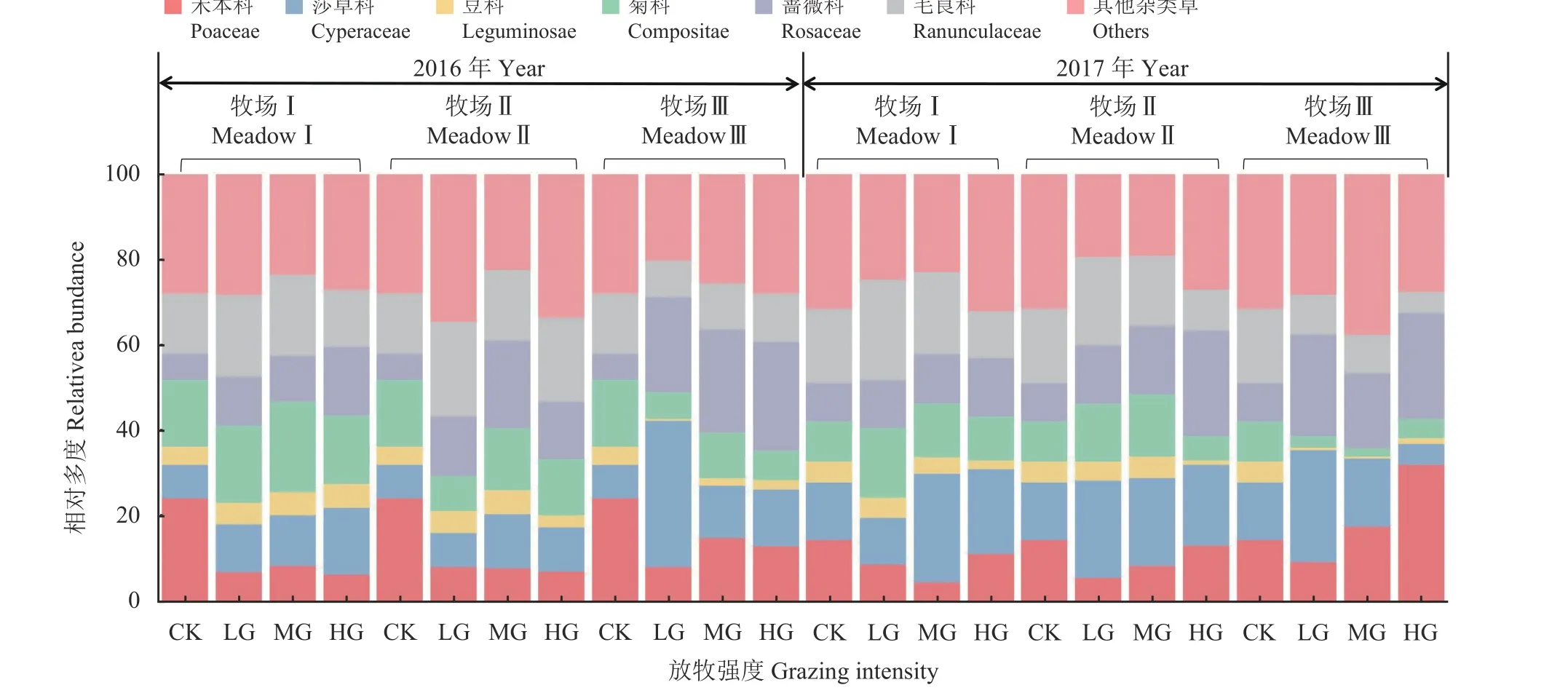
图4 放牧强度对植被功能群组成的影响Figure 4 Effects of grazing intensity on composition of vegetation functional groups
2.5 功能群比例与牧草营养品质的关系
相关性分析发现,高寒草甸牧草粗蛋白含量含量与禾本科及蔷薇科植物在群落内占比呈极显著正相关关系(P< 0.01) (图5),而与毛茛科、菊科及豆科植物在群落中占比呈显著负相关关系;中性洗涤纤维含量与莎草科植物功能区占比呈显著正相关关系,而酸性洗涤纤维含量则与毛茛科植物占比呈极显著正相关关系。此外,粗灰分含量与菊科和毛茛科植物功能群占比呈极显著负相关关系,与蔷薇科植物占比呈显著正相关关系。

图5 功能群比例与牧草营养品质Figure 5 Proportion of functional groups and nutrient quality of herbage
3 讨论
3.1 不同放牧强度干扰下甘南高寒草甸植被生物量与群落特征的变化
放牧行为是天然草地最主要、最直接的利用方式,牦牛的选择性采食会选择适口性好的牧草,通过采食直接使植被群落的生物量和多样性组成发生改变。李永宏[19]的研究表明,在拥有相似的气候条件下,植被群落结构主要由放牧所主导。群落各指标参数在放牧的干扰下进行变化,群落的变化情况也随之体现[20]。本研究结果表明,放牧会显著降低群落地上生物量,但增加了地下生物量,这可能是因为长时间牦牛采食及行走践踏导致植物地上的营养器官生长发育受限,采食导致叶面积指数减少,进而引发植物的再生率下降;家畜践踏对植物地下根系比地上部分影响大,但家畜采食对植物地下根系影响相对较小,被家畜干扰的植物可通过自身分配营养物质来增加地下根系生物量,这一过程在一定程度上可抵御家畜行为活动对草地带来直接的干扰胁迫[21]。另外,由于禾本科、莎草科植物易被牦牛采食,且牦牛通过高强度的选择性采食直接导致禾本科、莎草科等优质牧草的株高冠幅降低、地上生物量减少的同时,诸如鹅绒委陵菜(Argentina anserina)等不被家畜喜食的蔷薇科植物获得生长空间进而迅速生长繁衍,且随着数量增多占群落重要值变大,导致优良牧草在群落中的优势地位逐渐被取代,导致草地质量下降[22-24]。同样,牦牛采食后残留和新生的植物叶片也因自身的株高降低,使影响其生长发育的遮蔽物减少,直接暴露在阳光下的叶片更多,光合作用能力增强;且对根系在土壤中的垂直分布在一定程度上有所促进作用。
一般来说,植物地下生物量的变化可以体现植物对极端环境的适应特征,也是植被通过根系吸收营养物质并转化固定的重要体现[25]。本研究结果显示,有放牧干扰的样地地下生物量相比于禁牧样地均有增加,但增量随着放牧强度变大而逐渐降低,结合地上生物量随放牧强度增加而出现的减少,这可能说明放牧使群落的总生物量逐渐减少且逐渐向地下转移。随着牦牛放牧活动强度的增加,试验地植物地上生物量和群落的物种丰富度总体呈下降趋势。除家畜采食和践踏等行为可以直接作用于草地外,也与高度封闭的牧场中,长期进行牦牛的重度放牧行为有关,以至于高频率的采食消耗了牧场草地表层土壤覆盖物,导致土壤紧实度上升并丧失原有土壤透水性,不利于土壤吸收表层汇积的降水,使水分向下渗透受阻。再加上试验地海拔高、高原热辐射量强、草甸蒸发量大,也与区域性的物质能量的吸收转化速率不同等间接的因素有关,造成草地植物群落生产力和物种丰富度的下降,前人的研究同样佐证了这一点[4-5,26]。
物种多样性是研究草地生态系统结构与功能的关键因子,多样性指数是物种水平上多样性程度与异质性程度的度量[27]。家畜通过采食、踩踏、排泄等行为对植物进行干扰,可以间接促进植物种子的传播,群落组成进而发生变化。植被群落特征对放牧响应显著,但物种多样性对不同强度的放牧干扰响应程度不同。本研究结果显示,中等放牧强度下的甘南高寒草甸有着较高的多样性、均匀度、优势度和丰富度,群落的多样性指数随放牧强度的增大呈现先升高后降低的趋势,这与王炜琛[28]在呼伦贝尔草甸草原的放牧研究结果一致,同时符合中度干扰假说[29]。适度放牧在一定程度上可以保持草地生态系统功能,过度放牧则可能减少家畜喜食牧草数量,导致杂类草获得更多生存空间,进而植被群落发生退化。在以牦牛为单一畜种放牧的牧场,这种对植被群落的影响表现的更为显著。这是由于在适度的放牧干扰下,家畜对群落中优势植物的采食抑制了其竞争力,让弱势植物有了更多生存机会,增加了群落结构的复杂性,从而使得群落多样性上升[30]。同时,考虑不同的牧场地形,牦牛的活动路径、饮水点和畜圈差异化的设置也会影响牦牛采食和践踏时间,导致不同强度的活动行为,从而直接地影响植物群落的物种组成。从轻牧到中牧,再到重牧的植被优势度指数呈现先增高后降低的变化趋势的原因,除以上中度干扰假说之外,也可能是由于植被群落在牧场内的空间异质性分布,或者草地的补偿性生长在轻度放牧干扰下有着更高的效能,导致优势种生物量增加,而非优势种减少。生长季的降水和积温也存在着差异,这点也不能排除。
3.2 不同放牧强度干扰下植物群落功能群与牧草营养品质的变化
家畜对草地的利用程度由放牧强度的大小所决定,各植物功能群也由此发生变化[31]。群落物种组合平衡性表现可以在植物功能群组成上体现[32],牧场内适口性较好、家畜喜食的植物优先被选择采食,家畜行走活动也对草地踩踏,这一系列家畜的行为则会扰动草地原有的平衡。因此,随着家畜采食和行走等行为活动的增强,优势种植物优先被采食并衍生其他植物,致其在群落中的地位逐渐下降[33]。有研究发现,家畜对植物的选择性采食仅发生在物种尺度上,并且群落的功能群组成和分布也直接受家畜的选择性采食的强度所影响[34-35]。本研究中放牧对植物功能群的影响表现为,随着放牧强度增加,优质牧草(禾本科和莎草科植物)占群落比例变化幅度明显,其中,禾本科植物和原有优势物种占比逐渐降低,莎草科与禾本科植物之间呈此消彼长的趋势。
高寒草甸上过度放牧使群落内不同功能群的比例发生变化,例如禾本科所占比例降低,莎草科及毒害杂草类所占比例增加[9,36]。本研究结果显示,随着放牧强度的增加,牦牛对植物的采食加强,群落中垂枝早熟禾(Poa szechuensis)、禾叶嵩草(Carex hughii)等牦牛喜食、适口性较好的植物被大量采食,寸草苔(Carex duriuscula)等低矮的莎草科植物增加生存机会、获得生长空间,植物功能群由此也发生变化,该结果与闫瑞瑞等[37]和杨思维等[38]的研究结果一致。其中,蔷薇科植物具有强大的根系系统且地上部分更耐践踏,由于放牧使优质牧草大量被牦牛采食,在竞争植物减少和牦牛踩踏的促使下,蔷薇科植物得到快速繁殖发育,所以,随着放牧强度的增大,蔷薇科植物在群落内占比有所增加。同时,由于家畜采食的植物大多以禾本科植物为主,导致禾本科植物株高、冠幅和数量下降进而优势地位降低,使得杂类草有更多繁衍空间,所以在本研究中,禾本科植物与杂类草的功能群占比会呈现此消彼长的关系,该结果与姜林等[39]在若尔盖区域的高寒草甸研究结果一致。此外研究表明,牛采食端和消化端的生理结构更适宜摄入纤维含量比较高的牧草种类[40],但重度放牧样地面积相对较小,而样地内过多的牦牛数量导致了大量禾本科和其他纤维含量较高的植物被粪便和尿液所污染,同时家畜又对被粪尿污染的植物拒绝取食[41],这一原因可能促使了2017 年牧户Ⅱ、牧户Ⅲ的重度放牧样地出现禾本科牧草随放牧强度增大而增加的趋势。
随着放牧强度的增加,牧草中粗蛋白和粗灰分含量呈上升趋势,这与Bai 等[11]、Schönbachp 等[12]以及王艳芬和汪诗平[42]的研究结果一致,他们研究发现重度放牧会使草地牧草中的蛋白质含量增高。出现这种现象的主要原因是牧草的养分含量与其不同的生长阶段有密切的关系。重度放牧样地内的牧草被牦牛采食后,该区域内会新生许多的嫩草,而嫩草的粗蛋白含量相对较高,且重度放牧区域内的枯落物较少,所以重度放牧区域内植被的粗蛋白含量较高。轻牧或禁牧的情况下则与之相反,轻牧和禁牧样地内地上分布大量粗老组织,故牧草粗蛋白含量较低。其次,由于放牧强度的增加,土壤容重增高,群落地上生物量被牦牛大量采食,样地内养分被现有植被大量汲取,使得植被营养含量相对较高。
放牧导致禾本科、莎草科、菊科、蔷薇科和毛茛科植物功能群优势种的营养价值的季节性变化,这可能是由于放牧延迟了牧草的成熟和木质化的过程[43]。禁牧下植物的酸性洗涤纤维和中性洗涤纤维含量显著高于放牧处理的样地,这表明牧草的中性洗涤纤维含量对放牧存在着明显的响应。另外,已有研究表明牧草营养品质对家畜采食量也有影响。其中牧草中性洗涤纤维的含量对家畜采食起一定的制约作用,主要表现在牧草中性洗涤纤维含量高会增加消化过程所需的时间,提高牦牛饱腹感进而降低家畜采食频度和强度[44-46]。并且本研究结果表明,莎草科植物在群落内的占比与中性洗涤纤维含量呈显著正相关关系,这可能是中性洗涤纤维变化主要基于两个因素,其一是功能群因家畜采食胁迫而使群落组成成分发生变化,导致粗纤维含量高的物种分布有所扩散,其次是放牧干扰使植被群落各种不同功能群所占比重发生变化。
4 结论与展望
牦牛放牧对青藏高原东麓甘南高寒草甸植被群落地上与地下生物量、群落结构特征、牧草营养品质和功能群组成均有显著影响。轻度和中度放牧可以适当提高植被地下生物量和群落多样性。高强度放牧干扰会降低高寒草甸生产力及减少群落物种组成种类,使群落多样性降低。但放牧潜在促进地下根系年季生长,生物量通过地上转移向地下,某种程度上维持着高寒草甸总生产力的相对平衡。禾本科和莎草科为主的优势牧草及其他家畜喜食的植物在群落中占比随放牧强度的增加而逐渐变小。放牧行为提高了牧草中粗蛋白和粗灰分含量,即植被群落的功能群组成密切影响着牧草的营养品质。
综上,如若在牧草不同的生长季,再结合当季的水热条件,适当地控制放牧牦牛的数量,将有利于植被群落结构和功能的稳定性,并一定程度上促进植物的补偿性生长,同时改善牧草的营养品质,也同样有利于满足牦牛的营养与能量需求(图6)。因此,在长期以牦牛放牧为主的高寒草甸,通过调整牦牛的数量及其活动范围,合理地规划牦牛的放牧活动,对维持草地生态系统的健康与可持续发展十分重要。
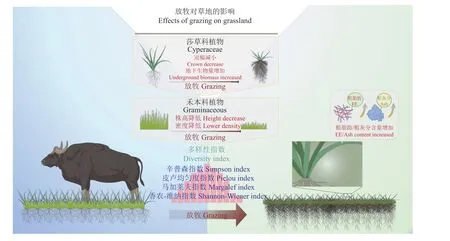
图6 牦牛活动对高寒草甸影响概念图Figure 6 Concept map of yak activity on meadow
