脓毒症相关性脑病发病机制的研究进展
2024-03-13曲艳平贺文廷范智东
王 旭 曲艳平 贺文廷 董 浩 范智东
1.大理大学临床医学院,云南大理 671000;2.大理大学第一附属医院麻醉科,云南大理 671000
脓毒症是由于机体对感染的应激反应失衡而导致的一种危及生命的器官功能紊乱综合征[1]。全球每年诊断脓毒症的患者总例数约为3 100 万,是全球重症加强护理病房患者入院和死亡的主要原因,约70%的脓毒症患者会并发脑功能障碍[2-4]。脓毒症相关性脑病(sepsis associated encephalopathy,SAE)是评价脓毒症严重程度、临床发展及转归的一个重要因素,通常是由全身炎症反应导致的弥漫性脑功能障碍。SAE 患者的死亡率明显升高,在急性期SAE 常表现为意识的急速变化,包括注意力缺陷、焦虑、行为异常、定向障碍、意识模糊、瘫痪、嗜睡、昏迷、伸肌强直及周围神经功能障碍等,随后可能导致长期或永久性的认知功能损害[5]。研究报道,在盲肠结扎穿刺(cecal ligation and puncture,CLP)小鼠模型中,存活的小鼠出现了急性脑病并伴有长期认知功能损害[3]。尽管SAE 引起的危害已经引起了临床的重视,但目前对其发病机制的研究仍缺乏深入的了解。本文主要对近年来关于SAE发病机制的研究进行综述,以期为SAE 更深入的理解及临床治疗提供参考。
1 SAE 的发病机制
1.1 血脑屏障(blood-brain barrier,BBB)破坏
SAE 发生的始动和关键环节是系统性炎症引起的BBB 损伤[6]。BBB 是中枢神经系统中一种复杂的血管多细胞结构,主要依赖于脑微血管内皮细胞及其间的紧密连接(tight junction,TJ)实现屏障功能,将中枢神经系统与系统循环相隔离,从而保持中枢神经系统的稳态环境[7-8]。脓毒症时,炎症介质主要通过模式识别受体靶向作用于BBB 上的内皮细胞[9]。内皮细胞内脂多糖(lipopolysaccharide,LPS)与Toll 样受体4 结合后释放细胞因子[10]。细胞因子、活性氧(reactive oxygen species,ROS)等介质活化并上调基质金属蛋白酶的表达,其通过降解TJ 蛋白(闭锁小带蛋白1、闭合蛋白、密封蛋白),以及连接黏附分子,引起BBB 通透性增加,炎症因子进入大脑,导致SAE 的发生[9]。此外,Yue 等[8]发现,肥大细胞激活可增加脑微血管内皮细胞对LPS 的易感性,CLP 诱导的SAE 小鼠海马中的肥大细胞过度激活,通过抑制肥大细胞激活可减少BBB 的破坏,提高生存率,改善认知功能障碍。综上,脑微血管内皮细胞损伤后,TJ 蛋白表达下降,造成BBB 损伤。这是SAE 发生发展的关键始动环节,炎症因子在其中发挥重要作用。见图1。
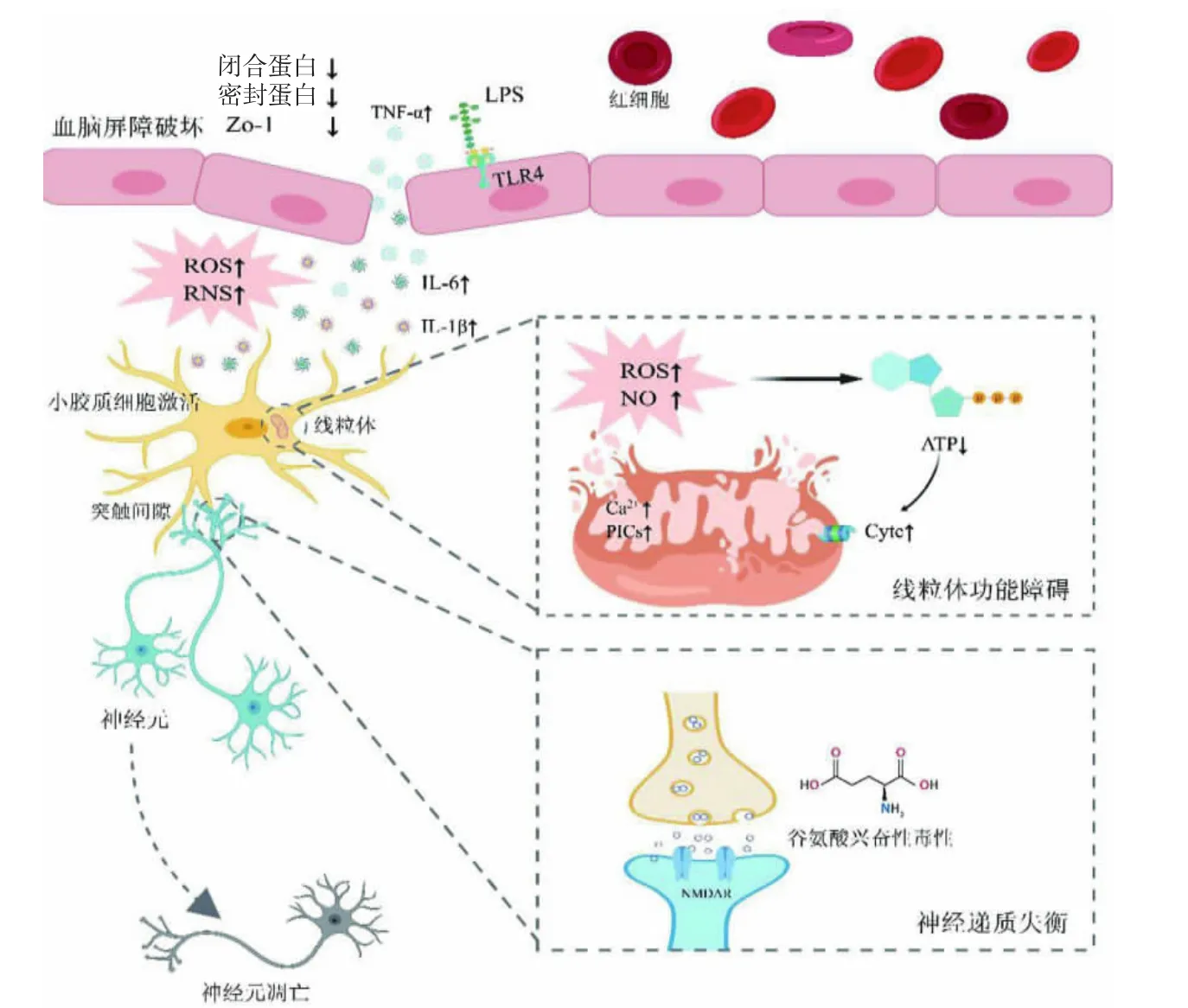
图1 脓毒症相关性脑病的发病机制
1.2 小胶质细胞激活
小胶质细胞是脑内固有免疫细胞,其活化是SAE发生发展的中心环节[11-12]。当BBB 被破坏时,静息状态的小胶质细胞迅速活化[10]。活化后的小胶质细胞分为经典M1 和替代M2 两种类型。LPS 或干扰素可通过诱导经典M1 型活化,促进核因子κB 和转录激活子1 的激活,上调小胶质细胞标志物CD86 和组织相容性复合体Ⅱ的表达,产生大量的ROS 和活性氮(reactive nitrogen species,RNS),并释放白细胞介素-1α、白细胞介素-1β、肿瘤坏死因子-α、白细胞介素-6 等多种炎症因子。然而,白细胞介素-4/白细胞介素-13可诱导替代M2 型活化,促进炎症消退和组织修复[13]。此外,小胶质细胞可通过过表达一氧化氮合酶或直接释放谷氨酸引起兴奋性毒性神经元死亡[14]。同时,小胶质细胞活化可通过CD40 和配体的表达,促进白细胞介素-6 和白细胞介素-1β 的合成,以及核因子κB的激活,从而导致炎症攻击的持续存在[15]。持续激活的小胶质细胞可增加炎症因子和ROS 的产生,从而形成恶性循环,加重了脓毒症BBB 的损伤[16]。因此,抑制小胶质细胞活化是SAE 防治的关键。
1.3 线粒体功能障碍
线粒体是维持机体正常生理活动的物质基础,其被视为机体能量代谢中枢。线粒体损伤是脓毒血症所致多脏器损伤的主要原因,线粒体稳态对脓毒血症患者的预后及存活具有重要意义[17]。线粒体通过持续运动、融合、分裂以保持其自身稳态,这种融合与分裂的稳态可对破坏的细胞器进行修复,以适应改变的能量需要,并适时启动凋亡[18]。然而,线粒体分裂/融合失衡可能是SAE 中神经细胞凋亡和氧化应激的主要原因[19]。在SAE 模型中,LPS 刺激会引起线粒体膜电位丢失,引发蛋白相关蛋白1 增殖,并招募抑癌基因P53,进而启动细胞死亡途径[20]。ROS、一氧化氮含量升高,可抑制线粒体腺苷三磷酸生成,进而引起细胞色素c 释放,引起神经细胞凋亡。RNS、一氧化氮、ROS等激活后,还可通过抑制电子传递链复合体Ⅰ、Ⅳ,导致线粒体功能紊乱,参与SAE 的发生、发展。同时,ROS/RNS 可增加线粒体与内质网的膜透性,上调Ca2+及促炎性细胞因子水平,进而导致线粒体损伤[21]。据报道,线粒体损伤后与核苷酸结合并激活寡聚结构域样受体蛋白3 炎症小体[22]。活化的寡聚结构域样受体蛋白3 炎症小体通过半胱氨酸蛋白酶1 对细胞中死亡调节蛋白进行剪切,释放白细胞介素-1β 和白细胞介素-18 等炎症因子,引起细胞凋亡。此外,Kobayashi等[23]发现,通过对线粒体通透性转换孔的抑制,可降低ROS 和其他自由基的产生,从而对SAE 中线粒体和细胞功能起到保护作用。因此,维持线粒体的分裂/融合稳态和线粒体膜完整性是防治SAE 的有效途径。
1.4 神经递质失衡
神经递质的变化与SAE 患者谵妄密切相关[24]。脓毒症中神经递质失衡导致小胶质细胞和脑内微环境失衡[25]。谷氨酸作为主要的兴奋性神经递质,可使N-甲基-D-天冬氨酸受体过度活化,进而导致神经损伤[26]。Wu 等[27]发现,LPS 可升高小鼠小胶质细胞内Ca2+,引起细胞外调节蛋白激酶及核因子κB 磷酸化,促进小胶质细胞激活及炎症因子释放,然而N-甲基-D-天冬氨酸受体抑制剂可抑制这些反应。神经递质与小胶质细胞具有相互调节作用,稳态小胶质细胞可释放中等水平的谷氨酸,但激活后的小胶质细胞会释放毒性剂量的谷氨酸,因此谷氨酸可能对小胶质细胞激活有正反馈作用[25]。此外,乙酰胆碱、γ-氨基丁酸、多巴胺、去甲肾上腺素、5-羟色胺等神经递质在SAE 的发生和发展中也起到了重要的作用[24]。
1.5 神经细胞凋亡
凋亡作为一种细胞的死亡调控方式,在SAE 的发生、发展中扮演着重要角色[21-22]。细胞内稳态被破坏,如感染、缺氧或氧化应激等可导致蛋白质错误折叠数目增加,从而触发内质网应激(endoplasmic reticulum stress,ERS)。ERS 是维持细胞内蛋白质的稳态的重要机制[28]。然而,过度ERS 与脓毒症所致细胞损伤关系密切,Li 等[29]发现,在SAE 的细胞模型中,ERS标志物(GRP78)的过表达以时间依赖性的方式使细胞活力显著降低并导致细胞凋亡。线粒体促凋亡分子丝氨酸蛋白酶在半胱天冬蛋白酶依赖性凋亡中起重要作用,在凋亡损伤之后,从线粒体移位到胞质,介导了X 连锁凋亡蛋白抑制剂的降解,阻碍了半胱天冬蛋白酶-9 的降解,进而引起半胱天冬蛋白酶-3 的激活,导致细胞DNA 损伤[30-31]。此外,Gao 等[32]发现,通过对IL-17RA/Akt/ERK1/2 信号通路的抑制,可以缓解氧化应激和神经元凋亡,从而改善CLP 诱导的SAE 大鼠的认知功能障碍。在SAE 病理过程中,PERK/STING/RIPK3 信号通路介导神经细胞坏死性凋亡,进而导致认知损害[33]。在CLP 小鼠模型中,中性粒细胞及中性粒细胞胞外杀菌网络可导BBB 破坏、神经元凋亡及小胶质细胞活化,导致认知功能损害[34]。综上,SAE 所致细胞凋亡受多种因素影响,但具体机制仍不清楚,上述发现可为SAE 的进一步预防和治疗方案的探究提供新的思路。
2 小结与展望
SAE 严重影响患者生活及生存,其发病机制复杂且尚未完全阐明。近年来,SAE 的机制得到了广泛而深入的研究。本文着重从BBB 破坏、小胶质细胞激活、线粒体功能障碍、神经递质失衡、细胞凋亡等多个角度概述了SAE 的发病机制。目前,虽然临床研究者在SAE 的发病机制研究中取得了一些进展,但有效的治疗手段仍然有限。未来的研究应集中于探索新的治疗方法与SAE 相关炎症反应、免疫反应及神经损伤之间的关系,以寻找对SAE 有效的药物作用靶点。此外,临床研究的设计和方法需要进一步改进,以提高研究的可靠性和可重复性,进而提高SAE 患者的生存率和生活质量。
利益冲突声明:本文所有作者均声明不存在利益冲突。
