牧草的重金属胁迫效应及修复机制研究进展
2024-03-09吴雅芬周柳婷王义祥黄小云黄秀声
吴雅芬,周柳婷,王义祥,罗 涛,黄小云,黄秀声
(1.福建农林大学资源与环境学院,福建 福州 350002;2.福建省农业科学院农业生态研究所,福建 福州350013;3.福建省丘陵地区循环农业工程技术研究中心,福建 福州 350013;4.福建省农业科学院土壤肥料研究所,福建 福州 350013)
随着我国工农业现代化进程不断加快,矿山开采、金属冶炼以及城市生活垃圾、污水排放等人为活动使土壤重金属污染日趋严重[1]。据《全国土壤污染状况调查公报》分析,2014年我国土壤镉(Cd)、汞(Hg)、砷(As)、铜(Cu)、铅(Pb)、铬(Cr)、锌(Zn)、镍(Ni)8 种重金属点位超标率分别为7.0%、1.6%、2.7%、2.1%、1.5%、1.1%、0.9%、4.8%[2]。重金属污染具有毒害性、持久性和生物富集性,可严重危害人体健康、引发环境污染和破坏生态。因此,对重金属污染土壤进行治理刻不容缓[3-4]。植物修复是土壤重金属修复的重要手段之一,主要利用植物及其根际圈微生物体系的吸收、挥发、转化和降解等作用清除环境中的污染物质,是一种绿色、经济、环保的重金属污染土壤修复技术[5-8]。植物修复通常利用超富集植物进行土壤修复和重金属提取,可以在维持超富集植物自身生长的情况下富集土壤中的重金属离子,其积累能力远超普通植物[9]。然而由于生长速率慢、生物量小、富集重金属单一等缺点,在实际应用中受限[10-11]。因此,经济和生态效益并存的牧草类植物成为植物修复领域的研究热点[12-13]。
牧草植物种类丰富,作为饲料可多次刈割增收产量,具有较高的经济生产价值[14]。与其他草本植物相比,牧草植物生长快速、生物量大,适应性、再生性强,可以同时吸收多种重金属元素,在生态恢复、水土保持等方面有重要作用[15-17]。此外,利用牧草进行植物修复还具有治理效果持久、操作管理简单、无二次污染等优点,已成为重金属污染土壤修复的主要方法之一[17]。因此,拟从牧草对重金属胁迫的响应特征和重金属富集效果及其强化措施进行综述,并对牧草修复技术的进一步优化提出展望,旨在为高效利用牧草植物治理重金属污染土壤提供参考。
1 牧草对重金属胁迫的响应
1.1 重金属对牧草生长发育的影响
重金属胁迫会造成植物生理活动异常,生长发育受阻,并存在低促高抑的剂量效应[18]。高浓度的重金属对植物的光合作用、呼吸作用、水分代谢、有机物质运输、胁迫耐受性以及矿物质营养等生理和生化过程产生破坏性影响,从而抑制植物的生存、生长和发育[19-23]。牧草在高浓度重金属环境下生长迟缓,根系数量减少、根长变短、发芽率降低、根部和茎叶干质量显著下降[24-27];除此之外,植株还会出现细胞膜透性增大、可溶性糖及脯氨酸含量增加[28-30]、体内抗氧化酶活性随重金属浓度增加而增强等生理应激现象[31]。然而,与高浓度的重金属胁迫效应相反,低浓度的重金属添加会促进牧草的生长发育[23]。大量研究表明,牧草在低含量重金属环境下,其发芽指数、根长、芽长、株高以及生物量出现不同幅度的提升[32-34]。俞花美等[34]研究发现,Cd含量为0.3 mg∕kg 时热研8 号坚尼草的株高和干质量分别增加了10.9%、27.4%。ZHANG 等[32]用不同含量Cd 处理杂交狼尾草发现,Cd 含量在0~15 mg∕kg 时植株的叶面积、叶长和叶宽分别增加了2.4%、15.8%,Cd含量在15~100 mg∕kg时叶面积下降了27.2%。鱼小军等[35]、MORADI 等[36]发现,Cu 质量浓度为300 mg∕L时红豆草发芽率增长了11%。
1.2 重金属对牧草光合作用的影响
重金属胁迫会加速植物叶绿素分解、破坏叶绿体结构,使反应所需的酶活性降低,从而导致净光合速率下降[37]。高浓度的重金属胁迫会诱导植物细胞发生氧化应激,生成大量活性氧促使叶绿素氧化降解,一些重金属离子如Hg、Cu、Pb、Ni、Cd 和Zn 进入细胞后会取代叶绿素中的镁(Mg),造成植物中毒[36,38-39]。有研究表明,过量的Cu、Pb 和Cd 不仅抑制叶绿素合成,还会使叶绿体膜降解直至消失[38]。GONZÁLEZ-MENDOZA 等[40]发现,过量的Cu 会干扰叶绿体和类囊体膜的组成,诱导植物细胞氧化应激,降低光合色素、电子载体的含量和电子转移速率,同时抑制光反应和暗反应的进行[41]。这与宋华伟等[42]、SONG 等[43]、PER 等[44]的研究结果一致。除此之外,植物的光合速率还与叶片气孔导度(Gs)、细胞间CO2浓度(Ci)、蒸腾速率(Tr)相关[45]。相关研究表明,低浓度的重金属会刺激Gs、Ci、Tr等相关参数增大,在短时间内促进牧草的光合作用,但随着重金属浓度的上升,代谢系统会因超出防御范围加速损伤,造成代谢失调,最终导致光合速率下降[45-46]。皱文桐等[47]对黑麦草、皇竹草和龙须草进行Pb、Cd、Zn、Cu 复合污染处理后发现,3种牧草叶片的光合参数均随污染程度的增加呈先增后减的变化趋势。孙启鑫[48]、闫志强等[49]也有类似的发现。
1.3 牧草对重金属的富集和体内分布
对重金属富集效果较强的牧草主要集中在禾本科和豆科,这些牧草的生物量极高,对Cd、Pb、Zn、Cu、Ni 和Cr 等重金属元素具有较强的耐性和积累能力,是理想的修复植物[50]。表1 介绍了部分牧草对污染土壤重金属的富集和体内重金属分布情况[17,51-65]。相比其他牧草品种,多花黑麦草和高羊茅对Cd 和Zn 以及披碱草对Cd 的富集系数均>1,高于其他牧草,可作为优势种类进一步研究[17,55-57]。然而牧草对重金属的转运系数均<1,表明重金属在牧草体内的转运能力不强。牧草根部对重金属的积累量高于茎叶,这与非超富集植物特征相符。
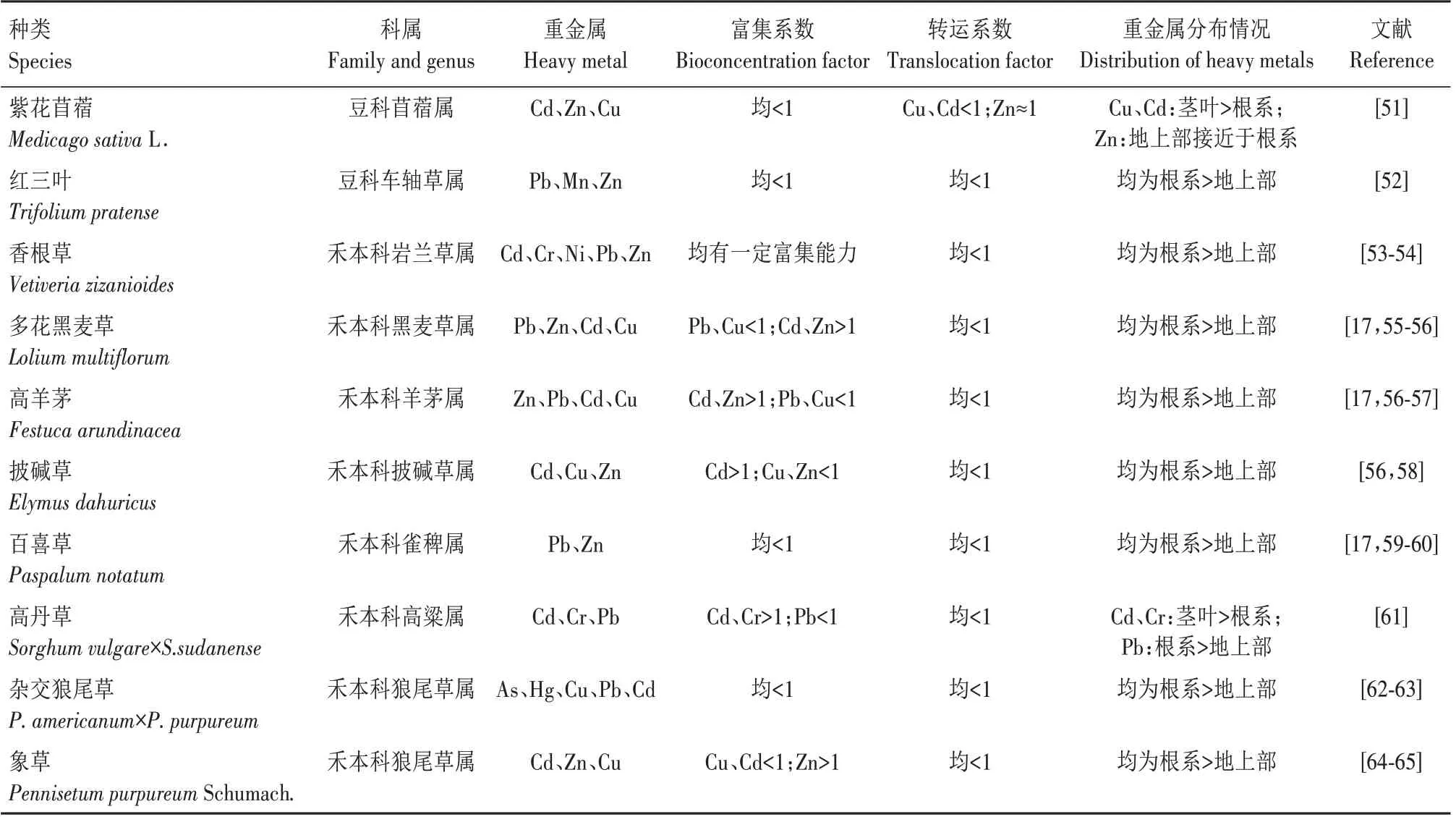
表1 牧草对污染土壤重金属的富集和分布情况Tab.1 Enrichment and distribution of heavy metals in contaminated soil by forage grass
2 牧草修复重金属污染土壤的机制
2.1 对土壤重金属进行活化和吸收
牧草在生长发育过程中会通过根系向根际环境释放有机化合物,这些有机化合物统称为根系分泌物。重金属胁迫会导致牧草根系有机酸分泌增加,有机酸可以转化土壤中有毒重金属的形态,如活化土壤中难溶态重金属化合物从而促进植物对重金属的吸收富集[66-68],还能与土壤重金属螯合形成溶解度低、迁移能力低的复合物,降低毒性[69-70]。有机酸的释放还会导致植株根际土壤pH 值、Eh(氧化还原电位)值下降,进一步促进重金属的溶解和移动。根际土壤中的重金属活化成为离子和金属螯合物形态后,会经质外体或共质体途径进入根细胞,或再通过根细胞转入木质部导管被植物地上部吸收[71]。例如植物根系分泌的低分子质量有机酸能与Cd结合形成低分子络合物,增加土壤Cd的溶解,而根系分泌物促进微生物的活动,导致土壤Eh值下降,形成硫化镉(CdS)降低Cd的活性[70]。ZENG 等[72]的研究表明,根系释放的氨基酸和有机酸会与根际土壤中的Pb、As、Zn、Cr 等重金属元素进行活化反应,使它们的生物有效性增加,便于植物的吸收、转运和富集。
2.2 液泡区隔化
重金属隔离在植物液泡中是牧草维持体内重金属稳态、减少其毒性作用的重要方式[71]。重金属进入细胞后有主动向液泡中运输的趋势。部分金属离子会与细胞质内的有机酸、氨基酸、多肽等物质形成结合态区隔在液泡中,从而阻止重金属以游离态的形式进入细胞器,有效缓解重金属毒害,维持牧草正常生长[73]。同时液泡中蛋白质、有机酸和有机碱均可与重金属结合,降低重金属毒性[71,73]。在紫花苜蓿的叶片亚细胞中,Cd主要以盐酸提取态形式分布于液泡,并在施加外援谷胱甘肽时含量占比升高,有效缓解Cd的毒害作用[74]。
2.3 重金属螯合
螯合作用是牧草对金属的解毒机制之一,重金属胁迫会诱使植株细胞螯合肽(PCs)的合成,同时促进PCs 和细胞中原有的金属硫蛋白(MT)与金属离子结合形成硫肽复合物,最终在液泡膜转运蛋白作用下转移至液泡中以降低细胞游离态金属离子浓度,从而提高牧草对金属的耐性[75-77]。何娇等[78]发现,在铯离子(Cs+)处理下,菊苣会通过合成较多的PCs 和MT 来缓解Cs 毒害。HELAOUI 等[79]的研究表明,随Ni 浓度升高,紫花苜蓿中参与PCs 合成的相关基因的表达将逐渐上调,随后PCs 与Ni 结合形成PCs-Ni复合物,实现对重金属的解毒作用。
2.4 与植物细胞壁结合
细胞壁是重金属进入细胞内部的第1 道屏障,细胞壁中的多糖分子和蛋白质分子可与金属离子结合而贮藏部分重金属,降低重金属对植株的毒害,牧草耐重金属胁迫能力与细胞壁和该重金属的结合能力有一定的相关性[80]。王爱云等[81]的研究发现,白花三叶草、高羊茅、紫花苜蓿在300 µg∕kg Cr处理下细胞壁的Cr 含量分别为46.25%、39.17%、32.56%,高于细胞质、线粒体和叶绿体中Cr含量,且3种牧草均表现为对Cr的耐受性较高。董名扬等[82]通过水培试验以不同浓度(0、200、800、2 000µmol∕L)硝酸镉溶液处理Monto(MT)和Veriveria nermorlis(VN)2 种生态型香根草7 d,香根草在200~2 000 µmol∕L Cd 处理下叶片中细胞壁的Cd 含量均随浓度增加而增加,其中VN 型香根草叶片中细胞壁组分Cd含量由18.87%增加到75.93%,MT型香根草叶片在2 000µmol∕L Cd 处理时细胞壁组分Cd 含量由40%增加到94.62%,表明植物细胞壁是实现重金属耐受与解毒机制的重要场所。
3 牧草对重金属污染土壤修复效果的影响因素
植物修复主要通过根系吸收土壤中的重金属并将其输送到茎、叶等地上器官,最终收割植物地上部以达到重金属去除的目的。在此过程中种植方式、土壤理化性质以及重金属间的相互作用是影响牧草对重金属污染土壤修复效果的关键因素。
3.1 种植方式
植物对重金属的富集、转运除与自身有关外,还与其种植方式密切相关[83-84]。与单作相比,间作能够增加牧草的生物量、抗氧化酶活性、根系分泌物数量、根际土壤养分含量、酶活性以及微生物数量,降低丙二醛、活性氧等物质含量,从而降低重金属的迫害,提高植物的修复效率[84-88]。由此可见,间作对牧草的种植培育具有显著的生态学意义[84]。王小慧等[89]研究发现,象草与苦楝间作时,象草植株的生物量可提高23.4%;象草与构树间作时,象草茎和叶中Cd 含量较单作分别增加71.6%、24.0%,富集和转运系数较单作分别增加29.3%、36.2%。孟楠等[70]将空心菜与不同草本植物间作发现,空心菜与高丹草、苏丹草、狼尾草间作能显著降低空心菜茎叶的Cd 含量,同时对污染土壤中Cd 具有较高的移除率。王梦珂[90]将黑麦草和苜蓿间作后发现,黑麦草和苜蓿生物量均显著提高,同时提高了黑麦草对Pb、Cd和Zn 的吸收,其中对Cd 和Zn 的吸收显著提高了40%、31%。
3.2 土壤理化性质
土壤理化性质影响重金属形态,从而影响植物对重金属的吸收与迁移,常见的与重金属生物有效性相关的土壤理化性质指标有pH 值、有机质含量、Eh 值等[84]。通常情况下,土壤pH 值与重金属有效性呈负相关关系,pH 值越低金属离子活性越强,越容易被植物吸收固定[91]。有机肥是常用的土壤改良剂,能改变土壤表面负电荷数量、分解小分子有机酸,与重金属形成稳定的络合物,降低重金属离子活性。伍港繁等[92]发现,有机肥调控能促进象草生长及其对Cd 的吸收累积,显著降低土壤有效态Cd含量和根际总Cd含量。土壤Eh值也是影响重金属活性的因素之一,其通过改变离子价态决定重金属的形态和迁移能力,进而影响重金属的富集[93]。一般情况下,土壤Eh 值与含水量呈负相关关系,土壤湿度越大还原性越强。荣伟英等[93]发现,Cr6+在土壤水位升高时会还原成Cr3+,被土壤胶体和其他组分吸附固定残留在土壤中[93-94]。
3.3 重金属交互作用
土壤中的重金属主要以复合形式存在,多种重金属之间会发生交互作用,从而影响金属离子的富集效果[95]。常见的交互作用类型分为拮抗作用和协同作用。有研究表明,重金属协同作用产生的毒害效应大于单个重金属污染效应之和,拮抗作用则与之相反,由此可见,重金属协同作用对生态环境的威胁更大[95]。目前常见的有Zn-Cd交互作用。大量野外调查及试验证明,缺Zn 条件下,植物极易吸收和积累土壤中的Cd,在缺Zn 的土壤中施加Zn 会明显降低植物对Cd 的吸收和积累,如施Zn 会抑制苜蓿对Cd的吸收[93,96]。
4 牧草修复重金属污染土壤的强化措施
4.1 农艺措施
间作、轮作和套作是我国传统的农田耕作模式,其主要通过利用生态位和生物多样性原理等提高作物对资源的有效利用、为共植作物增量提质,这同样适用于重金属污染土壤的治理修复[84-85]。大量试验表明,将生长习性相适的植物与牧草间作可提高其对土壤重金属的富集效率[84]。狼尾草与空心菜间作模式下,狼尾草可为空心菜提供遮阴环境,空心菜则能增加狼尾草对土壤重金属的移除率[70],类似的还有象草和构树间作[89]、黑麦草和苜蓿间作[90]等。套作和轮作模式亦可有效提高牧草对某些重金属的吸收。肖伟等[97]采用蒜苗与黑麦草套种,黑麦草对Pb 的吸收较单独种植显著增加了133.5%。陈璘涵等[98]研究甜菜-菊苣轮作模式对Cd的富集情况发现,甜菜和菊苣轮作其地上部对Cd的富集系数分别达到11.34、8.22,显著高于单作。
施肥是传统农艺管理方式之一,也是强化植物修复效果的有效手段之一。合理施肥可以通过提供给植物生长所需的营养元素及改善土壤质地达到高产、优质的目的[99]。同时,施肥还可以改变土壤的理化性质,如团聚体结构、pH 值、Eh 值等,进而改变重金属在土壤中的形态和有效性,直接或间接影响重金属在土壤中的迁移和分布[100]。秦秦等[101]研究发现,施用有机肥能促进土壤团聚体形成,增加大粒径团聚体的占比,使土壤对Cd2+的吸附量和缓冲能力显著提升。肖艳平[102]通过盆栽试验发现,蜈蚣草在施加氮、磷、钾肥后生物量显著提高,对As富集效率增强。
4.2 化学添加剂-牧草联合修复
常见的添加剂有乙二胺四乙酸(EDTA)、谷氨酸(GLDA)、氨基三乙酸(NTA)、柠檬酸(CA)等[103]。土壤施用添加剂能提高植物根际土壤酶活性、养分含量、金属有效性以及植物的生长速率和抗逆性,强化植物对重金属的吸收富集,是实现高效修复的新途径[103-105]。添加剂主要分为螯合剂、土壤酸碱调节剂、植物生长调节剂3类。其中,缓释螯合剂可以通过控制螯合剂释放速率使修复效果发挥到最大。植物生长调节剂如生长素(IAA)具有缓解重金属毒性、促进根系生长、增加生物量和提高重金属提取量的作用。此外,植物生长调节剂与螯合剂协同作用还可以强化植物修复效果。土壤酸碱调节剂如石灰、羟基磷灰石通过影响土壤pH 值来影响重金属有效性,从而提高植物的吸收效率。并且有研究表明,多种螯合剂联用施用的效果比单一螯合剂施用效果佳[106]。刘宝勇等[55]的研究表明,在高浓度Pb污染土壤中,黑麦草与EDTA、FeCl3联合施用后,相比于无添加剂的对照组,Pb的富集系数分别提高了2.6、2.1倍,转运系数分别提高了0.75、0.84。
4.3 微生物-牧草联合修复
微生物联合植物修复是目前常用的修复方式,成本低廉、绿色环保。该方式主要通过微生物、植株以及内生菌之间的协同作用提高植物抗性,减轻重金属毒害,以此强化植物的修复能力[107]。促生根际微生物(PGPR)是目前研究较多的一类。大量研究表明,PGPR 能够通过固氮、溶磷、分泌生长激素增加根、茎生物量,促进植物生长并改变植物对重金属的利用率[108-110],还能通过分泌有机酸来促进植物对重金属的吸收[111]。丛枝菌根真菌(AMF)-植物共生体系也是常见的微生物修复组合[112]。艾艳君[113]利用AMF-紫花苜蓿联合修复钒钛磁铁尾矿土壤的研究表明,该共生体系使土壤有效态Cu 和Pb的含量分别降低了21.60%、40.65%。
5 展望
综上所述,牧草具有很强的耐重金属性,能在较高浓度的重金属胁迫下生长,利用其高生物量吸附和去除土壤中的重金属,是一种绿色环保的治理手段。与其他植物相比,牧草修复投入成本低、重金属富集量高,在长成之后可供观赏或作为能源植物获取收益,具有良好的应用前景。目前,将牧草植物用于修复重金属污染土壤尚在探索阶段,仍存在许多不足。为了进一步优化牧草的修复效果,建议从以下几个方面加强研究。一是牧草品类繁多,不同种类的牧草特征特性不同,其对不同重金属的富集效果也存在差异。因此,应对牧草品种进行试验和筛选,尤其是重点对高产牧草品种进行选育,并创建数据库,便于调研参考。二是做好牧草修复重金属的后期材料处理工作。吸附重金属后重金属含量超标的牧草不宜再制成饲料饲喂动物,应对其做好分类、规范用途,不造成资源浪费和环境污染。三是探索重金属的回收工艺。植物将重金属从土壤提取到地上部之后如何处理才可以避免二次污染是当前值得研究的问题。如果能实现土壤重金属回收,达到循环利用变废为宝,在净化土壤污染的同时获取效益,将是很大进步。
