水体富营养化及物种入侵对星云湖食物网的影响
2024-03-08邓素炎温雯雯王明果黄林培陈子栋陈光杰赵帅营云南师范大学地理学部云南省高原地理过程与环境变化重点实验室云南昆明650500河北省丰宁满族自治县气象局河北承德06850云南省地矿测绘院有限公司云南昆明6507
邓素炎,郭 雯,温雯雯,王明果,黄林培*,陈子栋,陈光杰,赵帅营 (.云南师范大学地理学部,云南省高原地理过程与环境变化重点实验室,云南 昆明 650500;.河北省丰宁满族自治县气象局,河北 承德 06850;.云南省地矿测绘院有限公司,云南 昆明 6507)
水体富营养化及物种入侵已成为严重威胁我国湖泊生态安全的重要因素,导致湖泊生态系统退化,并可能对湖泊食物网结构与功能产生不可逆的影响[1–3].湖泊富营养化引起藻类大量繁殖,导致水体透明度和溶解氧降低,沉水植物消亡,使得食物网中的碳源构成发生改变,造成浮游碳源占比上升底栖碳源下降[4].另一方面,我国许多水系在不同程度上受到外来物种入侵的影响[5],外来物种排挤环境中的原生种,破坏当地生态平衡,严重影响本地群落的组成与结构,已成为除富营养化之外影响水生生态系统食物网结构的重要因素.然而,围绕水体富营养化与物种入侵双重作用下,对湖泊食物网结构影响与评价的研究工作仍较少,限制了多重环境胁迫下湖泊生态系统食物网结构与功能的认识.
碳、氮稳定同位素技术(δ13C 和δ15N)具有所需样品量少、能反映较长时间食物来源等特点,已成为分析消费者食性、构建食物网的有效手段[6-7].δ13C 值可用于识别消费者的碳源[8],而δ15N 值在食物网中相邻营养级的消费者间存在分馏效应,通常用于计算消费者的营养级[9-10].相关研究利用碳、氮稳定同位素技术探讨了营养生态位宽幅的有效性,以δ13C 值变幅(CR)指示食物来源和种类、栖息环境类型,δ15N 值变幅(NR)指示群落营养长度,并提出生态位校正标准椭圆面积(SEAc)等概念[11-12].在我国,不少学者使用碳氮稳定同位素构建白洋淀[4]、太湖[13]、保安湖[14]、兴凯湖[15]等水生生态系统的食物网结构特征.温周瑞等[13]研究结果表明浮游植物和底栖藻类为太湖贡湖食物网大多数生物有机体的主要碳源;张传鑫等[16]利用MixSIAR模型研究小清河口邻近海域底栖食物网结构,发现存在以浮游生物和沉积物有机质为主要能量来源的两条食物链.然而目前针对水生生态系统食物网研究多集中在东部湖泊及沿海区域,对西南高原湖泊的研究十分欠缺[17].
东部湖区多与江河连通,生物具有较强的扩散能力和适应能力,食物网稳定性强[18].相较于水文连通性较好的东部湖区,西南高原湖泊多为封闭-半封闭型湖泊,孕育了滇池金线鲃、抚仙湖鱇浪白鱼、大理弓鱼等众多特有土著鱼类,食物网易受人类活动和外来物种的干扰[19].近几十年来随着水体富营养化[20]和外来物种的引入[21],云南湖泊生态系统弹性降低,食物网脆弱,大量土著鱼类处于濒危处境[22-23],多样性下降.为探究富营养化和外来物种入侵对湖泊食物网的影响,以云南省星云湖为研究区域,研究湖泊初级生产者与消费者碳、氮稳定同位素特征,基于MixSIAR 模型量化消费者食物来源,对比分析消费者营养级计算的不同方法,构建湖泊食物网结构特征,旨在为星云湖的生态修复提供参考.
1 材料与方法
1.1 研究区域概况
星云湖(24°17′~24°24′N,102°45′~102°49′E)为滇中五大高原湖泊之一,湖面高程为1722m,平均水深5.3m,最大水深11.0m[24].湖泊面积为34.71km2,主要入湖河流有12 条,流域面积为378.0km2[25].星云湖年均气温15℃左右,受季风影响,年内降水分配不均,干湿季分明.过去几十年间,星云湖流域内生产生活用水增加、入湖营养盐负荷增多,导致星云湖自净能力下降和营养水平的上升,成为典型的藻型湖泊,显著影响了湖泊生态系统的能量流和物质流[4,26-27].而外来物种的引入则进一步威胁着星云湖土著鱼类的生存[22],引起食物网结构关系的改变.星云湖渔业历史悠久,资源丰富,大头鲤(Cyprinuspellegrini)、星云白鱼(Anabariliusandersoni)为星云湖特有的土著经济鱼类[28].太湖新银鱼(Neosalanxtaihuensis)、鲢(Hypophthalmichthysmolitrix)、鳙(Aristichthys nobilis)、间下鱵(Hyporhamphusintermedius)、鲫鱼(Carassiusauratus)、鲤鱼(Cyprinuscarpio)为星云湖外来鱼类[28],其中太湖新银鱼原产于长江中下游,自20 世纪80 年代引入星云湖后迅速成为优势鱼类种群,对土著鱼类形成严峻威胁.2009 年以来,政府取得农业部土著鱼增殖放流项目,通过人工繁殖、苗种培育等措施使大头鲤和星云白鱼有所增殖[29].
1.2 样品采集与处理
1.2.1 样品采集 在星云湖敞水区设置6 个采样点(图1),于2019 年3 月、6 月、9 月、12 月共四个季度使用采水器采集湖泊表层水质样品,共计24 个.水样置于1L 样品瓶低温避光保存,所有水样带回实验室用于水质指标测试分析.使用手持测深仪测量深度(Depth),用赛氏盘测定采样点的透明度(SD),水温、pH 值、溶解氧(DO)使用美国YSI 6600V2 多参数水质分析仪现场测定.根据《水和废水监测分析方法》,使用Smartchem200 全自动分析仪测定湖泊总氮(TN)、总磷(TP)浓度,使用丙酮萃取分光光度法测定叶绿素a(Chl.a)浓度.
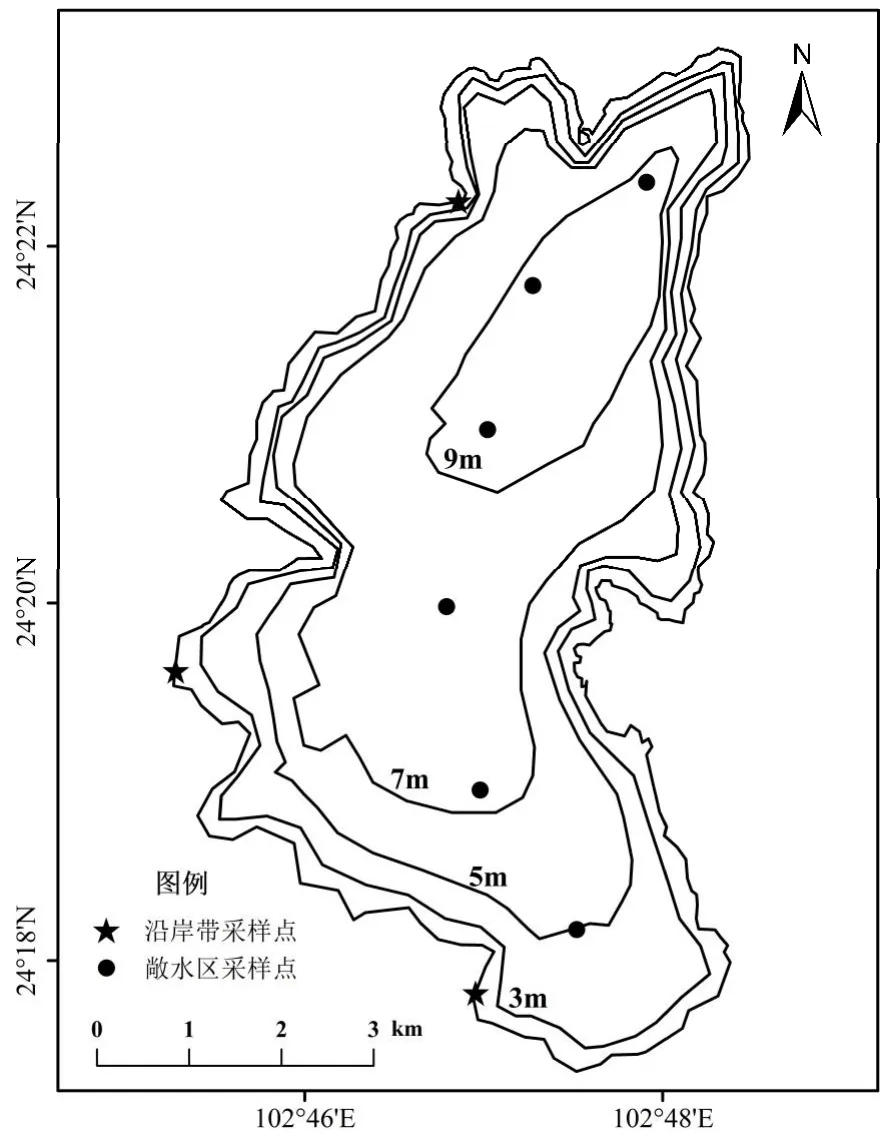
图1 研究区域地形图和星云湖等深线及采样点位置Fig.1 The Topographic map of study area and bathymetric map of Xingyun Lake and locations of the sampling sites
由于浮游生物生命周期短,其碳、氮稳定同位素受环境影响存在明显的季节性变化[30-32].为消除浮游生物同位素信号的季节差异,在采集水质样品的同时,使用22μm 和64μm 的浮游生物网在敞水区的6 个采样点采集浮游生物,分别获得浮游植物(22~64μm)、浮游动物(>64μm)样品各24 个,采集完成后将浮游生物样品放入125mL 采样瓶,使用4℃移动冰箱冷藏保存运回实验室.
2019 年9 月和2020 年1 月,在湖滨带随机采集了陆地有机物(枯落物和土壤)样品5 个.在湖泊沿岸带设置3 个采样点,利用抓斗式采草器采集沉水植物和底栖动物,获得6 种沉水植物样品16 个,4 种底栖动物样品13 个,共计29 个样品.采集到的沉水植物分别是菹草(Potamogetoncrispus)、金鱼藻(Ceratophyllumdemersum)、篦 齿 眼 子 菜(Potamogetonpectinatus)、光 叶 眼 子 菜(Potamogetonaceaelucens)、黑 藻 (Hydrilla verticillata)和穗状狐尾藻(Myriophyllumspicatum).底栖动物分别是扁卷螺(Hippeutiscantor)、耳萝卜螺(Radixauricularia)、沼虾(Macrobrachium)和龙虱(Dytiscidae).由于星云湖有严格的开渔期和禁渔期,且渔获物获取难度大,因此鱼类样品于开渔期向当地渔民购买,共计9 种鱼类,50 个样品,与2008 年调查的8 种鱼类[25]种类数量上相近,能较全面地代表星云湖鱼的种类信息.表1 记录了水生动物样品的名称、体长、体重、食性等信息,其中鲤鱼的体长相差较大,且随着鲤鱼个体的生长,其食性会发生转变[33],故在本研究中将鲤鱼细分为小鲤鱼和大鲤鱼.

表1 星云湖水生动物样品基本信息Table 1 Basic information of aquatic animal samples in Xingyun Lake
1.2.2 样品处理 用细毛刷轻轻刷去沉水植物表面附着的杂质,然后将沉水植物和浮游生物样品分别浸泡在装有1mol/L 盐酸的烧杯中,静置24h 以去除样品中的无机碳.水生植物使用去离子水反复清洗至中性,放入55℃的烘箱烘干,使用玛瑙研钵研磨至200 目以下均匀粉末.浮游生物样品使用离心机离心沉淀,反复清洗直至上清液呈中性,放入-50℃的冻干机冻干至恒重.使用十万分之一天平分别称取2.0~2.5mg 沉水植物样品和0.3~0.5mg 浮游生物样品,包入锡杯待测.相关研究表明,肌肉组织能够整合较长时间尺度的摄食信息,是分析动物食性、确定营养级、构建食物网的常用指标[41-42].因此使用手术刀、细胞夹解剖获取鱼类的背部肌肉、虾类的腹部肌肉以及螺类去壳取腹足肌肉用于碳、氮稳定同位素分析.使用冻干机干燥并称取1.2~1.5mg 肌肉样品,包入锡杯.利用Thermo Scientific 的MAT-253 气体同位素比质谱仪连用Flash EA元素分析仪测定δ13C和δ15N,标准物质为 USGS-40(δ13C 为(-26.389±0.042)‰;δ15N 为(4.5±0.1)‰).为确保数据的精准,在测样过程中每4 个样品插入1 个标准样品,所有分析测试在云南省高原地理过程与环境变化重点实验室完成.
1.3 研究方法与数据处理
碳、氮稳定同位素比值计算公式为:
天葬师此时身体尚未完全直起,眼见上方黑影一晃,一道黑芒挂着尖锐的破空声插下,他心知不妙,急忙向旁拧身躲避,却终究慢了一步。
式中:X表示13C 或15N,Rsample为样品中重同位素与轻同位素的比值;Rstandard为标准物质中重同位素与轻同位素比值.
利用δ15N 计算营养级(TL),计算公式为:
式中:δ15Nconsumer为待测样品的氮稳定同位素比值;δ15Nbaseline为基准生物的δ15N 比值;TEF 为相邻营养级间的传递富集系数,根据前人研究结果,本文使用3.40‰[43-44]作为TEF;λ为基准生物的营养级.
利用ArcGis 10.8 制作采样区域图,显著性差异采用Student’st-test 检验方法检验,使用R 软件运行MixSIAR 包(3.1.12)[45-46]计算碳源对消费者的相对贡献并进而计算营养级,使用SIBER 包(2.1.8)[47]构建鱼类营养生态位.数据的统计分析与制图在Excel、Origin 和R 软件中完成.
2 结果与分析
2.1 星云湖水化学特征
调研期间星云湖pH 值变化较小,为(9.16±0.27),水体年平均温度为(20.90±4.81)℃,透明度平均值为(0.63±0.15) m,水体溶解氧为(9.80±1.99) mg/L.总磷、总氮和叶绿素a 浓度季节变化较大,年平均值分别为(0.38±0.17) mg/L、(3.44±1.67) mg/L 和(85.66±58.79) μg/L.根据《Encyclopedia of Ecology》中综合营养评价法(TLI)[48]计算调研期间星云湖营养水平,其TLI 为6.86,属重度富营养水平.
2.2 水生生物C、N 稳定同位素组成特征
2.2.1 初级生产者δ13C、δ15N 特征 星云湖主要生物δ13C 和δ15N 特征如图2 所示,浮游植物δ13C 值季节变化显著,9 月最高,为(-21.32±0.37)‰,12 月最低,为(-23.51±0.08)‰(P<0.01),全年平均值为(-22.67±0.88)‰.6 种沉水植物δ13C 值较浮游植物的显著偏正,变化范围在-19.46‰~-10.38‰之间,平均值为(-13.45±2.93)‰(P<0.01).流域内枯落物和土壤有机质的δ13C 值差异较小,平均值为-25.84±2.50‰,为3 种类型初级生产者中最小(P<0.01).

图2 星云湖食物网主要生物δ13C 和δ15N 特征Fig.2 δ13C and δ15N characteristics of main organisms in the food web of Xingyun Lake
浮游植物δ15N 值的季节变化范围为11.30‰~14.88‰,平均值为(13.19±1.14)‰.6 种沉水植物δ15N值变化范围较大,为9.30‰~15.29‰,平均值与浮游植物的相近,为(13.13±2.56)‰(P>0.05).与湖泊内源相比,陆源的枯落物和土壤的δ15N 值相对较低,平均值为(7.46±1.04)‰(P<0.01).
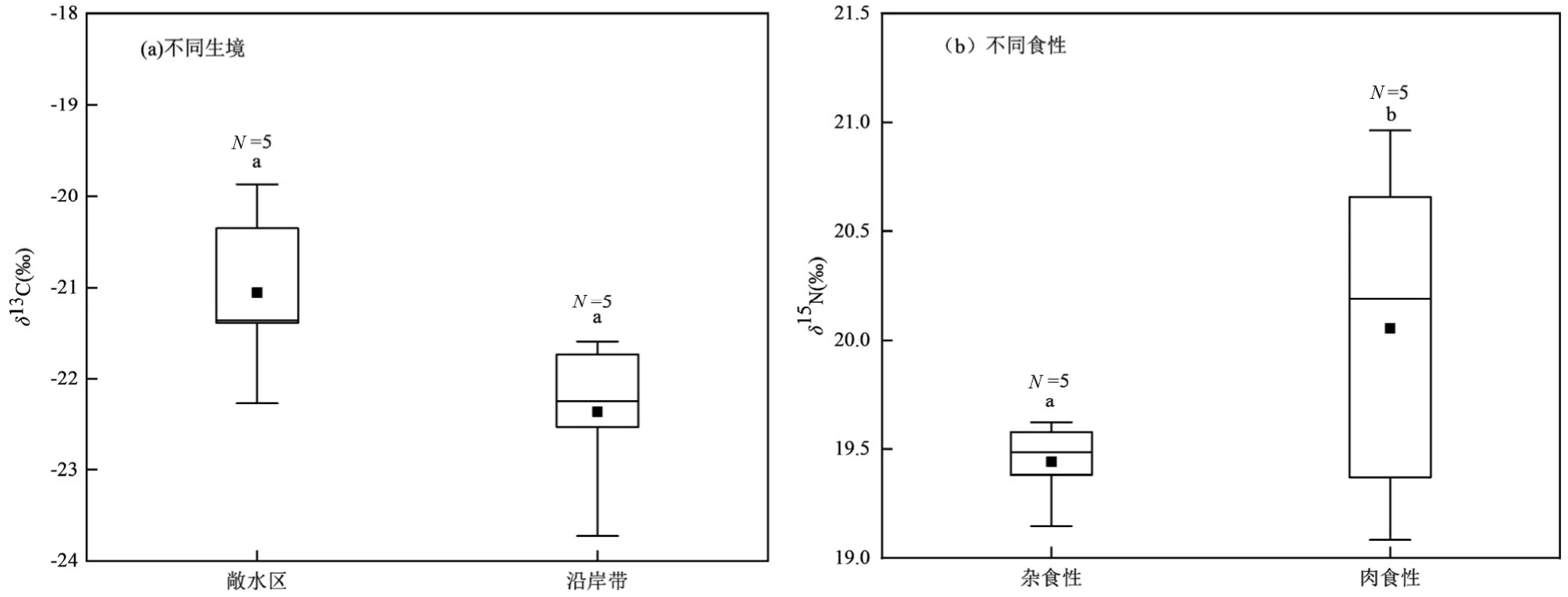
图3 不同生境、不同食性鱼类碳、氮稳定同位素分布特征Fig.3 Distribution characteristics of stable isotopes of carbon and nitrogen in fishes with different habitats and diets
消费者δ15N 值的范围在15.05‰~22.15‰之间,由大到小依次为:抚仙鲇>太湖新银鱼>间下鱵>沼虾>星云白鱼>鲫鱼>黄颡鱼>小鲤鱼>鳙>大鲤鱼>龙虱>鲢>耳萝卜螺>浮游动物>扁卷螺.浮游动物δ15N 值全年平均为(16.00±1.36)‰,较浮游植物高2.81‰.底栖动物δ15N 平均值为(17.30±2.11)‰,沼虾最高为(20.59±0.77)‰而扁卷螺最低为(15.14± 0.13)‰.鱼类δ15N 值的分布范围在15.77‰~22.15‰之间,平均值(19.27±1.59)‰.按照食性将星云湖鱼类分为杂食性鱼类和肉食性鱼类,杂食性鱼类δ15N 值为(18.49±1.35)‰显著低于肉食性鱼类(20.04± 1.37)‰(P=0.02).
2.3 消费者碳源及营养级
星云湖食物网初始碳源中浮游碳源(浮游植物)、底栖碳源(沉水植物)和陆源(枯落物、土壤有机质)之间的碳、氮同位素信号存在显著差异(P<0.05,P<0.05).利用MixSIAR 模型示踪3 种碳源对消费者的贡献,结果表明星云湖消费者以浮游碳源为主要食物来源,浮游碳源的贡献高达77.3%,而底栖碳源和陆源贡献分别为12.2%和10.5%(图4).
为确定计算营养级的基准物,本研究对比分析了3 种不同模式.模式A,浮游动物和敞水区鱼类以浮游植物δ15N 平均值作为营养级计算的基准;底栖动物和沿岸带鱼类以沉水植物δ15N 平均值作为营养级计算的基准,λ=1.模式B,根据MixSIAR 计算的3 种碳源对消费者贡献率,按加权平均获得δ15N值作为各消费者营养级计算的基准,λ=1.模式C,以浮游动物δ15N 值作为敞水区鱼类、以δ15N 值最低的扁卷螺作为底栖动物和沿岸带鱼类的营养级计算基准,λ=2.
根据这3 种模式分别计算了不同模式下星云湖消费者的营养等级(图5).模式A 结果显示,星云湖消费者营养级为1.59~3.57,浮游动物的营养等级为(1.83±0.40),底栖动物营养级为1.59~3.19,鱼类的营养级整体较高,为2.02~3.57.模式B 中消费者营养级为1.99~3.73,浮游动物的营养级为(2.39±0.40),底栖动物和鱼类的营养级分别为1.99~3.33 和2.34~3.73,消费者营养级总体略高于模式A.模式C 计算的消费者营养级最高,分布范围2~3.97,底栖动物营养级在2~3.60 之间,鱼类营养级为2.19~3.97,食物链长度接近4.
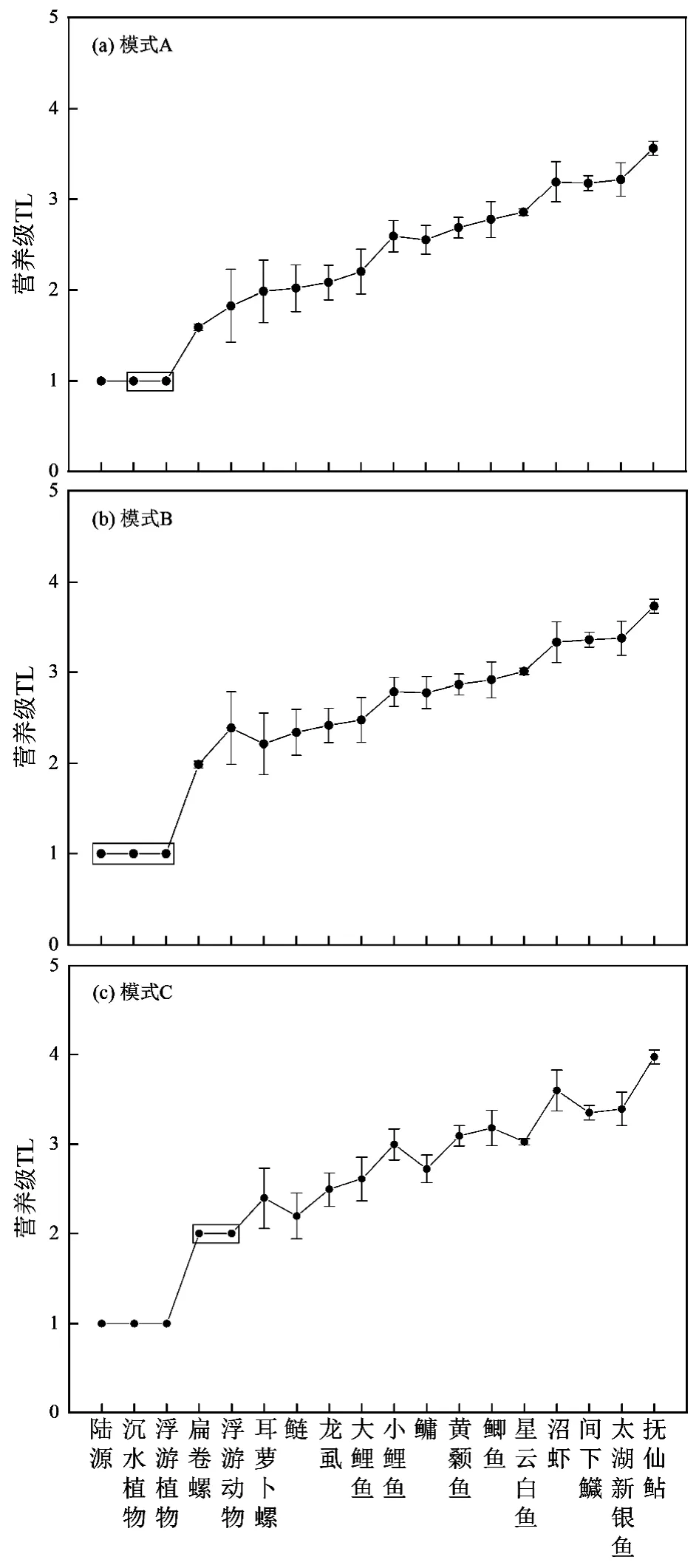
图5 基于3 种模式计算的星云湖消费者营养级Fig.5 Consumer nutrition levels calculated in terms of 3models in Xingyun Lake
2.4 星云湖食物网结构特征
在R 语言中使用SIBER 工具包构建星云湖杂食性和肉食性鱼类的营养生态位,结果表明星云湖杂食性和肉食性鱼类种群的营养生态位存在部分重叠(图6(a)).杂食性鱼类的CR 值为3.86,比肉食性鱼类的CR 值(2.69)高1.17.而杂食性鱼类NR 值则比肉食性鱼类低0.44,分别为3.85 和4.29.杂食性鱼类的核心生态位面积(SEAc)为3.79,大于肉食性鱼类(2.46,图6(b)).
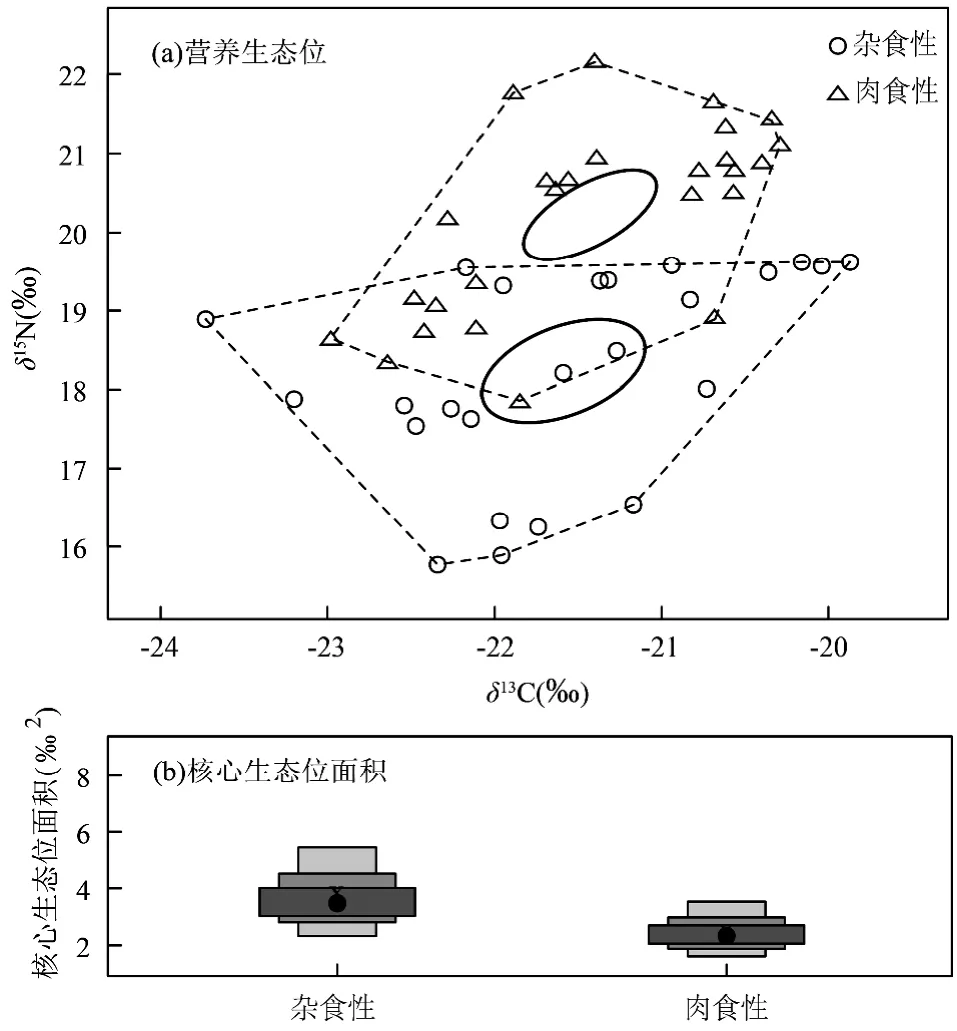
图6 杂食性与肉食性鱼类营养生态位与核心生态位面积Fig.6 Nutrient niche and core niche area of omnivorous and carnivorous fishes
为探讨鱼类个体大小与营养级之间的关系,将星云湖鱼类的体长分别与δ15N 值、营养级(模式B结果)进行相关性分析(图7),结果显示星云湖大个体鱼类反而具有较低的δ15N 值与营养级特征,即δ15N值和营养级随着鱼类体长的增加而减少,如太湖新银鱼体长最小却位于较高的营养级位置,但相关性并不显著(P>0.05),异常点为土著鱼类抚仙鲇,抚仙鲇个体较大,营养级最高.剔除抚仙鲇后,相关系数分别为-0.80(P<0.01)和-0.81(P<0.01).
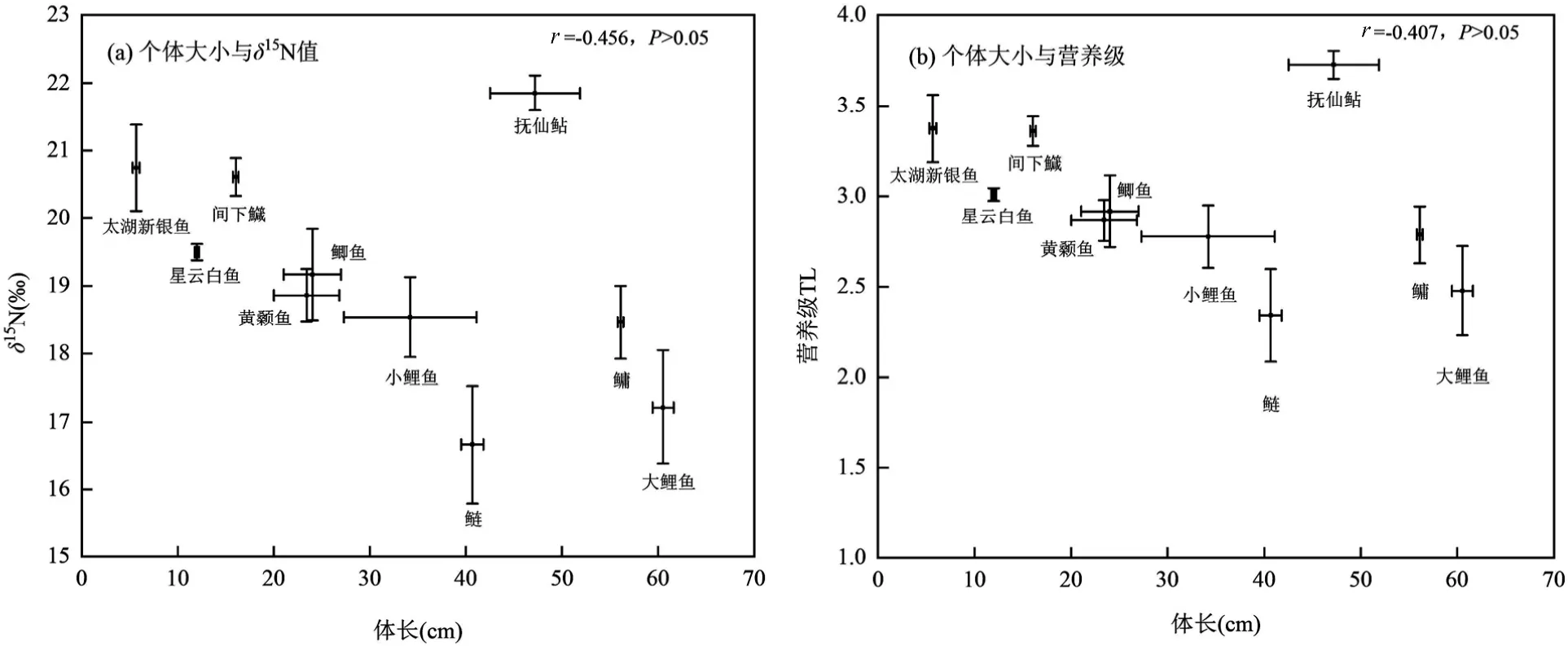
图7 鱼类个体大小与δ15N 值、营养级的相关性Fig.7 Correlations of fish size with δ15N value and trophic level
不同体长鲤鱼的δ15N 值和营养级差异显著,小鲤鱼δ15N 值(18.54‰)高于大鲤鱼(17.21‰,P<0.05).小鲤鱼在模式B 中的营养级为2.78,而大鲤鱼仅为2.48.
3 讨论
3.1 食物网初始碳源贡献及能量传输路径识别
不同营养水平湖泊初始碳源对食物网的贡献存在明显差异,贫营养湖泊以底栖初级生产力为主,随着营养水平的提升,则逐渐向浮游初级生产力转变[49-50].星云湖属重度富营养水平,MixSIAR 模型结果显示,浮游碳源对星云湖消费者的贡献率为77.3%,而陆源和底栖碳源对消费者的贡献相对较低.水体富营养化会促进藻类大量繁殖,遮蔽水下光照,导致沉水植物的衰退甚至逐渐消亡[51],从而削弱底栖碳源对食物网的贡献.调研期间星云湖透明度为0.63m,叶绿素a 浓度为85.66μg/L,属于典型的藻型湖泊.调查表明,星云湖沉水植物面积仅为0.6km2,群落较为单一,覆盖度约为1.8%[52-53].MixSIAR 模型结果与星云湖高营养水平和低沉水植物覆盖度现状一致,指示浮游碳源对星云湖等藻型湖泊食物网具有重要贡献.
沉水植物群落的衰退不仅使星云湖食物网中浮游碳源占比升高,能量传输路径也发生转变.湖泊消费者利用的初始碳源构成及能量传输路径往往与其生境密切相关,底栖动物及生活于沿岸带鱼类多利用底栖碳源(沉水植物、底栖藻类),能量传输以底栖路径为主[13,54-55].而星云湖食物网中,除扁卷螺外,其余所有消费者的δ13C 值均靠近浮游植物端元(图2).且沿岸带和敞水区不同生境鱼类的δ13C 值并不存在显著性差异(图3),指示在富营养化湖泊中,不同生境鱼类及底栖动物利用的初始碳源具有相似性,均以浮游碳源为主,能量传输路径则依赖较单一的浮游传输路径.
3.2 消费者营养级计算的最优模式选择
不同的食物网其基准物的氮同位素值差异较大[56],基准物的变化会通过食物链的传递作用最终影响食物网各生物类群的营养级,进而影响对食物网结构与功能的判断[57],因此合适的基准物是确定食物网中消费者营养级的关键.陈俊伊等[14]对保安湖食物网的研究发现在计算湖泊消费者营养级时,不能以单一的基准物来确定不同食性的鱼类营养级.星云湖初级生产者的δ15N 值存在显著差异,陆源较内源低5.70‰(图2).在模式A 中,尽管考虑到能量传输在浮游和底栖路径上的差异,区分了敞水区消费者与底栖、沿岸带消费者营养级的计算基准,但忽视了陆源对消费者的贡献.陆源有机质δ15N 值较低[58],对于陆源贡献相对较大的消费者,模式A 计算的营养级偏低.如基于模式A 计算的扁卷螺营养级仅为1.59,食物链长度3.57 也是3 种模式中最短的.
目前国内利用碳、氮同位素技术构建水域生态系统多以蚌类(或浮游动物)和螺类分别作为浮游食性和底栖食性消费者营养级计算的基准物[57].模式C以初级消费者浮游动物和扁卷螺δ15N值作为基准,综合考虑了各碳源对食物网中消费者的贡献,较模式A 更为优化.但是不同消费者所利用的初始碳源比例并不一致,会导致营养级计算结果失真.MixSIAR 模型结果显示,星云湖浮游动物和扁卷螺的陆源贡献分别为33.3%和24.1%,在所有消费者中最高(图4).以较高陆源占比的消费者作为基准物,会造成营养级计算基准的δ15N 值降低,从而导致其他消费者营养级被整体高估.
模式B以各消费者所利用初始碳源贡献权重计算的δ15N 值作为基准,即每一消费者都有不同的基准,这种计算模式虽然繁琐,但充分考虑3 种碳源对消费者的贡献及不同碳源间δ15N 值的差异性,更能精准获得各消费者营养级.基于模式B 的结果与消费者食性(表1)进行对比发现,消费者营养级与其食性相吻合,如初级消费者扁卷螺营养级为1.99.模式B中浮游动物营养级为2.39,这是由于浮游动物样品粒径>64μm,包含了不同大小个体的浮游动物,而某些大个体浮游动物(如剑水蚤)是捕食性浮游动物[59],导致浮游动物营养级略高于2.因此,若以浮游动物作为基准(λ=2)需剔除捕食性物种,否则可能会导致营养级计算结果偏离真实.上述分析表明,模式B 的结果能够真实地反映星云湖各消费者食性,可用于湖泊消费者营养级计算,有助于为其他生态系统消费者营养级计算提供模式参考.
3.3 星云湖食物网结构特征
基于模式B 计算结果,星云湖消费者营养等级在1.99~3.73 之间,鱼类的营养级为2.34~3.73,其中营养级超过3 的鱼类有星云白鱼(3.01)、间下鱵(3.36)、太湖新银鱼(3.37)和顶级捕食者抚仙鲇(3.73).抚仙鲇是滇中湖泊土著鱼类,个体较大,生性凶猛,常以小型鱼类为食,肉食性的鱇浪白鱼和鰕虎鱼都可以成为抚仙鲇的食物,因此抚仙鲇营养级较高.
太湖新银鱼原产于长江中下游浅水湖泊,于1982 年引入星云湖,虽然平均体长仅有5.67cm,但其具有能快速适应当地环境形成自然繁殖群体的特点,且在入侵地比原产地具有更高的遗传多样性[60],使其快速入侵并成为优势种.云南高原湖泊多为封闭-半封闭型湖泊[61],流域生态脆弱,在长期的进化过程中,云南湖泊土著鱼类缺少与其他相似物种的竞争能力,当有生态位重叠的物种入侵时,土著鱼类容易失去生存能力[62].仅3a 时间,太湖新银鱼便迅速成为星云湖的优势种[63],营养级高于星云白鱼而仅次于抚仙鲇.星云白鱼为星云湖特色本土鱼种,其δ13C 值为(-20.61±0.69)‰与太湖新银鱼的(-20.75±0.63)‰极为相近,表明两者的食物来源构成上相似(图4),都以枝角类、桡足类为主要食物,存在食物竞争关系.在生活习性上,太湖新银鱼与星云白鱼生活在水体的中上层[36,38],在生存空间和摄食上均存在竞争.因此,太湖新银鱼的入侵在较大程度上挤占了星云白鱼的生态位,导致星云白鱼资源的衰退.渔获数据表明,星云白鱼曾是星云湖重要经济鱼类,60 年代初,年产2.5~3万kg,占鱼产量的6%~8%[36],最高年份产量占20%[28].1985~1987 年,太湖新银鱼产量呈倍数增长,分别为 7.8×104kg、20×104kg 和 40×104kg[25].与太湖新银鱼的快速增长趋势相反,星云白鱼数量在20 世纪80 年代末锐减,到90 年代后期,已经到了濒临灭绝的地步,现如今星云白鱼仍需要依靠放流来维持种群数量.星云白鱼与太湖新银鱼数量上的相互消长现象与碳氮稳定同位素组成揭示的生态位挤占结果吻合,表明太湖新银鱼等外来物种的入侵,破坏了星云湖固有的生态平衡状态,导致土著鱼类种群数量衰减甚至物种灭亡,进而使星云湖生物多样性减少,生态系统功能弱化.
星云湖原有15 种土著鱼类,本次调研除了顶级捕食者抚仙鲇外,只捕获到杂食性的星云白鱼.营养生态位结构显示,不同食性鱼类之间的生态位出现重叠现象(图6),表明杂食性和肉食性鱼类之间存在一定的食物竞争关系.杂食性鱼类的基础食物源(CR)比肉食性鱼类的跨度大,而营养长度(NR)比肉食性鱼类的低,指示杂食性鱼类具有比肉食性鱼类更广的食物来源以及较低的营养级.杂食性和肉食性鱼类均会摄食小鱼、小虾及浮游动物等,因而生态位部分重叠.除此之外杂食性鱼类还摄食浮游植物、水草等初级生产者,使得杂食性鱼类营养级总体较肉食性鱼类低,但食物来源较肉食性鱼类更丰富.此外,杂食性鱼类占据的核心生态位面积(3.79)大于肉食性鱼类(2.46),核心生态位相互分离,表明星云湖鱼类食物来源丰富,且杂食性鱼类拥有较肉食性鱼类更高的适应性,在食物短缺时,能够通过更换主要食物而得以存活.杂食性星云白鱼与肉食性太湖新银鱼主要以枝角类和桡足类为食,星云白鱼竞争不过太湖新银鱼,但杂食性的星云白鱼食物来源更广、选择性更大[36,38],可以通过改变食物来源结构,如摄食水生昆虫、藻类、水草等(更低的δ15N 值),而实现与太湖新银鱼共存,避免了物种灭绝,体现了杂食性鱼类的适应性.
星云湖鲤鱼个体差异较大,碳氮同位素结果显示,大、小鲤鱼的δ13C 值不存在显著性差异,而δ15N值差异显著,指示小鲤鱼较大鲤鱼具有更高的营养级.随着鱼类个体发育,其体长增大,口器日趋完善,摄食的食物大小、种类、比例等发生一定的变化[64],进而影响鱼类的同位素组成.MixSIAR 模型结果显示,大鲤鱼利用的初始碳源中浮游碳源较小鲤鱼下降约10%,而底栖碳源和陆源略有上升,这与大鲤鱼食物构成中摄入更多的水生植物、螺蛳和水生昆虫幼虫等底栖物种有关.虽然鲤鱼属于杂食性,但在幼鱼阶段更喜吃动物性食物,如轮虫、甲壳类及小型无脊椎动物等,使得小鲤鱼营养级(2.78)略高于大鲤鱼(2.48).鲤鱼不同发育阶段食物来源构成差异及营养级变动能够很好地反映在碳氮同位素组成中,为精细研究动物食性转变提供帮助.
前人研究表明,在不同物种间,鱼类体长增加往往伴随着高δ15N 值和高营养级[65],大个体鱼类多位于食物链的上层.然而星云湖食物网结构并不存在个体越大营养级升高的特征,相反,鱼类营养级具有随着体长的增加而降低的趋势.星云湖为重度富营养化湖泊,大量藻类的分泌液或死亡后产生的黏液会附着在鱼类腮上,且藻源性有机质分解耗氧等会导致原有本土鱼类窒息死亡,降低物种多样性,弱化鱼类个体与营养级关系.另一方面,外来物种的引入也破坏食物网结构,如小个体高营养级的太湖新银鱼、间下鱵以及大个体低营养级的鳙、鲢、鲤.因此,在富营养化和物种入侵的双重影响下,星云湖食物网结构发生改变,形成小个体鱼类营养级反而高的异常现象.
4 结论
4.1 富营养湖泊星云湖食物网中,不同生境鱼类及底栖动物利用的初始碳源具有相似性,均以浮游碳源为主,平均贡献率为77.3%,能量传输路径依赖较单一的浮游传输路径.
4.2 基于碳源对消费者贡献率计算的δ15N 值作为各消费者营养级计算基准(模式B),最贴合星云湖消费者营养级结果.
4.3 星云湖食物链长度3.73,顶级掠食者为抚仙鲇,入侵物种太湖新银鱼(3.37)次之.太湖新银鱼等入侵物种与土著鱼类在生存空间和摄食上的竞争,是导致星云湖土著鱼类如星云白鱼种群数量锐减的重要原因,造成星云湖食物网的生物多样性减少,生态系统功能弱化.
4.4 富营养化和物种入侵已显著改变星云湖食物网结构特征,形成小个体鱼类营养级反而高的异常现象.
致谢:本实验的现场采样工作由云南省高原地理过程与环境变化重点实验室各位老师和同学协助完成,在此表示感谢.
