华南中小型闸控入海河流DOM 特征及藻源影响
2024-03-08范中亚罗千里贾政博赵长进曾凡棠生态环境部华南环境科学研究所广东省水与大气污染防治重点实验室广东广州510655
李 丹,范中亚,罗千里,贾政博,陈 钢,赵长进,曾凡棠,黄 露 (生态环境部华南环境科学研究所,广东省水与大气污染防治重点实验室,广东 广州 510655)
溶解有机质(DOM)是地球上最主要的活性碳库之一,是全球碳循环的重要组成部分[1].作为能被微生物直接利用的碳源,DOM 对维持河流-海湾生态系统功能和生物地球化学循环具有重要意义.溶解有机碳(DOC)是DOM 的主要组分,常被用来量化DOM[2].上世纪80 年代,全球河流DOC 平均浓度约为5.75mg/L[3].然而,随着气候变化和人为干扰影响的加剧,河流中DOM 浓度、来源构成及组分特征发生显著变化[4],导致全球DOC 浓度和入海通量的估算存在很大的不确定性[5].国内对河流碳循环的研究主要是为了评估流域风化在区域碳循环中的贡献[6-7]或者量化大型河流有机碳的入海通量[8-9],对中小型入海河流DOM 特征及来源解析研究较少,相关研究的不足限制了对河流-海湾低氧现象及区域碳循环的认识.
近年来,人为干扰特别是富营养化和大规模拦河建坝/闸极大地改变了河流原有的生物地球化学特性及水体流动性,河流藻类异常增殖现象时常发生[10].藻类大量增殖加速无机营养元素向有机形态转化,产生的藻源DOC 甚至可能超过陆源DOC 输入量[11].研究表明,湖泊中藻类异常增殖时DOC 浓度与藻类生物量正线性相关[12],并且这种相关性的季节性变化可以表征藻类的生消过程[13].相关研究表明[14]藻类生长循环显著影响巢湖DOM 来源和性质,太湖DOM 及分子组成受藻类增殖过程控制[15].然而,藻类异常增殖对河流DOM 的浓度、来源构成和组分的影响仍缺乏系统全面的研究.
华南亚热带季风气候区分布着数百条中小型河流,因气候因素和人为干扰叠加导致藻类异常增殖现象频发[16].中小型河流富营养化不仅破坏河流生态系统健康,还直接限制流域城市生活生产活动和经济发展[17].如果不加强研究和管理,这些数量众多、水面分布广的中小型河流汇入海湾后可能会导致海湾发生赤潮、加速海湾低氧区域增加和海洋生态系统破坏[18].黄江河、东溪河和练江(三河)是华南典型的中小型闸控入海河流,水质现状分别是Ⅱ类、Ⅲ类和Ⅳ类水平[19-20],呈不同程度藻类增殖特征.本研究以三河为研究对象,于2022 年开展为期1 年的采样监测,分析河流DOM 浓度、来源构成与荧光组分的时空分布特征及其影响因子,以期为华南中小型闸控入海河流碳循环及河流-海湾低氧现象研究提供基础.
1 材料与方法
1.1 研究区域概况
黄江河、东溪河和练江(三河)位于广东省东部,是典型的中小型闸控入海河流.其中,黄江河发源于广东莲花山脉腊烛山,流域面积1121km2,常住人口约76.2万人,流域内土地利用类型主要为林地和耕地,占比74.0%;东溪河为黄江河入海通道的分支,河道起点位于黄江河汇合处,终点位于漯河汇合处,流域面积475.5km2,常住人口约8.2 万人,流域内土地利用类型主要为耕地、林地和果园,占比80.1%.练江流域发源于普宁市,流域面积1353km2,常住人口约380 万人.三河均处在南亚热带海洋性气候区,风力强劲,阳光充足,降雨主要集中在每年5~9 月份,为丰水期,其他月份为枯水期,2022 年三河丰水期月均降雨量分别为573, 427, 243mm, 枯水期月均降雨量分别为112, 96, 87mm;全年平均温度均超过20℃,且夏季温高雨多,冬季稍冷雨少.根据地理位置和流域土地利用类型情况,于黄江河(HJ1~HJ4)、东溪河(DX1~DX4)和练江(LJ1~LJ5)沿程共布设13 个采样点位,点位分布如图1 所示.
图1 黄江河、东溪河和练江采样点位分布Fig.1 Sampling sites of the Huangjiang, Dongxi, and Lianjiang Rivers at the Eastern Guangdong Province
1.2 样品采集与处理
于2022 年1~12 月对黄江河、东溪河和练江进行每月2 次沿程采样.现场采样时,利用便携式多参数水质分析仪(YSI, ProDSS)原位测定表层水体水温(T)、pH 值、盐度(SAL)、溶解氧(DO).采集后的水样置于冷藏箱中,遮光低温(4℃)保存并于当天运回实验室.TOC、DOC 浓度测定分别使用原水样和过滤后的水样(孔径为 0.2µm 的聚碳酸酯膜,Millipore).降雨量(Precipitation)数据由各流域所在县区气象局提供.闸控调度数据由各流域所在县区生态环境部门提供.
1.3 样品分析与数据处理
1.3.1 基本化学参数分析 水样预处理及基本参数分析均按照《地表水和污水监测技术规范》(HJ/T 91-2002)[21].其中,总氮(TN)浓度采用过硫酸钾消解-紫外分光光度法测定(HJ 636—2012)[22];总磷(TP)浓度采用过硫酸钾消解钼酸铵分光光度法测定(GB 11893-89)[23];高锰酸盐指数(CODMn)采用酸性高锰酸钾法测定(GB 11892-89)[24];叶绿素a(Chl a)浓度采用分光光度法测定(HJ 897-2017)[25].
1.3.2 有机碳浓度分析 使用高温催化氧化总有机碳分析仪(日本岛津,Shimadzu TOC-L CPH/CPN)测定总有机碳和DOC 的浓度.每测12 个样品后用Milli-Q 水和标准品(1.0mg/L 邻苯二甲酸氢钾为DOC)进行数据质量检查.在样品运行过程中,参考样品测量的精度优于10%,相对标准偏差在5%以内.
1.3.3 三维荧光光谱分析 使用荧光光谱分析仪(日立公司,F-7000)测定DOM 的荧光特性,测定使用1cm 四通石英比色皿.主要测定参数如下:激发波长Ex为200~450nm,扫描间隔为5nm;发射波长Em为220~550nm,响应时间0.02s,扫描速度为12000nm/min;每一批样品测定前均采用Milli-Q 超纯水作为空白测定其三维荧光光谱,并以此检查仪器稳定性.所有样品的三维荧光光谱均需进行空白扣除以减少样品中非目标信号的干扰.
三维荧光光谱数据分析在Matlab 软件(美国Mathworks,R2019b)环境下进行操作,采用 DOM Fluor 荧光工具包进行辅助分析.将样品扣除空白之后,以超纯水在350nm 激发波长(发射波长370~430nm)下水的拉曼峰的积分强度进行荧光强度标准化(Raman Unit R.nm-1),并进行瑞利散射的校正.使用DOMFlour 工具包对已处理好的荧光数据进行平行因子分析,并基于半分裂分析和残差分析进行组分识别和确定[26].
1.3.4 吸光特性分析 使用紫外-可见分光光度计(UV-1800,Shimadzu)对过滤后的样品进行吸光特征分析.取3mL 样品在270,350 和700nm 波长处进行吸光度测量.在每个样品批次之前,使用Milli-Q 水进行空白测量,并用于校正样品的光谱.
卡特模型[27](Carter Model)是根据DOM 紫外光谱特性,构建的三组分模型,其中组分A 具有强吸光性、强芳香性和强疏水性;组分B 有弱吸光性和强亲水性;组分C 为藻源,其特征是不吸光且浓度恒定较低.然而,后续研究表明卡特模型显著低估了富营养化水体中藻源DOC 浓度[28].因此, Adams 等[28]通过测算各组分的消光系数、水体DOC 浓度值及270和350nm 吸光度,利用修正后的卡特模型估算藻源DOC 对总DOC 的贡献(式(1)~(4)).
式中:EAB,λ为DOM 组分A、B 在波长λnm 下的消光系数,R为DOM 两个波长吸光度之比,Aλ为DOM在波长λnm 下的吸光度.组分A 中E270=69.3,E350=30.0,组分B 中E270=15.4,E350=12.0[28].
1.4 数据处理
用EXCEL2021、SPSS24.0 软件做数据整理及统计分析,数据均以(平均值±标准差)表示,采用ANONA 单因素方差分析沿程DOC、叶绿素a 和藻源DOC 浓度变化,满足方差齐性检验则用邦佛伦尼法,不满足方差齐性检验则用塔姆黑尼法分析显著相关性,采用非参数检验分析方法及Mann-Whitney U 检验法分析丰、枯水期三河叶绿素a 浓度和荧光强度占比变化;采用Spearman 相关性分析法分析DOC 浓度与环境因子相关性,用Origin 2021 作图.
2 结果与讨论
2.1 三河主要监测参数
根据2022年1~12月监测结果,黄江河TP、TN、CODMn和叶绿素a 浓度分别是(0.04 ± 0.01), (0.88 ±0.53), (2.8 ± 0.5) mg/L 和(20± 8) µg/L.东溪河TP、TN、CODMn和叶绿素a 浓度分别是(0.03 ± 0.01),(1.33 ± 0.17 ), (4.5 ± 0.8)mg/L 和(31 ± 13) µg/L.练江TP、TN、CODMn和叶绿素a 浓度分别是(0.22 ± 0.78),(4.96 ± 1.95), (4.6± 1.2) mg/L 和(82 ± 61) µg/L.根据我国《地表水环境质量标准》( GB 3838-2002),2022年黄江河水质类别为Ⅱ类,东溪河水质为Ⅲ类,练江水质为Ⅳ类,三河富营养化程度为练江>东溪河>黄江河(图2).
2.2 DOC 浓度时空变化特征
黄江河、东溪河和练江DOC 平均浓度分别是(2.6±0.9), (4.3±1.5)和(5.1±1.6) mg/L,分别占TOC浓度的78.6%、81.3%和67.2%,是三河TOC 主要存在形态.时间上,三河DOC 浓度均呈枯水期>丰水期特征(图3);空间上,黄江河和东溪河DOC 浓度总体呈沿程逐渐升高趋势,而练江DOC 平均浓度中上游(LJ1~LJ3)没有显著性变化,下游DOC 平均浓度呈显著升高趋势.
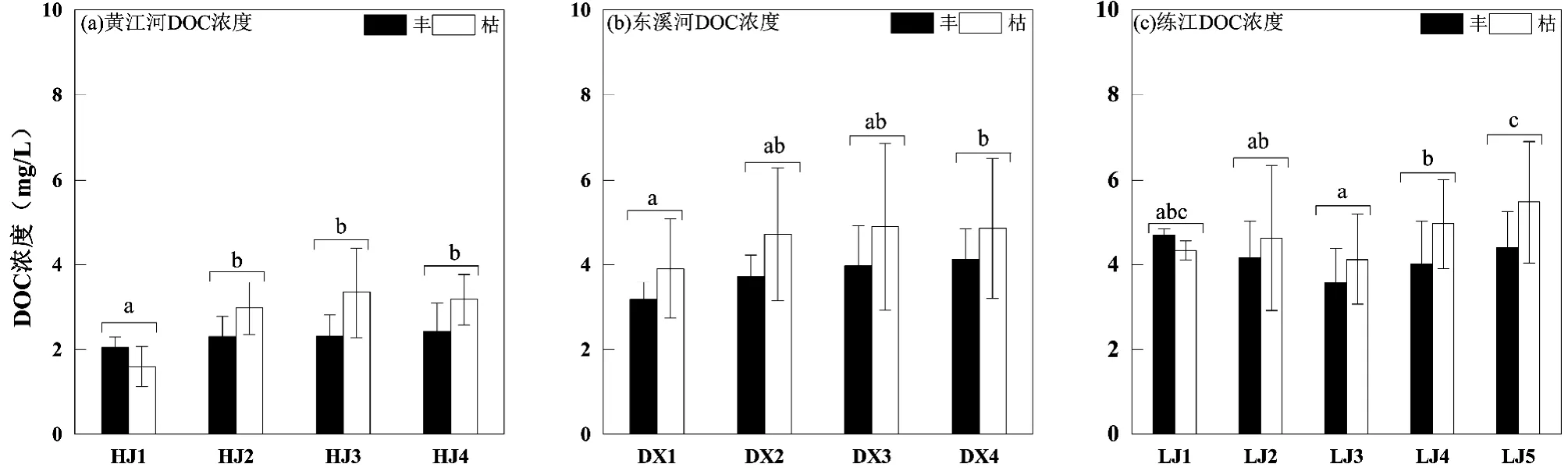
图3 黄江河、东溪河、练江DOC 浓度时空变化Fig.3 Spatial and temporal variations of DOC concentration in Huangjiang River, Dongxi River and Lianjiang River La
三河DOC 浓度显著低于世界河流平均浓度(表1),但是高于我国长江[29]、珠江[30]等大型流域以及华南地区相同气候区大型河流如西江[31]、东江[32]、桂江[33]..此外,黄江河DOC 浓度与同为小流域河流南苕溪[34]和脱甲河[35]浓度较为接近,而三河DOC 浓度均低于农业型入海河流大沽河[36].

表2 三河荧光光谱参数特征Table 2 Fluorescence spectral parameters of the three rivers

表3 三河荧光组分特征Table 3 Fluorescent components of DOM in the three rivers
三河水体所在地区人口密集,土地利用类型中林地和农田面积占比高,且三河均存在不同程度藻类增殖现象.研究表明,河流DOC 浓度受河流大小、气候、植被、土壤类型、人类活动等因素影响[5].河流DOC 浓度主要由外源输入(陆源和人为源等从地表或者地下径流输入)、内源产生(水生植物光合作用)和河流内的迁移转化(颗粒有机质分解、吸附/解吸附,沉淀、降解等)过程共同决定.多闸坝更多情况下减小了外源输入量,特别是避免高污染的支流汇入.本文认为多闸坝、低流量主要通过改变水体滞留时间影响河流的生物地球化学过程,进而影响DOC浓度.沉积物中颗粒态有机质分解、吸附和与浮游植物相关的一系列过程可能导致河流内DOC 升高,流量小和多闸坝降低水体流速、增加水体透光性和改善浮游植物生境使藻类大量增殖是导致三河DOC浓度较大型河流高的原因之一.
河流在全球碳循环中起到重要作用,然而河流输运的有机碳入海通量仍存在很大误差,主要原因之一是中小型河流数据缺失[37].典型中小型闸控入海河流黄江河、东溪河和练江流量约为珠江水流量的0.5%~1%,但是在外源和内源协同作用下,三河DOC浓度是珠江的1.5~3.0 倍[30],产生的DOC 入海通量为珠江(珠江有机碳入海通量[38]为0.61Tg C)的0.8%~1.5%.华南地区中小型闸控入海河流数量多、分布广,其在区域有机碳入海通量中的作用可能被低估.
2.3 三河DOM 来源解析
2.3.1 三河DOM 荧光特性与来源 三河FI 均值为练江(1.75)>黄江河(1.72)>东溪河(1.66),均大于陆源DOM 荧光指数(1.3),略小于微生物或藻类活动产生的内源DOM 荧光指数(1.8)[39],表明河流DOM 有内源和陆源双重特征,且微生物或藻类活动产生的内源DOM 贡献较大.此外,三河FI 均呈现枯水期>丰水期,表明枯水期内源特征较丰水期更明显.三河自生源指数(BIX)均值为练江(0.83)>黄江河(0.74)>东溪河(0.735),BIX 可以反映水体中DOC 自生源贡献特征,当BIX<0.8 时,DOM 组分中自生源贡献较小,BIX>0.8 时,DOM 组分中自生贡献较高,当>1.0 时,自生源特征显著[10].研究结果表明练江自生源贡献相对较高,黄江河和东溪河自生源贡献比练江略小.三河腐质化指数(HIX)均值为练江(0.64)>东溪河(0.52)>黄江河(0.42),均小于1,呈现枯水期<丰水期,当腐殖化指数HIX<1.5 时,有较强水生生物来源特征,当1.5
2.3.2 藻源DOC 贡献特征 黄江河丰、枯水期叶绿素a 浓度(图4)分别为(20±8)和(20±8)µg/L,东溪河丰、枯水期叶绿素a 浓度分别为(35±9)和(27±13) µg/L,练江丰、枯水期叶绿素a 浓度分别为(69±33)和(87±55)µg/L.三河叶绿素a 浓度丰、枯水期变化不显著;从沿程来看,黄江河和东溪河叶绿素a 浓度沿程没有显著性变化,而练江下游(LJ4~LJ5)叶绿素 a 平均浓度显著(P<0.05)高于中下游(LJ3)点位,LJ3~LJ5 点位平均浓度分别为(36±30), (98±54)和(118±87) µg/L.
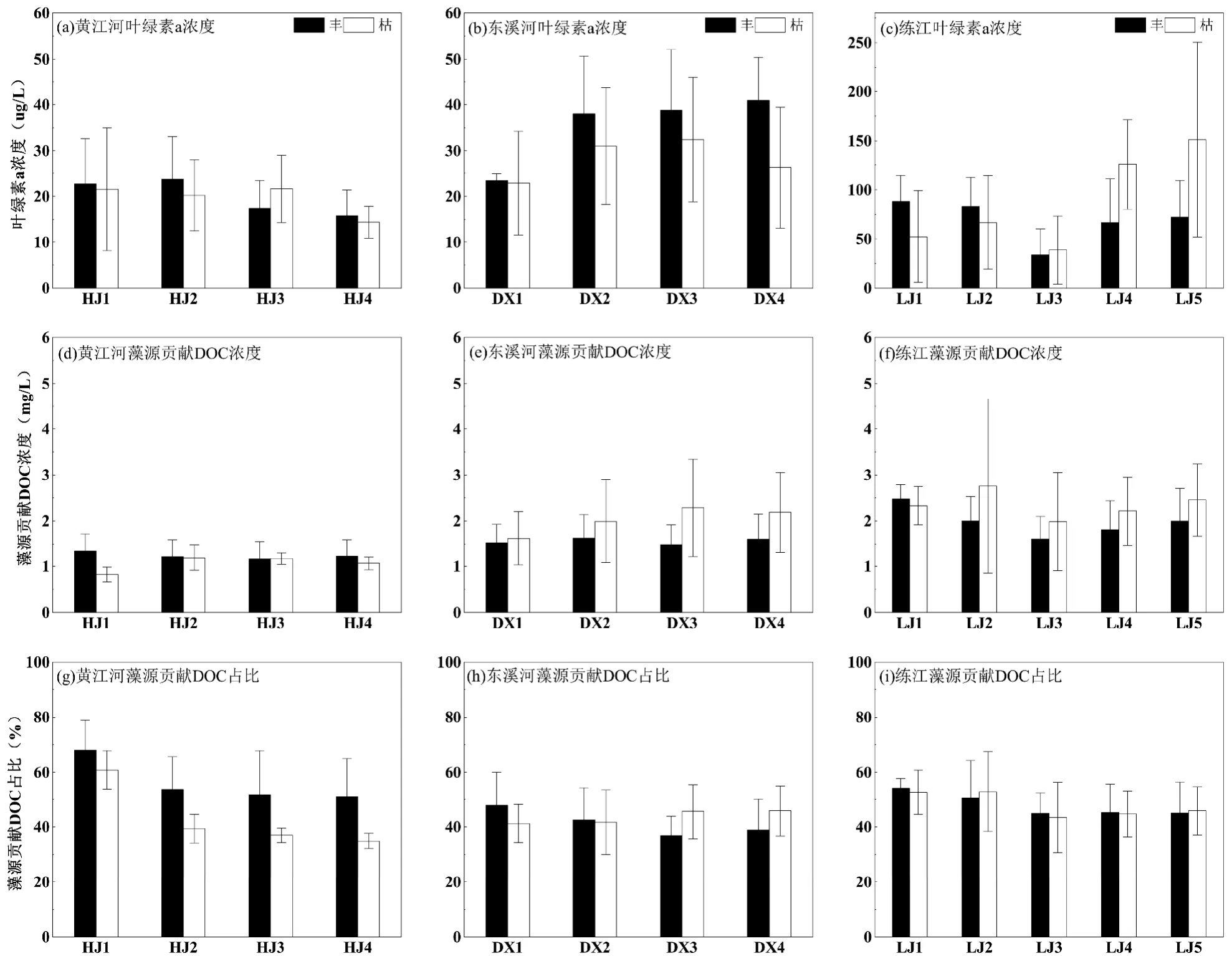
图4 黄江河、东溪河、练江叶绿素a 浓度、藻源贡献DOC 浓度和占比沿程变化Fig.4 Chlorophyll a concentration、algal-derived DOC concentration and proportion in Huangjiang River,Dongxi River and Lianjiang River
藻类中的DOC 具有比陆源DOC 更弱的吸光特性,本文利用藻源DOC 在特定波长时的紫外吸光度结合卡特模型量化三河藻源DOC 浓度[27].黄江河,东溪河和练江藻源DOC 浓度(图4)分别为1.2,1.8,2.2mg/L,占三河DOC 浓度的49.6%、42.6%和48.0%.富营养化水体中,藻类生消过程中产生DOC 对水体浓度贡献较大,Adams 等[11]通过模型预测得出藻类生消过程是长江流域富营养化水体DOC 主要来源,刘金科等[42]研究表明浮游藻类生物量决定了九龙江流域DOC 浓度.三河均呈富营养化状态,且藻源DOC 占比约为50%,表明藻类内源是三河DOC 主要来源,藻类增殖产生的内源可能是三河DOC 比华南地区其它河流浓度高主要原因之一.
时间上,黄江河丰、枯水期藻源DOC 浓度分别为(1.3±0.3)和(1.0±0.2)µg/L,东溪河丰、枯水期藻源DOC 浓度分别为(1.5±0.4)和(2.0±0.8)µg/L,练江丰、枯水期藻源DOC 浓度分别为(2.1±0.4)和(2.2±1.0)µg/L,三河藻源DOC 浓度丰、枯水期变化不显著;从沿程来看,东溪河中下游(DX2~DX4)和练江中下游(LJ2~LJ5),藻源DOC 平均浓度呈枯水期高于丰水期趋势,但三河藻源DOC 浓度沿程无显著变化.分析三河藻源DOC 浓度占比发现(图4),藻源DOC 浓度无显著时间变化和沿程变化(HJ除外).
总体来讲,下游藻源DOC 浓度呈枯水期高于丰水期趋势,可能是因为枯水期挡潮闸处于关闭状态,下游水动力条件减弱,环境更适宜藻类增殖.藻类大量增殖的过程加速无机营养元素向有机形态转化,进而会改变水体有机碳的浓度和形态[43].而藻源DOC 占比变化不大,可能是枯水期藻源DOC浓度增加的同时有陆源DOC 输入,比如鱼塘排水,也可能是华南地区河流生化反应比较活跃,对于藻源产生活性较强的有机质能短时间发生降解,这需要对于华南地区河流有机质迁移转化机制开展进一步研究.
2.4 三河有机质荧光组分特征
利用平行因子分析法对三河三维荧光光谱数据进行分析,共提取出5 种荧光组分(图5),组分C1(275(225)nm/325nm)和C3(230(285)nm/340nm)代表类色氨酸物质和类酪氨酸类物质[44],主要由微生物和浮游藻类降解作用产生,易发生降解[45],在太湖夏季水体DOM 来源解析研究[46]及苏州古城景观河道 DOM 组分研究[47]中发现类似组分.组分C2(230nm/420nm)代表来自生物/微生物活动的类富里酸,通常存在于海洋环境、废水、湿地和农业环境中[48].组分C4(265nm/485nm)被广泛用于表征陆地来源的类腐殖质组分[49].组分C5(220nm/325nm)代表紫外区类色氨酸物质,常见于有外源输入的河道[50].

图5 黄江河、东溪河、练江5 个组分EEMsFig.5 EEMs of the five DOM components in Huangjiang River,Dongxi River and Lianjiang River
时间上,三河表征内源有机质的类蛋白组分C1 和C3 荧光强度和占比枯水期均高于丰水期(表4 和图 6),C1 组分荧光强度占比分别提高了14.88%、25.55%和11.90%,组分C3 荧光强度分别提高了11.95%、20.76%和6.77%.这可能是因为富营养化河流枯水期藻类大量增殖,藻类生消过程导致来自微生物和浮游藻类降解的类蛋白荧光组分C1 和C3 占比升高.表征陆源腐殖质组分C2、C4和类蛋白组分C5 荧光强度时间变化不大(练江C5组分除外),但占比呈丰水期高于枯水期特征,表明三河有稳定的外源输入如生活污水等,而练江荧光组分C5 主要出现在丰水期,枯水期荧光强度基本测定不到,表明这种组分可能主要随降雨进入水体或受降雨影响.
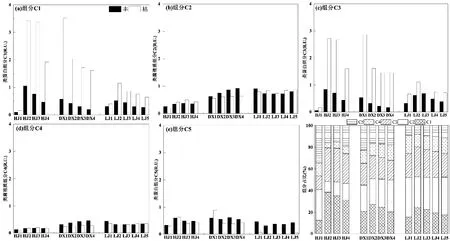
图6 黄江河、东溪河、练江5 个组分荧光强度时空变化及组分占比Fig.6 Intensities and proportions of the five DOM fluorescent components along Huangjiang River,Dongxi River and Lianjiang River
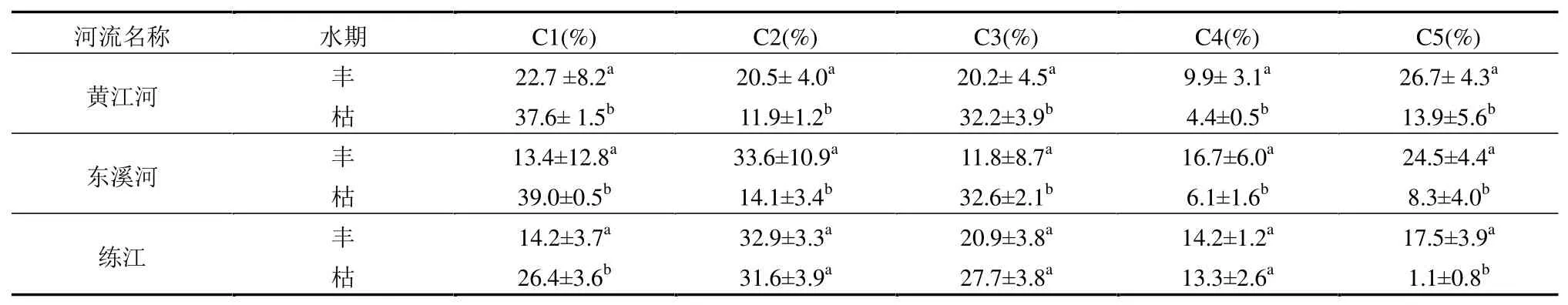
表4 三河荧光组分占比时间变化Table 4 Relative abundance of the five fluorescent components and their temporal variations in the three river
空间上,三河类蛋白组分C1 和C3 沿程变化特征相同,上游(点位HJ1、LJ1)荧光强度低,下游荧光强度高,主要原因是上游污染源较少,受人类活动影响较小.三河腐殖质组分C2 和C4 荧光强度沿程变化特征相同,且沿程变化不大,表明这两种腐殖质来源相同,或受相同条件影响.
三河不仅DOC 浓度高,蛋白组分(C1 和C3)占比也高于大型河流,约为珠江流域河流的 2倍[51].类蛋白有机质通常活性较强,在河口-海湾输运过程中容易被微生物降解、消耗大量氧气并生成CO2,不仅加剧海湾低氧现象,还可能促进温室气体排放.
2.5 DOM 特征与环境因子相关性分析
2.5.1 DOC 浓度与环境因子 三河DOC 均与藻源DOC 浓度呈显著正相关,表明三河有机质藻类内源输入特征明显.三河DOC 浓度受环境因素影响复杂,三河DOC 浓度与水质参数(pH 值、溶解氧、盐度)呈正相关,而与气象参数(水温和降雨)呈负相关.
华南地区光照充足,藻类光合作用较强,富营养化程度高.三河藻类大量增殖通过消耗CO2产生氧气使得水体pH值和溶解氧升高.同时,pH值升高导致水体碳酸盐平衡发生变化,水体CO2分压降低,有利于水-气界CO2吸收和周转,从而进一步促进藻类光合作用[52].三河DOC 浓度与盐度的显著正相关关系表明盐度是影响DOC 浓度变化的因素之一,这与吴红宝等[35]对脱甲河及王江涛等[53]对鸭绿江河口研究结果相一致.当盐度为0~1.6 时,盐度升高会导致水体悬浮颗粒物表面结构发生变化[54],继而使吸附在颗粒物上的DOC 解吸出来.低盐度环境有助于微生物增殖,微生物分解DOC 也是盐度与DOC 呈显著正相关的原因之一[55].
黄江河和练江DOC 浓度与降雨量、水温均呈显著负相关,而东溪河DOC 浓度与降雨量、水温无显著相关性.降雨和温度对DOC 的浓度影响与多种因素有关,例如当流域土壤有机质含量较高时,降雨可以将大量DOC 冲刷进入河流导致河水DOC 浓度升高;而短时间内大量降雨对DOC 浓度的影响则可能主要是稀释作用;高温既可以促进DOC 的合成,也可以促进DOC 的降解.此外,黄江河、东溪河和练江均为闸控型河流,闸控调度主要集中在丰水期(5~9 月份),占总调度次数比例分别为92%、64%和67%.挡潮闸的频繁调度增加了水体的流动性,不利于藻类大量增殖.而枯水期降雨量较少,挡潮闸处于关闭状态,会导致水体流速减慢,水力滞留时间延长,沉积物中养分的积累和微生物丰度的增加促使无机营养盐释放,进一步促进水体中藻类增殖,这些生物生长、代谢进一步产生DOC,导致水体DOC 浓度升高.三河所处的华南地区雨、热同期,降雨、温度和人为干扰(闸控调度)共同作用导致其与DOC 浓度的相关性具有很大的空间差异性.
2.5.2 藻源DOC 与环境因子 藻源DOC 与环境因子相关性分析(图7)表明,黄江河和练江藻源DOC与叶绿素a 呈显著正相关,东溪河藻源DOC 与叶绿素a 无显著相关性.这可能是因为藻源DOC 活性较强,当外界条件适宜时(温度、营养盐浓度等),藻源DOC 会被迅速降解.例如,Bai 等[41]通过室内培养实验发现,藻类生消过程产生的活性(周转时间从几小时到几周)和半活性(周转时间为几个月到几年)DOC 分别占总DOC 的53%和26%,藻源DOC 约有50%会在几天至几周内被降解.河流有机碳迁移转化的作用方向和大小受生物种类和丰度、河流的物理化学特征、气候条件甚至人为干扰的影响,有很大的时空差异[56],因此,藻源DOC 的浓度还受其在河流中迁移转化的机制影响,这可能是三河叶绿素a与藻源DOC 浓度相关性不一致的主要原因.而华南中小型闸控河流溶解有机碳迁移转化机制仍不清楚,需要进一步深入研究.

图7 黄江河、东溪河、练江DOM 特征与环境因子的Spearman 相关性Fig.7 Spearman correlation coefficients between DOM characteristics and environmental factors in Huangjiang River,Dongxi River and Lianjiang River*表示P<0.05,**表示P<0.01
2.5.3 DOM 荧光组分与环境因子 三河类蛋白组分C1 和C3均与水温和降雨量显著负相关,与水质参数(pH 值、溶解氧、盐度)正相关;而三河类腐殖质组分C2 和C4 与水温和降雨量相关性各不相同,与水质参数(pH 值、溶解氧、盐度)相关性也无明显规律.
三河DOM 荧光组分分析结果表明分子量小易降解的类蛋白物质为主要组分,占比分别为77.8%、67.1%和53.3%,三河类蛋白组分C1 和C3之间均显著正相关,表明这两种组分来源相同或受相同条件影响.三河类蛋白组分与水温和降雨量的显著负相关可能是因为华南地区为雨、热同期,温度越高类蛋白组分越容易被降解,同时降雨量增加不利于藻类增殖,从而导致表征内源类蛋白组分含量降低.
三河类腐殖质组分C2和C4与水温和降雨量相关性各不相同,黄江河相关性不显著,东溪河呈显著正相关,而练江呈显著负相关.河流中的类腐殖质组分主要来自陆源DOM,当降雨稀释作用大于冲刷作用,腐殖质含量随着降雨量升高而降低,比如练江.当冲刷作用大于稀释作用时,比如东溪河流域土地利用类型以耕地、林地和果园为主,丰富的面源污染物输入导致东溪河类腐殖质组分含量随降雨量升高而升高.三河类腐殖质组分与水质参数相关性无明显规律,表明陆源腐殖质组分含量主要受气象条件影响下的外源输入的影响,河流内的生物地球化学过程对陆源腐殖质影响较小.
2.6 讨论
首先,本研究证明中小型入海河流DOC 浓度高于华南地区大型河流,其在区域乃至全球碳循环中的作用可能在过去被低估,本研究补充了全球河流有机碳循环数据库,为进一步精确量化DOC 的入海通量提供基础数据.其次,本研究对DOM 浓度和组分时空变化的解析可以指示DOM 在河流-海湾连续体中的迁移转化路径,比如类蛋白质类有机质活性强,在运移的过程中容易被微生物降解产生CO2,从而影响区域碳循环.根据DOM 时空变化特征精确量化污染物入海通量及深入探究DOM 迁移转化路径及其环境效应是研究团队下一步重点开展的工作.
3 结论
3.1 黄江河、东溪河、练江有机碳主要存在形式为DOC,三河 DOC 平均浓度分别为 2.58 ,4.31,5.13mg/L,低于世界河流平均浓度但高于华南地区大型河流.
3.2 荧光参数结果表明三河DOC 具有陆源及内源双重特征,藻源DOC 特征更明显,并且东溪河和练江藻源DOC 平均浓度枯水期高于丰水期.三河藻源DOC 浓度分别为1.15, 1.79, 2.17mg/L,藻源DOC 占比接近50%,分别为49.6%、42.6%和48.0%,表明藻类内源输入是三河DOC 主要来源之一.
3.3 利用平行因子解析出3种类蛋白组分和2种类腐殖质组分,易降解的类蛋白组分占比较大.此外,受藻类异常增殖影响,表征藻类内源的类蛋白组分C1和C3 总荧光强度枯水期显著高于丰水期,表征来自陆源类腐殖质组分C2 和C4 荧光强度呈练江>东溪河>黄江河.
