表观气速对除磷污泥颗粒化性能及代谢特征的影响
2024-03-08张瑞峰宫延哲蔡虎林曹银焕杨文浩穆瑞花西安工程大学城市规划与市政工程学院陕西西安70048西安净水处理有限责任公司陕西西安7006西安工程大学环境与化学工程学院陕西西安70048
陈 希,胡 彬,张瑞峰,宫延哲,蔡虎林,曹银焕,杨文浩,穆瑞花 (.西安工程大学城市规划与市政工程学院,陕西 西安 70048;.西安净水处理有限责任公司,陕西 西安 7006;.西安工程大学环境与化学工程学院,陕西 西安 70048)
与传统活性污泥系统相比,好氧颗粒污泥系统可减少20%~50%的能源消耗和25%~75%的占地面积[1].表观气速的大小对污泥颗粒化影响很大.高表观气速引起的强水力剪切力会加快污泥颗粒化,抑制丝状菌的过度生长[2],而低表观气速不利于污泥颗粒化[3].Tay 等[4]研究发现,在表观气速为0.3cm/s时,没有发生污泥颗粒化,而在表观气速为1.2cm/s时,可以诱导颗粒污泥分泌更多的胞外多糖,促进形成结构紧密的颗粒污泥.虽然高表观气速能促进絮状污泥的颗粒化[5],但却意味着更高的能耗,此与好氧颗粒污泥节能降耗的初衷存在矛盾.
研究表明,采用厌氧/好氧交替模式筛选具有胞内储存功能的缓慢生长微生物(如传统聚磷菌和聚糖菌),即使在较低的剪切力下也可以培养出性能稳定的好氧颗粒污泥——生物除磷颗粒污泥.研究发现,以乙酸钠为碳源通过厌氧-好氧交替环境可以成功培养好氧颗粒污泥,这些颗粒污泥即使在较低的剪切力和溶解氧条件下(40%甚至20%饱和溶解氧)也能稳定存在[6-7].目前,研究者尝试采用低表观气速或低溶解氧培养好氧颗粒污泥,多以小分子有机物为碳源,对大分子碳源的关注较少.
在厌氧好氧交替的生物除磷系统,采用小分子碳源易实现厌氧饱食好氧饥饿代谢模式,污泥更易颗粒化.但实际生活污水中小分子碳源占比较低,缓慢降解的大分子有机物才是碳源的主体,约占总COD 的50%以上.由于大分子碳源需经过复杂的水解发酵过程才能实现厌氧储存[8],因此常常延缓甚至阻碍污泥的颗粒化.Layer 等研究[9]发现,污水中存在不可扩散的有机基质(大分子有机物)会导致小颗粒的形成,同时当复杂基质增多时,发酵功能菌群比传统聚磷菌和聚糖菌更易富集,系统中的絮体污泥增多.在生物除磷污泥系统中,表观气速不仅提供剪切力,同时可能影响大分子碳源的水解发酵和胞内储存,进而影响污泥的颗粒化.但是目前关于复杂碳源条件下表观气速对污泥颗粒化影响的研究较少.
本研究以小分子和大分子有机物复合基质为碳源(60%NaAc+40%可溶性淀粉),考察复杂碳源条件下,表观气速对除磷污泥颗粒化性能及代谢特征的影响,从碳源代谢和菌群结构的角度,揭示复杂碳源条件下表观气速影响生物除磷污泥颗粒化的机理,以期为好氧颗粒污泥技术的应用提供参考.
1 材料与方法
1.1 实验装置与运行方式
采用3 组等工作体积的SBR 反应器(图1),材料为有机玻璃.3 组反应器的高径比相同且体积相同,高度均为110cm,内径7.5cm,有效水深92cm,高径比12.27,有效容积为4L,排水比为50%.曝气量由转子流量计控制,分别为1.2、0.8、0.4L/min,对应表观气速分别为0.45 (R1)、0.30 (R2)、0.15(R3) cm/s.采用蠕动泵(保定兰格BT100-2J)控制进水,由反应器底部的布水盘均匀布水,进水的同时通过顶部溢流口推流出水.采用恒温水浴系统(西安东瑞YHX-030)将反应器保持在(20±1)℃下运行.
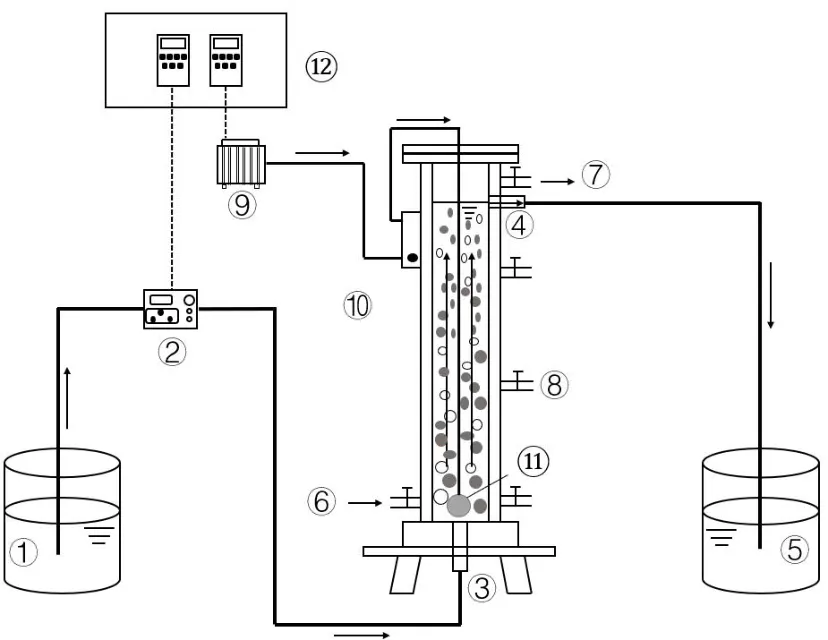
图1 等工作体积SBR 反应器示意Fig.1 Diagram of the SBR with constant working volume
反应器进出水、曝气等程序通过时控开关自动控制.每天运行6 个周期,每个周期4h,包括60min 同步进出水(推移流),30min 厌氧静置,120~145min 曝气,以及30~5min 沉淀(在32d 内通过延长曝气时间将沉淀时间从30min 逐渐减少至5min).在运行过程中,反应器中水面高度保持不变,即为等工作体积.反应器不设置机械搅拌,泥水混合通过曝气实现;不控制污泥龄,仅通过测定污泥浓度和胞内外代谢排泥.
1.2 接种污泥及进水水质
接种污泥采用西安市第四污水处理厂回流污泥.污泥为絮体,结构松散,沉降性较差,丝状菌较多,主要为微丝菌(Microthrixparvicella).用孔径1mm 的筛子将取回的污泥过滤,筛除杂质后移入反应器.
以60%乙酸钠+40%可溶性淀粉为碳源模拟生活污水,进水COD 为400mg/L;总氮为45mg/L,其中5mg/L 硝氮由配水所用自来水带入,其余氮素由NH4Cl 提供;总磷为10mg/L,由60%的K2HPO4和40%的KH2PO4提供.每升模拟污水中分别加入1mL微量元素和10mL 矿物质浓缩液[10],微量元素和矿物质浓缩液组分及其浓度见表1.
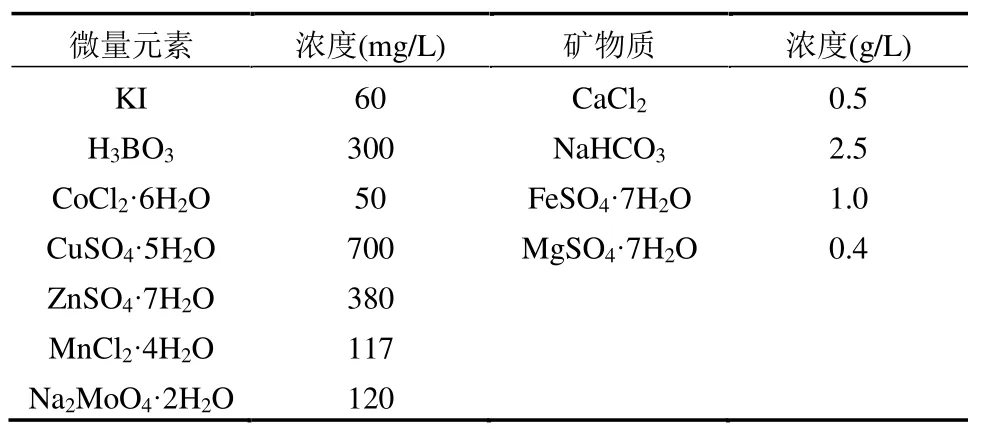
表1 微量元素和矿物质浓缩液组分及其浓度Table 1 Components and concentrations of trace elements and minerals in concentrated solution
1.3 分析方法
1.3.1 常规指标分析方法 常规水质指标的检测参考《水和废水监测分析方法》(第4 版)[11].污泥浓度(MLSS)测定从反应器中部取样,SVI30和SVI5分别为污泥在曝气末期沉降30min 和5min 时的污泥体积指数.采用SVI30/SVI5表示污泥的颗粒化进程,当SVI30和SVI5均较低时,该值越接近1,表明污泥颗粒化效果越好[12].采用光学显微镜(Olympus BX53)每周对污泥进行一次常规镜检.污泥粒径的测定采用激光粒度分析仪(S3500).COD 测定采用重铬酸钾法,测定采用紫外分光光度法,PO43--P 测定采用钼锑抗分光光度法[11].常规水质和污泥特性测定从反应器中部取样;典型周期内水质和污泥特性测定,厌氧阶段从反应器底部取样,好氧阶段从反应器中部取样.
1.3.2 污泥胞外聚合物的提取和测定 污泥胞外聚合物(EPS)提取采用超声+阳离子交换树脂法,采用修正的Lowry 法测定蛋白质,牛血清蛋白为标准物质;采用蒽酮比色法测定多糖,葡萄糖为标准物质[13].每30d 对EPS 进行1 次提取和测定,连续监测3 组系统EPS 的变化.
采用傅里叶红外光谱仪(IRPrestige-21)测定疏松型EPS(LB-EPS)和紧密型EPS(TB-EPS)的红外光谱,具体方法为:将待测样品冷冻处理(-20℃ ), 然后使用冷冻干燥机将冷冻后的样品在-55℃下冷冻干燥24h.将冻干后的样品粉末和烘干后的溴化钾(100℃下干燥5h)按1:50 的配比称量混合,于研磨机中研磨均匀后压片(厚度小于0.5mm).采用傅里叶红外光谱仪对样品进行扫描,分辨率为0.4 /cm,测定波长范围为400~4000/cm,扫描32 次.随后对扫描所得红外光谱中蛋白质酰胺I 区(1600~1700 /cm)的二阶导数采用PeakFit 进行分峰拟合,统计代表不同结构的各峰面积,计算α-螺旋/(β-折叠+无规则卷曲)的数值.该数值越小,表明EPS 的疏水性越高[14].
1.3.3 挥发性脂肪酸和乳酸的测定 挥发性脂肪酸(VFAs)的测定采用气相色谱法(双检),色谱柱为PE WAX TER 型毛细管柱(30.0m×250µm×0.25µm),检测器为氢火焰离子化检测仪(FID).采用程序升温的方法,初始温度为 100℃,初始时间为 2min,以3 ℃/min 的速度升温至160℃后停留1min.氢焰室温度230℃,N2、H2和空气气速分别为20,35,350mL/min.进样口(汽化室)温度为200℃,载气为高纯氮气,分流比10:1,微量进样器进样1μL.乳酸参照诸葛健等[15]和高庆等[16]的方法,采用对羟基联苯法测定.VFAs和乳酸取样时间均在第215d,此时为反应器运行后期,3 组系统均较为稳定.
1.3.4 胞内储存物的测定 生物除磷污泥的厌氧胞内储存物主要为聚羟基烷酸酯(PHAs)和糖原,其中聚β-羟基丁酸盐(PHB)和聚β-羟基戊酸盐(PHV)为PHAs 的主要成分[17].本研究中PHB 和PHV 均采用气相色谱仪(PE680)测定,色谱柱为HP-5 型色谱柱(30m length×320μm ID×0.25μm film),进样方式为自动进样(进样量2μL),检测器为氢火焰离子化检测器(FID),采取恒定流模式,以N2作为载气,进样口温度250℃,分流比为2.7:1,分流流量为2mL/min,FID检测器的工作温度为300℃.空气流量为400mL/min,尾吹气流量为20mL/min,氢气流量为40mL/min,采用程序升温,从50℃升温至270℃,之后保持6min.糖原采用蒽酮比色法测[18]定.PHB、PHV 和糖原的预处理均采用冻干法[19],取样时间为第215d.
1.3.5 16S rRNA 基因高通量测序 采用 16S rRNA 基因高通量测序对接种污泥和3 组反应器关键时间节点(污泥颗粒化初期及培养末期)污泥的菌群结构进行解析(上海美吉生物医药科技有限公司).3 组反应器颗粒化初期的时间节点分别为:90、200、140d,培养末期时间节点均为256d.具体分析方法为:采用E.Z.N.A.® soil 试剂盒(Omega Biotek,Norcross,GA,U.S.)提取污泥的总DNA,DNA 浓度和纯度采用NanoDrop2000 进行检测,采用1%琼脂糖凝胶电泳检测DNA提取质量.聚合酶链式反应(PCR)扩增子文库使用Illumina MiSeq PE300 平台构建,引物 338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)[10].采用UPARSE 软件,对序列进行OTU 聚类(97%的相似度),并在聚类过程中去除单序列和嵌合体.利用RDP classifier 对每条序列进行物种分类注释,和Silva 数据库(SSU123)进行比对,最后对所得的细菌分类信息进行多样性和群落组成分析.
2 结果与讨论
2.1 系统的污泥浓度及沉降性能变化
由图2 可见,R3 反应器由于水力剪切力最小,污泥流失较小,在整个运行过程中MLSS 始终最高,保持在8000~10000mg/L.R1 和R2 由于表观气速均较高,MLSS 在60d 前不断减低,变化规律基本一致,2个反应器的MLSS 在第80~90d 出现拐点,此后持续上升.R2 的表观气速和水力剪切力低于R1,但MLSS在运行中期(60~110d)比R1低2000~ 3000mg/L,此现象与R3 污泥浓度整体较高的现象并不一致.这是由于在R2的表观气速下,系统中孳生了大量的仙女虫,这种后生动物不断吞食和破坏污泥絮体和颗粒污泥雏形[20],导致MLSS 最低降至2300mg/L.在第78d将仙女虫用200μm 筛网筛除,R2 的MLSS 短暂降低后持续升高.表观气速在提供水力剪切力的同时,也改变了系统的微生物组成和污泥状态.
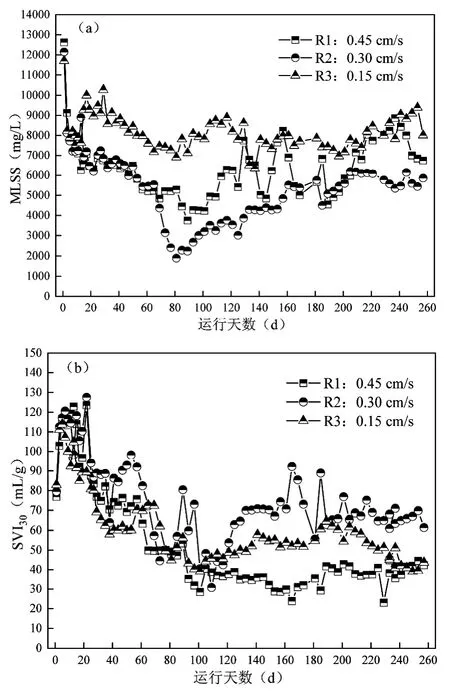
图2 三组反应器中MLSS 和SVI30 的历时变化Fig.2 Variations of MLSS and SVI30 in the three reactors
在R1 运行后期(111~260d),其中部取样测得的MLSS 在4500~9000mg/L 呈周期性波动,但污泥沉降性始终很好,SVI30为20~45mL/g.R1 中下部污泥浓度不断升高,并沿轴向呈梯度分布,241d 反应器上部(液面以下23cm)、中部(液面以下46cm)和下部(液面以下69cm)的MLSS 分别为7289,8434,8921mg/L.表观气速越大反应器中污泥所受水的剪切力越大,R1 由于颗粒化进程最快,粒径越大的污泥受重力影响越大,因此离曝气盘越近污泥浓度越高.而在表观气速更小的R2和R3中,由于污泥颗粒化进行较慢,污泥粒径较小.尽管剪切力较小,曝气盘提供的水力强度仍能保证污泥的完全混合,因此R2 和R3 中污泥浓度分布均匀,并未出现沿反应器轴向梯度分布的现象.在R1 反应器中,伴随着污泥浓度的周期性波动,厌氧静置阶段泥面高度不断增加.R1 反应器污泥浓度的周期性降低并非由污泥流失引起,而是污泥粒径增大后其悬浮位置向反应器下部移动的结果.因此推断R1 在该阶段存在颗粒污泥再生过程,规律为新生颗粒-悬浮中上部-悬浮底部-再次新生颗粒.
污泥沉降性较好时,SVI30/SVI5越接近1 代表污泥颗粒化效果越好[12].由图3(a)可知,R1 颗粒化进程最快,在110d 基本实现颗粒化;R2 在200d 后开始快速颗粒化;R3 在140d 后快速颗粒化,至190d 实现颗粒化.3 组反应器污泥稳定性良好,直至实验末期MLSS 和污泥粒径仍不断增大.图3(b)中225d 3 组反应器污泥的粒径分布,中值粒径分别为374,192, 291μm,污泥颗粒化进程由快至慢依次为R1、R3 和R2.

图3 三组反应器中SVI30/SVI5 的历时变化和第225d 污泥的粒径分布Fig.3 Variations of SVI30/SVI5 in the three reactors and distribution of particle size on day 225
van Dijk 等[21]以数学模拟的方法探究好氧颗粒污泥的颗粒化机理,发现采用厌氧-好氧交替环境筛选缓慢生长微生物形成好氧颗粒污泥时,污泥的颗粒化存在一个“迟滞期”,约在120d 附近,之后污泥将快速颗粒化.本研究中3 组反应器均出现了污泥颗粒化的迟滞期,R1 的表观气速最大,污泥颗粒化的迟滞期最短,大约持续60d;R2 表观气速虽然介于R1 与R3 之间,但颗粒化的迟滞期最长,为200d;R3 表观气速最小,迟滞期长于R1 但短于R2,约为140d.不同表观气速下反应器的“迟滞期”并非随表观气速的降低而递减,这可能由于表观气速不仅提供水力剪切力,同时影响微生物的碳源代谢.
2.2 污泥的外观形态变化
由图4 可见, R1 颗粒污泥粒径大,形状规整,边界清晰.R2 絮体较多,颗粒粒径较小,边界较为清晰,形状较为规整.R3 颗粒边界不清晰,形状不规整,含有较多丝状菌,颗粒粒径较为均匀且小于R1.R1、R2和R3 污泥的密实度依次降低.Liu 等[22]研究剪切力在生物膜和颗粒污泥形成中的作用时发现,2.5cm/s的表观气速可形成致密、形状规则的颗粒污泥.Beun等[23]也在高表观气速下(4.1cm/s)培养出了光滑、致密的颗粒污泥.此与本研究R1 和R2 的结论一致.但由于R2 在200d 后才开始颗粒化,培养末期(260d)仍在颗粒化中,因此絮体占比较高.Yang 等[24]在探究不同DO 对颗粒污泥稳定性的影响时发现,降低曝气量后,颗粒粒径明显降低.因此R3中较小的粒径和较低的密实度与较低的表观气速有关.
2.3 除磷污泥系统的处理性能
3 组反应器中氨氮的去除率为100%,亚硝氮的去除率接近100%,如图5 所示.3 组反应器COD 的去除率分别为88%、88%和94%.随着表观气速的降低,总氮的去除率依次升高,分别为54%、61%和67%.尽管R1和R3中污泥的中值粒径和氧的传质阻力较大,但是表观气速相比粒径对氮去除的影响更大.较低的表观气速增大了颗粒内部的缺氧区,同时促进了大分子碳源的水解发酵,为反硝化提供了更多的易降解碳源,因此提高了脱氮效率.周赟成等[25]在研究曝气强度对好氧颗粒污泥稳定性的影响时同样指出,降低曝气量可以强化同步硝化反硝化作用,提高系统的脱氮效率.
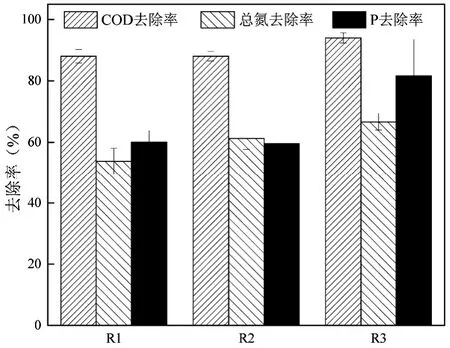
图5 三组反应器污染物去除率Fig.5 Removal efficiency of the pollutants in the three reactors
3组反应器中磷去除率的均值分别为60%、60%和82%,R2 和R3 磷去除率的变化幅度较大.R1 表观气速最高,污泥除磷率较低,并随着时间的延续不断降低.由于本研究主要关注污泥的颗粒化,未进行规律排泥,因此可能导致R1 中GAOs 丰度增加,与PAOs 竞争碳源导致除磷率逐渐下降[26].R2 在运行中期由于仙女虫的孳生和聚磷菌的流失,除磷率下降较多;但在运行末期发酵菌群与聚磷菌协同生长,除磷率大幅升高(该过程伴随着污泥的快速颗粒化),故除磷率变化较大.R3 的除磷率始终较高,这是由于R3 的低表观气速有利于发酵菌群的生长,系统中水解发酵菌群和除磷菌群的协同作用更好.R3 的除磷率在160d前变化不大,磷去除率始终较高;但是160d后污泥快速颗粒化,磷去除率有所降低,导致系统的除磷率变化较大.R3 的表观气速最低(0.15cm/s),提供的剪切力最小,在污泥颗粒化后无法使大粒径的颗粒污泥有效悬浮,部分大颗粒沉积在反应器底部而呈黑色,导致好氧阶段出现了底部污泥的厌氧释磷,降低了磷的去除率.为保证系统较高的磷去除率,好氧颗粒污泥系统可采取2 种方法强化磷的去除:(1)规律性排泥,促进PAOs 的优势生长;(2)在低表观气速系统中,采取一定措施避免好氧阶段大颗粒沉积在反应器底部,如增加水力循环或者机械搅拌等.
综上可知,降低表观气速可通过促进大分子碳源的水解发酵同步提高系统的脱氮和除磷效率.大颗粒的有效悬浮对除磷效果影响很大,低表观气速时需采取一定的措施避免大颗粒的沉积.
2.4 污泥的胞外聚合物
2.4.1 多糖和蛋白质含量 图6(a)中,根据污泥EPS 的变化规律和颗粒化进程以110d 为界解析污泥的颗粒化过程.在110d 前(第I 和第II 阶段),R1 和R2 污泥中EPS 的多糖含量较高,颗粒化速度较快,而R3 中EPS 的多糖含量较少,颗粒化速度较慢.在110d 后(第III 阶段),3 组反应器LB-EPS 中多糖含量的均值依次增加,TB-EPS 也呈现相似的规律.110d后R1 系统已基本完成颗粒化,多糖含量有所降低,R2 和R3 反应器颗粒化速度加快,多糖含量逐渐升高.此外,R3 系统在此阶段污泥颗粒化速度最快,EPS中多糖含量最高.EPS 可通过改变细胞表面性质,提高疏水性,与多价金属离子等结合,促进污泥颗粒化[27].EPS 中的多糖可通过提高絮体间的黏附作用促进污泥颗粒化[28].Tay 等[4]研究表明,好氧颗粒污泥系统中的剪切力与多糖的产生、细胞表面疏水性等均与呈正相关.在110d 前,R1 较高的剪切力通过促进EPS 的分泌加快了污泥的颗粒化;R2 表观气速介于R1 和R3 之间,尽管EPS 较高,但其正面影响被仙女虫的负面影响所掩盖;R3 表观气速最小,EPS 较低,污泥颗粒化较慢.110d后,R3LB-EPS较高的多糖含量与聚糖菌丰度的增加有关.
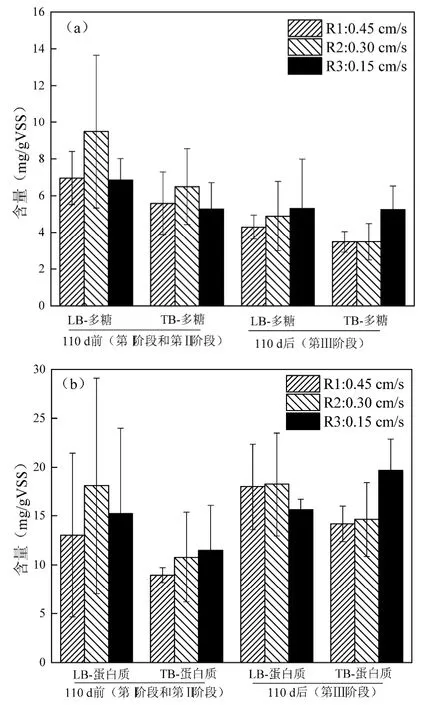
图6 三组系统污泥中EPS 多糖和蛋白质的含量Fig.6 Contents of polysaccharide and protein of the EPS of the sludge in the three reactors
由图6(b)可见, R1 和R3LB-EPS 和TB-EPS 中蛋白质的含量均在110d 后明显增加,此过程伴随着颗粒污泥的逐渐稳定.R2 由于尚处在污泥颗粒化早期,LB-EPS中蛋白质的变化较小,但TB-EPS的蛋白质增加明显.Yang 等[24]指出改变DO 值后会影响EPS 中蛋白质的组成,从而对好氧颗粒污泥的稳定性产生影响.Zhang 等[29]发现蛋白质含量降低使颗粒污泥的稳定性变差.因此推测EPS 中蛋白质的增加与颗粒污泥的逐渐稳定有关.
2.4.2 EPS 的红外光谱分析 由图7(a)可见, R1 反应器在170~235d 逐渐进入颗粒污泥“再生”阶段;R2反应器自200d 后污泥沉降性逐渐好转,不断颗粒化;R3 反应器从140d 开始沉降性不断好转,污泥逐渐颗粒化.采用厌氧好氧交替模式培养颗粒污泥时,无论表观气速高低,随着时间的延续,LB-EPS 蛋白质酰胺I 区α-螺旋/(β-折叠+无规则卷曲)的数值不断降低,疏水性的增加是一个必然趋势,因此即使R1、R2 和R3 反应器颗粒化进程不同,但污泥均向颗粒化的方向发展.由图7(b)可见,随着表观气速的降低,TB-EPS 中α-螺旋/(β-折叠+无规则卷曲)的中位数依次增加,分别为0.59、0.71 和0.81,TB-EPS 的疏水性依次降低.随着表观气速的降低,污泥聚集体中细菌之间的结合能力由于疏水性的降低而降低,污泥的密实度随之降低,与镜检结果一致(图4e~f).

图7 三组反应器污泥中LB-EPS 和TB-EPS 中α-螺旋/(β-折叠+无规则卷曲)不同时期的变化Fig.7 Changes of α-helix/(β-folding + random curling) of LB-EPS and TB-EPS of the sludge in the three reactors during different periods
2.5 碳源的水解发酵和胞内存储
2.5.1 典型周期内COD、挥发性脂肪酸及乳酸变化情况 图8 中,取样时间为215d.饱食饥饿是影响污泥颗粒化的重要因素,可以用COD 的变化来描述反应器的饱食饥饿情况[30],由图8(a)可见,在厌氧阶段,3组反应器的COD 在进水末期分别达到228,252,240mg/L.在好氧阶段,3 组反应器的COD 不断减少,曝气30min 后即降低至100mg/L,曝气60min 后降低至60mg/L 以下,周期末COD 分别为48,24,24mg/L.3组反应器在厌氧段碳源丰富,而在好氧段胞外碳源匮乏,实现了厌氧饱食好氧饥饿,同时R2 和R3 的好氧饥饿程度更高.图8(b)中,在厌氧阶段,R1、R2 和R3 对乙酸的利用速率依次升高.R1 和R2 系统分别在30,15min 内将乙酸利用完全,而R3 系统在进水的前30min 内未检测到乙酸.在整个周期内,R1 和R2未出现VFAs 的累积,R3 在30min 出现乙酸的累积.生物除磷系统中乳酸是淀粉水解发酵的重要中间产物[31].图8(c)中,随着表观气速的降低,乳酸峰值出现的时刻逐渐提前.R1 和R2 在75min 时乳酸浓度达到峰值,R2 在75min 前乳酸浓度始终大于R1,而R3在15min乳酸浓度达到峰值.曝气结束后,3组反应器乳酸浓度依次降低,分别为1.84,1.48,0.20mg/L.

图8 典型周期内COD、VFAs、乳酸及DO 变化曲线Fig.8 Changes of COD、VFAs、lactic acid and DO in a typical cycle
厌氧阶段的发酵产物“泄漏”到好氧阶段的量越多,系统实现好氧饥饿越困难,系统越不易稳定.Haaksman 等[32]探究易降解COD 的好氧利用对好氧颗粒污泥形态稳定性的影响时提出,碳源的泄露会导致颗粒污泥的不稳定.降低表观气速,有利于淀粉水解发酵为VFAs 等小分子有机物.R3 在厌氧阶段乳酸产生最快,在30min出现乙酸累积但75min后耗尽;在好氧阶段乳酸浓度始终低于R1 和R2,好氧末期几乎无残留,系统更易实现污泥颗粒化的理想环境——厌氧饱食好氧饥饿.在R1 和R2 中,由于乳酸产生速率较慢,进入好氧阶段时系统残余的乳酸较多,尽管该阶段乳酸不断下降,但直至好氧末期仍有少量残留,R1 的残余量大于R2.R1 中较大的剪切力抵消了碳源“泄漏”的负面影响,颗粒化较快;但R2 剪切力低于R1,不足以抵消碳源“泄漏”的不利影响,系统不易稳定,因此颗粒化进程最慢.
2.5.2 污泥胞内贮存物的变化情况 由表2 可知,215d 时,随着表观气速的降低,3 组反应器在厌氧阶段PHAs(PHB 与PHV 之和)的合成量依次增加,分别为2.28,8.16,23.96mg/gVSS,糖原的降解量依次减小.在以乙酸钠或以添加了乙酸钠的生活污水为碳源的好氧颗粒污泥系统中,PHAs 的含量约为19.45~42.62mg/gVSS[33-34],本研究中R1 和R2 反应器中PHAs 的厌氧储存量略低,但是R3 中PHAs 的厌氧储存量与文献相当.在好氧阶段,3 组反应器PHAs 的降解量依次增大,分别为 2.25,8.35,25.53mg/gVSS;糖原的最大合成量R1>R3>R2.厌氧条件下,聚磷菌(PAOs)水解聚磷酸盐产生能量用以吸收VFAs 合成PHAs(主要为PHB 和PHV)[24];在好氧阶段,PAOs 分解PHAs 获得能量和碳源,菌体内的糖原和聚磷酸盐含量增加.尽管进水中的大分子碳源不易被PAOs 和GAOs 储存于胞内,但是在进水碳源相同的条件下,随着表观气速的降低,大分子碳源的水解发酵更为彻底,生成VFAs 更多更快,在厌氧条件下储存于胞内的碳源更多,更易实现厌氧饱食好氧饥饿.因此R3系统水力剪切力最小,但污泥颗粒化进程快于R2.

表2 典型周期内胞内贮存物的合成量及消耗量(mg/gVSS)Table 2 Synthesis and consumption of PHB、PHV and glycogen in a typical cycle (mg/gVSS)
2.6 污泥的菌群结构解析
2.6.1 菌群结构门水平的差异 由图9(a)可见,在接种污泥中,优势菌门为变形菌门(Proteobacteria,13.67%)、放线菌门(Actinobacteriota,31.28%)、绿弯菌门(Chloroflexi,26.54%)、拟杆菌门(Bacteroidetes,10.79%) 和 Patescibacteria(7.52%), 而厚壁菌门(Firmicutes,4.58%) 和酸杆菌门(Acidobacteriota,2.41%)丰度较低.

图9 细菌门水平和属水平的群落结构解析Fig.9 Analysis of community structure at phylum level and genus level
随着表观气速的降低, 变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)和放线菌门(Actinobacteriota)的丰度依次降低.至培养末期,3组反应器中变形菌门(Proteobacteria)的丰度较接种污泥大幅升高,分别为51.02%、49.95%和40.11%.变形菌门(Proteobacteria)是好氧颗粒污泥系统中常见的主要菌门[35],这类微生物能通过分泌EPS 促进絮状污泥之间相互凝聚,在污泥颗粒化中起着重要作用[36].拟杆菌门(Bacteroidetes)的丰度较接种污泥变化不大,分别为16.89%、12.45%和9.55%.由于接种污泥中的微丝菌(Microthrix,属于放线菌门)在培养过程中被淘汰,3 组反应器中放线菌门(Actinobacteriota)的丰度较接种污泥均大幅降低,分别为16.89%、12.45%和9.55%.
降低表观气速有利于绿弯菌门(Chloroflexi)的优势增殖.至培养末期,3 组反应器绿弯菌门(Chloroflexi)的丰度依次升高,分别为7.86%、8.77%和32.74%.Bovio-Winkler 等[37]利用宏基因组探究Chloroflexi 在厌氧氨氧化、活性污泥和产甲烷反应器中的作用时发现,Chloroflexi 具有水解复杂有机物的功能,一些Chloroflexi 的菌属在分泌胞外多糖方面有较积极的作用,部分Chloroflexi 具有促进细胞聚集的能力,有助于颗粒污泥初始框架的形 成.
2.6.2 菌群结构属水平的差异解析 在属水平筛选相对丰度大于1%的菌群进行解析,如图9(b)所示.在接种污泥和3 组反应器中,缓慢生长微生物中的传统聚磷菌CandidatusAccumulibacter的丰度远低于1%,故未分析其作用.而传统聚糖菌Candidatus_Competibacter的丰度较高,是污泥颗粒化的核心菌群.R1 表观气速最大,Candidatus_Competibacter在培养前期和后期大量富集,90d 和256d 的丰度分别为18.85%和22.29%.R2 运行200d 时Candidatus_Competibacter丰度为11.45%,256d 时降低至4.93%,表明中等表观气速(0.30cm/s)不利于Candidatus_Competibacter的维持和优势增殖.R3 运行140d 时Candidatus_Competibacter丰度为3.58%,运行256d时增高至25.73%.这主要是因为在低表观气速下,大分子有机物水解发酵条件较好,在培养末期产生了充足的VFAs,促进了Candidatus_Competibacter的优势增殖.Candidatus_Competibacter在厌氧阶段通过酵解糖原产生能量吸收VFAs 并将其储存为PHAs,在好氧阶段氧化PHAs产生能量,合成糖原[38].Candidatus_Competibacter还能分泌大量胶状胞外多糖[39],有助于微生物的团聚,因此在R1 和R3 污泥的颗粒化过程中发挥了重要作用.
系统中可确认的聚磷菌为Tetrasphaera和Amaricoccus[40].Tetrasphaera是近年来关注的发酵型聚糖菌,该菌在接种污泥也即城市污水处理厂活性污泥中丰度较高(2.05%),在表面气速最高的R1反应器中始终维持较高的丰度(4.48%和3.11%).在培养末期(256d),随表观气速的降低,Tetrasphaera在3组反应器中的丰度依次降低,分别为3.11%、0.73%和0.06%,这表明该菌对剪切力敏感,不易在低表观气速的系统中优势生长.Tetrasphaera在厌氧环境下能够降解多聚磷酸盐并能发酵复杂有机物产生能量合成糖原,在好氧环境分解糖原,为生长提供能量并摄取PO43--P 合成聚磷酸盐[41].在R1 的淀粉降解和除磷中可能发挥了一定的作用.Amaricoccus是系统中的另一种聚磷菌[40],该菌的外观与Tetrasphaera相似,也为四联球菌,属于G-细菌的一种[42],好氧条件下可在胞内储存PHAs.其在R2 颗粒化前期(200d)和运行末期(256d)丰度均较高,分别为20.20%和7.69%.McIlroy 等[43]探究SBR 处理酒厂废水中的微生物群落时发现,Amaricoccus具有储存PHAs 的功能.Amaricoccus在好氧条件下具有很强的储存PHB的能力,但在厌氧条件下并没有储存PHB 的能力[44].由于在厌氧条件下无法储存PHB,相比其它聚磷菌和聚糖菌,Amaricoccus的竞争力较弱,所以在培养末期丰度降低.R2 在培养末期仍处于快速颗粒化阶段,故其颗粒化的核心菌群并不完全明确,根据已有数据推测Candidatus_Competibacter和Amaricoccus等具有胞内储存功能的菌群发挥了重要作用.
由于进水中含有40%的可溶性淀粉,系统中存在较多的水解发酵菌群,包括Saccharimonadales[45]、Saprospiraceae[46]、Tetrasphaera和一些绿弯菌门(Chloroflexi)的丝状菌(Caldilineaceae和Kouleothrix)等.除微丝菌(Microthrix)外,norank_f_Caldilineaceae和Kouleothrix是接种污泥中的主要丝状菌,丰度分别为4.90%和4.03%.随着表观气速的降低,在培养末期norank_f_Caldilineaceae的丰度依次增加,分别为0.96%、1.30%和2.78%.在R3 反应器中Kouleothrix(1851 型丝状菌)的丰度始终较高,污泥颗粒化前期和颗粒化后丰度分别为6.48%和10.44%.
R3 在系统运行140d 时,大分子有机物的降解主要 由Kouleothrix、norank_f_norank_o_Saccharimonadales、norank_f_Saprospiraceae、Tetrasphaera、norank_f_Chitinophagaceae完成,丰度分别为6.48%、6.97%、4.03%、3.75%和2.89%.培养至256d时,Kouleothrix的丰度升高至10.44%,其余4 个菌群丰度均有所降低,分别为0.12%、1.75%、0.06%和1.90%,因此在培养后期降解功能主要依靠Kouleothrix(1851 型丝状菌).1851 型丝状菌是一种能够发酵大分子有机物的丝状菌,当系统内糖类较多时,这种丝状菌容易大量繁殖[47].R3 较低的表观气速为Kouleothrix的大量生长和水解发酵大分子碳源创造了有利的水力条件和氧化还原环境,该菌一方面为颗粒污泥提供丝状骨架,另一方面分泌胞外聚合物和小分子碳源促进Candidatus_Competibacter等缓慢生长微生物的附着生长和优势增殖,促进了污泥的颗粒化.
2.7 不同表观气速下除磷污泥颗粒化的机理
以60% NaAc+40%可溶性淀粉研究复杂碳源条件下生物除磷污泥颗粒化的机理,根据污泥特性、代谢产物和菌群结构的结果,推测不同表观气速下污泥颗粒化的机理,见图10.

图10 不同表观气速下除磷污泥颗粒化的机理Fig.10 Mechanism of the granulation of phosphorus removal sludge at different superficial gas velocity
在含有大分子有机物的复杂碳源条件下,较大的表观气速(R1,0.45cm/s)会抑制丝状菌的生长,缓慢生长微生物更容易富集(以GAOs 为主,PAOs 较少),这些具有储存功能的缓慢生长微生物通过分泌较多EPS 加快污泥颗粒化.较高的剪切力容易将颗粒边缘松散的污泥冲刷至液相,形成的颗粒污泥密实度更高,形状更为规整.被剪切下的小聚集体凝聚性能较好、密度更大,可保留在系统中再次颗粒化,实现颗粒污泥的良性再生.低表观气速下(R3, 0.15cm/s)也能形成好氧颗粒污泥,但形成速度略慢于高表观气速.具有水解发酵功能的丝状菌(Kouleothrix)被保留在系统中,这些丝状菌一方面形成颗粒污泥的骨架使菌胶团菌附着生长,另一方面为传统聚磷菌和传统聚糖菌等缓慢生长微生物提供小分子碳源,缓慢生长微生物再通过分泌大量 EPS 尤其是多糖类物质,与丝状菌共同促进污泥的颗粒化.同时,尽管R3 形成颗粒污泥的速度略慢于R1 但仍快于R2,降低表观气速不但能节省能耗还能强化大分子碳源的水解发酵,为同步硝化反硝化和生物除磷提供较好的环境,系统的脱氮除磷效率更高.在中等表观气速下(R2,0.30cm/s),剪切力和具有水解发酵功能丝状菌的作用都不明显,系统不易稳定,需要较长时间使具有胞内储存功能的菌群(如Candidatus_Competibacter和Amaricoccus等)与水解发酵菌群和谐共生实现颗粒化.考虑到实际应用,在降低表观气速培养好氧颗粒污泥的过程中,为避免中等表观气速下系统不稳定,需关注碳源代谢产物的浓度,避免过多碳源泄露至好氧区.此外,当颗粒污泥形成后,低表观气速可能导致部分大粒径污泥沉积于反应器底部而降低除磷率,因此需采取一定措施避免好氧阶段大颗粒的沉积,如增加水力循环或者机械搅拌等.
3 结论
3.1 复杂碳源条件下,表观气速的递减与污泥的颗粒化进程并不完全一致.R1 系统表观气速最大(0.45cm/s),颗粒污泥以水力选择为主,污泥颗粒化进程较快,110d 实现污泥颗粒化,形成的颗粒污泥密实程度高,性能更好;R3 系统表观气速最小(0.15cm/s),污泥颗粒化主要以代谢选择为主,190d 实现污泥颗粒化,进程慢于R1 但快于R2,颗粒污泥密实度最低,形状不规整,边界不清晰;R2 系统表观气速介于R1和R3 之间(0.30cm/s),菌群筛选介于水力选择和代谢选择之间,污泥颗粒化进程最慢,培养至200d 后开始颗粒化.
3.2 厌氧好氧交替运行使3组反应器LB-EPS二级结构中α-螺旋/(β-折叠+无规则卷曲)的数值均逐渐降低,LB-EPS 疏水性的不断升高可能使污泥颗粒化成为一个必然趋势;TB-EPS 中该数值依次升高,导致污泥密实度依次降低.降低表观气速有利于大分子有机物水解发酵为更多的VFAs 和乳酸等小分子有机物,污泥PHAs 的厌氧合成量更大,更易实现厌氧饱食好氧饥饿的碳源代谢模式.
3.3 16S rRNA 基因高通量测序分析显示,随着表观气速的降低,绿弯菌门(Chloroflexi)的丰度依次升高,分别为7.86%、8.77%和32.74%.在属水平,传统聚糖菌Candidatus_Competibacter是污泥颗粒化的核心菌群,R1 较高的表观气速有利于该菌的维持和优势增殖,污泥颗粒化进程最快.R3 表观气速最低,水解发酵功能菌群Kouleothrix(1851 型丝状菌,属于绿弯菌门)大量生长(丰度10.44%),通过提供丝状骨架和小分子碳源促进Candidatus_Competibacter的优势增殖(丰度25.73%),加快了污泥的颗粒化进程.
