A dynamic regulation of nitrogen on floret primordia development in wheat
2024-03-07ZhenZhngYujingLiYuxunWuXioyuZhengXioleiGuoWnSunZhenciSunZhiminWngYinghuZhng
Zhen Zhng, Yujing Li, Yuxun Wu, Xioyu Zheng, Xiolei Guo, Wn Sun, Zhenci Sun,c,Zhimin Wng,c,*, Yinghu Zhng,c,*
a College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China
b College of Biological Sciences, China Agricultural University, Beijing 100193, China
c Engineering Technology Research Center for Agriculture in Low Plain Areas, Cangzhou 061800, Hebei, China
Keywords:Fertile florets Floret primordia Grain number Meiotic ability Morphological development
ABSTRACT Nitrogen (N) fertilization is critical for spike and floret development, which affects the number of fertile florets per spike(NFFs).However,the physiological regulation of the floret development process by N fertilization is largely unknown.A high temporal-resolution investigation of floret primordia number and morphology, dry matter, and N availability was conducted under three N fertilization levels: 0 (N0),120 (N1) and 240 (N2) kg ha-1.Interestingly, fertile florets at anthesis stage were determined by those floret primordia with meiotic ability at booting stage: meiotic ability was a threshold that predicted whether a floret primordium became fertile or abortive florets.Because the developmental rate of the 4th floret primordium in the central spikelet was accelerated and then they acquired meiotic ability,the NFFs increased gradually as N application increased, but the increase range decreased under N2.There were no differences in spike N concentration among treatments, but leaf N concentration was increased in the N1 and N2 treatments.Correspondingly, dry matter accumulation and N content of the leaf and spike in the N1 and N2 treatments was increased as compared to N0.Clearly,optimal N fertilization increased leaf N availability and transport of assimilates to spikes,and allowed more floret primordia to acquire meiotic ability and become fertile florets, finally increasing NFFs.There was no difference in leaf N concentration between N1 and N2 treatment, whereas soil N concentration at 0–60 cm soil layers was higher in N2 than in N1 treatment,implying that there was still some N fertilization that remained unused.Therefore,improving the leaf’s ability to further use N fertilizer is vital for greater NFFs.
1.Introduction
In wheat(Triticum aestivum L.),grain number per spike is a critical component of grain yield; grains at the maturity stage are developed from fertile florets at the anthesis stage.Numerous studies have concentrated on increasing grain number, primarily by increasing the number of fertile florets per spike (NFFs).In wheat, a spike inflorescence produces many spikelets, each containing 7–12 floret primordia[1–3].In fact,not all floret primordia in a spike continue fertile development, and only a small part of them could become fertile florets.In a wheat spike, nine floret primordia that appear in the central spikelet, F5–F9 are aborted and only F1–F4 continue fertile development (Fig.1).During the evolutionary process, selective abortion is a survival strategy in which some floret primordia are sacrificed to allow the rest to survive.

Fig.1.Developmental process of all floret primordia at central spikelet in spike from the terminal spikelet stage to anthesis stage.
In previous studies [2–5], floret primordia development was described on the Waddington scale[4],and it was found that floret primordia start abortion when the first floret primordium at the central spikelet is in the W7 and W9 stages.When the F1 at the central spikelet develops from the W7 to the W9 stage, F5–F9 initiate abortion (Fig.1C, D).Indeed, floret primordia fate is determined mainly by the anthers, because the anther is more sensitive and vulnerable to stress than the ovary[6,7].The anther’s function in determining floret primordia fate remains unclear.The process of selective abortion is inevitable,but the number of abortive floret primordia depends on environment, growth position,and developmental sequence.Previous studies [3,8,9] have revealed that early-developing floret primordia(F1–F4,those most proximal to the rachis) generally develop into fertile florets,whereas late-developing floret primordia (F5–F9, the most distal florets) never become fertile florets (Fig.1).Thus, accelerating the developmental rates of the middle-developing floret primordia(F5, intermediate to the rachis) can promote them to become fertile florets, increasing NFFs [9–11].However, it is difficult to identify which floret primordia have the potential to be regulated to become fertile florets on the sole basis of their position or developmental sequence within a spike.There are 6–7 floret primordia in the basal spikelet, but none of them develop into fertile florets,including the early-developing floret primordia(F1–F2,most proximal to the rachis)[12].There must be a criterion or threshold that predicts or distinguishes between floret primordia with fertile or abortive potential before they become fertile or abortive florets.
Under field conditions, optimum N-fertilizer application increases NFFs, while excessive N-fertilizer application barely further increases NFFs [9,11,13].It is widely acknowledged [2,14,15]that the NFFs exhibit a positive relationship with the accumulation and partition of spike assimilates at the anthesis stage, and that improving spike assimilates increases NFFs.It appears that the increased NFFs under optimum N-fertilizer application is due mainly to the rapid accumulation of spike assimilates.Spike growth is affected by the growth of non-spike organs; the ability of the spike to compete with non-spike organs for assimilates is reduced under drought stress conditions[2,12];this is further supported by a detailed examination [16] of the effects of semidwarfing genes (Rht) that confirmed that a reduction in stem height increases NFFs by increasing spike assimilates during the pre-anthesis stage.However, a dynamic relationship between the spike and non-spike organs under diverse N-fertilization conditions remains unclear.N-fertilizer in the soil is divided into many types according to the N-containing groups, of which nitrate N(N-NO3-) is the main form absorbed and used by wheat.N-NO3-is distributed mainly in the 0–200 cm soil layer and in particular the 0–100 cm soil layer[17,18].It is unclear how the plant coordinates the uptake and utilization of N between aboveground and belowground organs under optimum N-fertilizer conditions to facilitate the transfer of assimilates from non-spike organs to the spike and increase NFFs.And why does more N fertilization fail to increase it further?
The purpose of this study was to characterize the response of floret primordial development to N fertilization by sampling floret primordia during their development and recording N concentrations in plant organs and soil.
2.Materials and methods
2.1.Winter wheat growth conditions
A field experiment was performed at the Wuqiao Experimental Station of China Agriculture University (37°41′02′′N, 116°37′23′′E),Hebei province, China during the 2018/2019, 2019/2020, and 2020/2021 wheat growing seasons.The winter wheat cultivar Shi-Mai 22 (a medium-spike type cultivar whose central spikelet carries the most grains and its basal spikelet the fewest), was sown on October 15, 2018, October 17, 2019, and October 15, 2020.Before sowing, soil available N, P, and K contents in the topsoil(0–40 cm) were respectively 1.02 g kg-1, 18.4 mg kg-1, and 120.8 mg kg-1in 2018,1.00 g kg-1,20.3 mg kg-1,and 108.6 mg kg-1in 2019,and 0.98 g kg-1,24.4 mg kg-1,and 100.7 mg kg-1in 2020.The rainfall from sowing date to maturity stage was 51 mm in 2018/2019, 158 mm in 2019/2020, and 106 mm in 2020/2021.The mean daily temperatures were 8.9 °C in 2018/2019, 9.1 °C in 2019/2020, and 8.8 °C in 2020/2021.
2.2.Experimental design
There were three N levels as described previously [9]; briefly,0 kg N ha-1(N0, no N fertilizer applied), 120 kg N ha-1(N1,120 kg N ha-1applied before sowing) and 240 kg N ha-1(N2,120 kg N ha-1applied before sowing and another 120 kg N ha-1at the terminal spikelet stage).The experiment was constructed as a split plot with three replicates, with each plot 9 m × 4.5 m.All plots were irrigated with 50 mm water before sowing and at the terminal spikelet stage to ensure emergence and avoid water stress during each growing season.Before sowing, 90 kg K2O ha-1and 120 kg P2O5ha-1were applied to the soil in all treatments.The planting density was 400 seeds m-2.
2.3.Sampling and measurement
Tissues were sampled every 3 days from 3 to 39 days after the terminal spikelet stage (DAT).At each sampling date, 10–15 main shoots were taken.For each main shoot, the numbers of spikelets and floret primordia per spikeket were recorded.The main shoot was then separated into spike,stem,leaf,and leaf sheath and dried to constant weight at 75°C to determine dry matter and N concentration and content.Dry matter partition (%) in organs was the ratio of their dry matter to main-shoot dry matter.For all samples,Kjeldahl digestion was used to measure N concentration.N content was calculated following Li et al.[19]:organ N content=organ dry matter × organ N concentration.The dynamic process of development of floret primordia number, organ dry matter, and organ N content was fitted with a logistic model following Fang et al.[20],where V1,V2,and V3 represented respective rates of increase during the gradual-growing, fast-growing, and slow-growing phases.Soil samples were extracted with 0.01 mol L-1CaCl2, and NO3--N concentration was measured with an ultraviolet spectrophotometer [21].The morphology of floret primordia was recorded mainly in 2020/2021.
A high temporal and spatial resolution analysis of the floret primordia number and morphology within a spike was performed to determine which primordia finally reached the stage of fertile floret.Changes in floret primordia morphology at the basal spikelet(1st spikelet), central spikelet (10th spikelet) and apical spikelet(19th spikelet) were recorded from from 3 to 39 days DAT.The morphology of floret primordia and cellular structure of the anther were obtained as described in Zhang et al.[22].
2.4.Statistical analysis
All data were analyzed by R (V 4.2.1) and RStudio.Statistical significance was identified using the Student’s t-test at the P < 0.05 (*), P < 0.01 (**), and P < 0.001 (***).
3.Results
3.1.Floret primordia developmental phases
NFFs was significantly increased by 14.5%in N1 and 18.1%in N2 treatments relative to the N0 treatment in all three growing seasons (Fig.S1).Moreover, during the three growing seasons, there was no difference in mean NFFs between the N1 and N2 treatments.These results suggested that N-fertilizer application in the N1 and N2 treatments promoted more floret primordia to become fertile florets,whereas greater N-fertilizer application in N2 did not continue to achieve a significant increase in the NFFs than N1.Changes in floret primordia morphology at the basal spikelet(first spikelet, Fig.S2), central spikelet (10th spikelet, Fig.2) and apical spikelet(19th spikelet,Fig.S3)can be divided into three key phases as shown in [22]: the differentiation phase (the number of floret primordia increases, and each shows normal morphology from 3 to 21 days after terminal spikelet stage (DAT)), the predimorphism phase (this phase occurs at 24 DAT and indicates the end of the differentiation phase and the start of the dimorphism phase, i.e.booting stage), and dimorphism phase (all floret primordia in the spike show dimorphic development: many floret primordia begin to abort/degenerate,whereas the remaining floret primordia continue to develop fertilely from 27 to 39 DAT).
3.2.Morphology of floret primordia during dimorphism phases
During the dimorphism phase, all floret primordia within a spike were classified into three types based on their morphology(Fig.2): I) ‘‘fertile floret primordia” with a fertile morphology(e.g.F1–F3 in N0 and F1–F4 in N1–N2 treatments); II) ‘‘shortlived floret primordia” with a normal morphology for a short time(e.g., F4–F5 in N0 and F5 in N1-N2 treatments at 27 DAT); and III)‘‘abortive floret primordia” with an abnormal or wilted morphology (e.g., F4–F6 in N0 and F5–F6 in N1-N2 treatments from 30 to 39 DAT).The ‘‘short-lived floret primordia” would become ‘‘abortive floret primordia”.The number of all floret primordia per spike increased rapidly from 3 to 15 DAT,then slowly from 15 to 24 DAT,and finally decreased from 27 to 39 DAT (Fig.S4A).At 12 DAT,plants in N0 carried more floret primordia per spike than those in N1 and N2;at 18 DAT,plants in N0 carried more floret primordia than those in N1.The number of all floret primordia per spike increased significantly in N2 at 21–24 DAT and in N1 and N2 at 36–39 DAT relative to the N0 treatment.The dynamics of floret primordia development over time were fitted with a nonlinear model(a logistic model),and the rates of increase in the number of floret primordia between the N treatments were compared.The number of all floret primordia per spike increased at similar rates in all N treatments during gradual-growing phase(V1),fast-growing phase(V2), and slow-growing phase (V3) (Fig.S5).
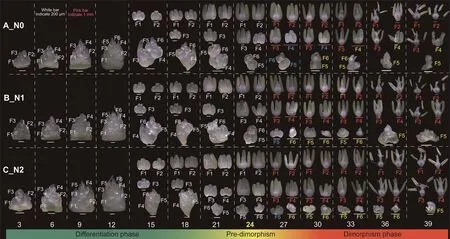
Fig.2.Changes in the number and morphology of all floret primordia in the central spikelet.
The number of‘‘fertile floret primordia”per spike maintained a steady level during the dimorphism phase,whereas the number of‘‘abortive floret primordia” per spike increased from 27 to 30 DAT and then decreased from 30 to 39 DAT, a behavior attributed mainly to rapid abortion of ‘‘short-lived floret primordia” from 27 to 30 DAT (Fig.S4B–D).The increased number of ‘‘fertile floret primordia” in the central spikelet (Fig.S6) resulted in a consistent increase in the total number of ‘‘fertile floret primordia” per spike in the N1 and N2 treatments relative to N0(Fig.S4B).At 30 and 39 DAT, the number of ‘‘abortive floret primordia” per spike was significantly higher in the N0 than in the N1 and N2 treatments(Fig.S4C).At 27 DAT, each treatment showed a similar number of ‘‘short-lived floret primordia” per spike (Fig.S4D).The results suggested that N-fertilizer application increased the number of‘‘fertile floret primordia” per spike and delayed the degeneration of ‘‘abortive floret primordia”, thereby increasing the number of all floret primordia per spike during the dimorphism phase.Although there were three types of floret primordia throughout the dimorphism phase, the floret primordia experienced only two fates: fertility or abortion.Ultimately, ‘‘short-lived floret primordia” and ‘‘abortive floret primordia” did not contribute to the increase in the number of ‘‘fertile floret primordia”.
3.3.Rates of floret differentiation at three N levels
To investigate the causes of the differences in the number of floret primordia per spike among N treatments, the floret primordia at each spikelet in a spike were counted.All spikelets were then divided into three parts based on their position in a spike: basal(spikelets 1–6), central (7–13), and apical (14–19) spikelets(Fig.3N).The number of all floret primordia at the apical spikelet was significantly lower in the N1 and N2 treatments than in the N0 treatment at 12–18 DAT (Fig.3D, F).The number of all floret primordia at the apical spikelet in the N0 treatment remained constant from 18 to 24 DAT (Fig.3F–H).The number of all floret primordia at the basal and central spikelet were increased in N1 and N2 treatments relative to the N0 treatment at 21 and 24 DAT(Fig.3G,H).These results implied that floret primordia differentiation in response to N supply occurred in the order of apical,middle, and basal spikelet.During the dimorphism phase, the number of all floret primordia at the basal and central spikelet was significantly higher in N1 and N2 treatments than in the N0 treatment from 30 to 33 DAT (Fig.3J, K).From 36 to 39 DAT, the number of all floret primordia at the apical, middle and basal spikelet was significantly lower in the N0 than in the N1 and N2 treatments (Fig.3L, M).
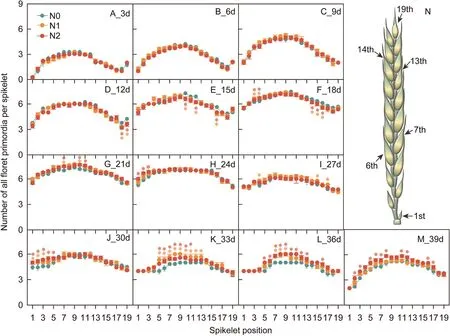
Fig.3.Changes in the number of all floret primordia per spikelet from 3 to 39 days after terminal spikelet.
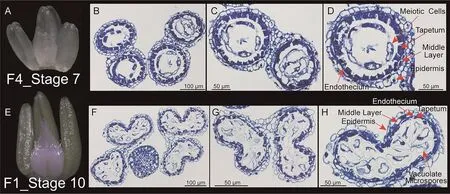
Fig.4.Morphology and anatomy of anther.
The number of all floret primordia per spikelet gradually increased until 21–24 DAT, and then decreased (Fig.3), indicating that each spikelet in a spike reached its maximal potential during 21–24 DAT (Fig.3G, H).The central spikelet had higher the maximum number of floret primordia than the basal and apical spikelets during 21–24 DAT (Fig.3G, H); correspondingly, the central spikelet had the highest number of fertile florets per spikelet at the anthesis stage(39 DAT,Fig.3M).Although the number of floret primordia in the 1st spikelet was lower from 3 to 6 DAT(Fig.3A,B),the 1st spikelet had a similar or even higher maximum number of floret primordia than the 19th spikelet (Fig.3G, H).This finding indicated that although the 1st spikelet differentiated floret primordia later than the 19th spikelet, the 1st spikelet had a faster differentiation rate within a shorter time.The number of fertile floret primordia was markedly higher in the 19th than in the 1st spikelet (Fig.S6).Thus, central spikelet had the highest maximum number of floret primordia and number of fertile florets;however,the maximum number of floret primordia in basal spikelet was greater than apical spikelet,and apical spikelet had a greater number of fertile florets than basal spikelet.The developmental duration of floret primordia was longest at the central spikelet and shortest at the basal spikelet.From 3 to 24 DAT, the 1st spikelet showed a faster increase rate of floret primordia number (from 0 to 6 floret primordia)than the 19th spikelet(from 2 to 6 floret primordia) (Fig.3A–H).Collectively, these results suggested that the developmental duration rather than the differentiation rate during differentiation phase and pre-dimorphism contributed to a high survival rate of floret primordia during the dimorphism phase.The central spikelet usually had the highest number of fertile florets mainly because those early-developing floret primordia had a sufficient developmental duration.Thus, the goal of increasing the number of fertile florets requires providing more time for floret primordia to develop into fertile florets.
3.4.Meiotic ability of floret primordia determined their fertile fate
To identify the difference between early- and late-developing floret primordia, the anthers of the earliest and latest developing floret primordia with fertile potential were used for cellular observation at the pre-dimorphism phase.In this study,F1 and F4 in the central spikelet in the N1 treatment represented respectively the earliest and latest floret primordium with fertile potential within a spike in all treatments (Figs.2, S2, S3).Transverse sections revealed that the anther of F4 and F1 was at the meiotic stage(stage 7, microspore mother cells begin to undergo meiosis as the tapetum continues to grow and vacuolate, Fig.4A–C) and vacuolate microspore stage (stage 10, microspores become vacuolate and pressed into tapetal wall Fig.4D, F), respectively, according to an anther developmental scale[23].The findings suggested that when the earliest developing floret primordia within a spike entered the vacuolate microspore stage, all floret primordia reached the pre-dimorphism phase.All floret primordia that could reach or pass the meiotic stage would continue to fertile development,whereas the other floret primordia that had not reached the meiotic stage would stop development and then gradually abort and degenerate in the subsequent dimorphism phase.In other words, meiotic ability was the threshold for distinguishing earlyfrom late-developing floret primordia and to determine whether or not floret primordia had fertile potential during the dimorphism phase.The meiotic ability occurred in parallel to morphological characteristics (e.g., the length of anthers was longer than that of the ovary; anthers turned green) during the pre-dimorphism phase.As expected,the number of meiotic floret primordia during the pre-dimorphism phase was similar to the number of ‘‘fertile floret primordia” per spike during the dimorphism phase: moreover, they were significantly higher in the central spikelet under the N1 and N2 treatments than under N0.The number of meiotic floret primordia was highest in the central spikelet,while the basal spikelet had the lowest (Fig.S6).These findings suggested that more floret primordia at the central spikelet acquiring meiotic ability during the pre-dimorphism phase were primarily responsible for the increase in NFFs after N-fertilizer application.In fact,accelerating floret primordia development was a stepwise process, and the performance of floret primordia at the pre-dimorphism phase was affected by floret primordia during the differentiation phase.These results suggested that accelerating the development of more floret primordia during differentiation phase to enter into meiosis at pre-dimorphism phase, leading them to become ‘‘fertile floret primordia” during the dimorphism phase, increased the number of fertile florets at the anthesis stage.
3.5.Primarily spike assimilates regulated early-developing floret primordia
Assimilate accumulation in the whole plant and its organs showed three patterns: whole plant (0.29–2.19 g) and stem(0.02–1.09 g) dry matter weight gradually increased as growth stage progressed (Fig.S7A, C); spike dry matter weight remained steady during early spike development (3–18 DAT) but rapidly increased during late spike development (21–39 DAT) (Fig.S7B);leaf and leaf sheath dry matter weight rapidly increased from 3 to 24 DAT and then remained steady (leaf and leaf sheath) during 27–39 DAT (Fig.S7D, E).Wheat organs showed four assimilatepartition patterns: spike dry matter partition (0.40%) remained steady during early spike development (3–18 DAT) and then rapidly increased (1.72%–14.04%) during late spike development(21–39 DAT) (Fig.S7F); leaf sheath dry matter partition remained steady (27.47%) from 3 to 24 DAT and then gradually decreased(28.24%–18.53%)from 27 to 39 DAT(Fig.S7I);stem dry matter partition increased(6.52%–49.48%)gradually with growth days;however, a decreasing trend(69.10%–17.96%) was observed in leaf dry matter partition (Fig.S7G, H).Thus, the leaf was the important organ for assimilate accumulation and partition during the differentiation and pre-dimorphism phases, followed by leaf sheath,stem,and spike.During the dimorphism phase,assimilate accumulation and partition among organs followed the order stem > leaf sheath > leaf > spike.
Plant(at 3, 9, 12,18,21,27,36,39,and DAT) and stem(at 3, 9,15, 18, 21, 36, 39, and DAT) dry matter weight was significantly greater in N1 and N2 treatments than in the N0 treatment(Fig.S7A,C).Spike dry matter weight did not differ among N treatments at 3–12 DAT,but spike dry weight was significantly greater in N1 and N2 than N0 treatment during 15–18, 21–24 and 33–39 DAT (Fig.S7B).Compared to N0, stem dry matter weight in N1 and N2 treatments was significantly increased during 3, 9, 15–21 and 36–39 DAT(Fig.S7C).Compared to N0,leaf dry matter weight in N1 and N2 treatments was significantly increased from 3 to 39 DAT, exception at 33 DAT (Fig.S7D).Significant difference among N treatments in leaf sheath dry matter weight was observed only at 3, 9 and 39 DAT (Fig.S7E).These results suggested that the leaf was the organ responding most strongly to N-fertilizer treatment and the leaf sheath responding most weakly.The logistic model revealed that the V2 of plant dry matter in N1 and N2 was significantly increased relative to N0 (Fig.S8A).The V2 and V3 of spike dry matter were significantly greater in N2 than in N0 and N1 treatments(Fig.S8B).The V1,V2, and V3 of stem dry matter were significantly greater in N1 and N2 than in N0 (Fig.S8C).The V1 of leaf dry matter was significantly greater in N0 and N2 than in N1,and the V2 of leaf dry matter was significantly greater in N1 and N2 than in N0(Fig.S8D).There was no significant difference among N treatments in V1,V2 and V3 of leaf sheath(Fig.S8E).Thus,increasing the rates of assimilate accumulation in plants during the fastgrowing phase, in spikes during the fast-growing and slowgrowing phases, in stems during every growing phase, and in leaves during the fast-growing phase, increased their assimilate accumulation.In comparison to N1 and N2 treatments,dry matter partition in the N0 treatment was significantly higher and occurred mainly during early leaf growth,and middle and late spike growth(Fig.S7F, H).Compared to N0, leaf sheath dry matter partition in N1 and N2 treatments was significantly decreased (Fig.S7I).Only a few significant differences were observed between N1 and N2 treatments in organ dry matter weight and partition (Fig.S7),implying that compared to N1, further increase of N fertilization negligibly increased dry matter weight and partition.
The number of all floret primordia per spike was not always closely associated with spike dry matter weight (Fig.S9).A weakly positive association between the number of all floret primordia per spike and spike’ dry matter was observed from 6 to 27 DAT,and in particular from 15 to 24 DAT (Fig.S9A–H), in parallel with late floret primordia differentiation.Thus,despite the rapid assimilate accumulation,the number of late-developing floret primordia did not respond to assimilate availability.A positive correlation between the number of all floret primordia per spike and spike dry matter weight was observed from 27 to 36 DAT (Fig.S9I–L),which occurred in parallel to the development of those earlydeveloping floret primordia.Although the number of‘‘fertile floret primordia” was significantly and highly correlated with the spike dry matter weight(Fig.S10),it showed a weak or negative correlation with the number of ‘‘abortive floret primordia” and ‘‘shortlived floret primordia”from 27 to 39 DAT(Fig.S11).These observations suggested that the increase in assimilate accumulation increased the number of ‘‘fertile floret primordia” and delayed the degeneration of ‘‘abortive floret primordia” during the dimorphism phase.Although early-developing floret primordia responded rapidly and positively to assimilate accumulation, the differentiation or abortion of those late-developing floret primordia was weakly regulated by assimilates.
3.6.N availability above and below ground
N availability changes assimilate supply to spike, indirectly affecting the number of floret primordia.N concentration in all organs decreased gradually (Fig.S12A–D), whereas spike, stem,and leaf sheath N content increased with time(Fig.S12E–H).Spike N concentration was not affected by N-fertilizer application,but N application significantly increased leaf N concentration (Fig.S12A,C).Stem and leaf sheath N concentration was significantly higher in N1 and N2 treatments from 3 to 12 DAT (Fig.S12B, D).These results suggested that N-fertilizer application increased mainly stem and leaf sheath N concentration in the early differentiation phase, but consistently increased N concentration in leaf from the differentiation to dimorphism phases.Compared to N0 treatment,spike, leaf, and leaf sheath N content significantly increased from 3 to 39 DAT in N1 and N2 treatments(Fig.S12E,G,H).The difference between N1 or N2 and N0 treatment in stem N content was found only at 27, 36 and 39 DAT (Fig.S12F).These results implied that N-fertilizer application increased mainly spike, leaf and leaf sheath N accumulation.The logistic model revealed that V1, V2,and V3 of spike N content in N2 were significantly greater than N0 (Fig.S13A); V1 of stem N content was significantly greater in N2 than in N0 and N1 (Fig.S13B); V1 of leaf sheath N content was significantly higher in N1 and N2 than in N0(Fig.S13C).Thus,the increases in the rates of N accumulation in spike during each growing phase and in leaf sheath during the fast-growing phase contributed to a high level of N accumulation in the plant.Spike and stem N partition increased gradually, while leaf N partition decreased over time (Fig.S12I–K).The stem N partition did not change between treatments (Fig.S12J).Compared to N0, spike N partition in N1 and N2 treatments during 33–36 DAT was increased (Fig.S12I).Leaf N partition was lower from 3 to 21 DAT but higher from 24 to 39 DAT in N1–N2 than that in N0 treatment,whereas leaf sheath N partition was decreased from 24 to 39 DAT (Fig.S12K–L).These results suggested that N-fertilizer application increased mainly N distribution to spike and leaf, but reduced distribution to leaf sheaths during the dimorphism phase.Thus, the leaf was the organ most responsive to N-fertilizer application and the stem the least responsive.
The uptake and utilization of N by plants is dependent mainly on NO3--N availability in the soil.In the 20 cm soil layer, soil NO3--N concentration in N1 increased significantly relative to N0 treatment at the terminal spikelet stage during the 2018/2021 growing seasons(Fig.5A-C).Three days after irrigation at terminal spikelet stage,soil NO3--N concentration in the N1 treatment in the 0–100 cm soil layer was significantly increased relative to N0 during the 2019–2021 growth seasons (Fig.5E, F).These results suggested that the N applied before sowing was stored mainly in the upper soil layer and that the N was carried after irrigation from the 20 cm to the 40–100 cm soil layer.Three days after Nfertilizer application and irrigation at terminal spikelet stage, soil NO-3-N concentration increased in the 0–100 cm soil layer in the N2 treatment relative to N1 during the 2018/2021 growing seasons(Fig.5D,F),indicating that N-fertilizer applied at the terminal spikelet stage was also distributed in the 0–100 cm soil layer.At the anthesis stage, soil NO3--N concentration at 0–140 cm in 2019/2020 and 80–100 cm soil layer in 2020/2021 was significantly higher in the N1 than in the N0 treatment,whereas soil NO3--N concentration at 0–40 and 120–160 cm in 2018/2019, 140–160 cm in 2019/2020, 0–60 and 120–140 cm soil layer in 2020/2021 was significantly higher in the N2 than in the N0 treatment.Soil NO3--N concentration at 0–40 and 120–140 cm in 2018/2019, 0–10 cm in 2019/2020 and in the 0–60 cm soil layer in 2020/2021 in the N2 treatment was significantly higher than in the N1 treatment(Fig.5G–I).These results suggested that N-fertilizer applied before sowing was absorbed and used by wheat mainly during the terminal spikelet to anthesis stages,whereas the part of N fertilizer applied at the terminal spikelet stage was left mainly in the 0–60 cm soil layer and was not absorbed and used.

Fig.5.Changes in soil N concentration in the 20 to 200 cm soil layers.
Distribution of NO3--N in soil layers was affected mainly by irrigation water, leading to a soil NO3--N concentration difference among treatments.At the terminal spikelet stage, soil water content gradually increased from the 0- to 200 cm soil layers(Fig.S14A–C), whereas no difference in soil water content among N treatments was observed in any soil layer.Compared to 0–100 cm soil layers, soil water content increased in each layer within 0–80 cm at three days after irrigation at terminal spikelet stage (Fig.S14A–F).These results suggested that irrigated water remained mainly in the 0–80 cm soil layer and did not move below 100 cm.Soil water content only at 0–20 cm in N2 in 2018/2019 and in N1 and N2 treatments in 2019/2021 was significantly higher than in the N0 treatment.Compared to three days after irrigation at terminal spikelet stage, soil water content at 0–80 cm soil layer rapidly decreased at the anthesis stage, particularly in the top soil layer (Fig.S14G–I), implying that soil water at 0–80 cm soil layer was used by wheat after irrigation.At the anthesis stage,soil water content in N1 and N2 at 0–60 cm in 2018/2019 and 2020/2021 and at 0–100 cm in 2019/2020 was significantly higher than in N0 treatment.
4.Discussion
4.1.Floret primordia initiate abortion because they lack meiotic ability
The challenge for increasing the number of floret primordia is identifying the dynamic changes of each floret primordium development in a spike.In this study,all floret primordia within a spike developed normally during differentiation (3–21 DAT) and pre-dimorphism phase (24 DAT), and then experienced dimorphic development (fertility and abortion) during dimorphism phase(27–39 DAT)(Figs.2, S2,S3).During dimorphism phase,only‘‘fertile floret primordia”developed into fertile florets,whereas‘‘shortlived floret primordia”and‘‘abortive floret primordia”degenerated and disappeared gradually.Compared to N1 and N2,the number of‘‘abortive floret primordia” per spike in N0 treatment was higher(Fig.S4C), and spike dry matter weight was higher in N1 and N2 than in N0 treatment (Fig.S7B).These results suggest that Nfertilizer application increased the availability of assimilates in the spike,delaying the degeneration or disappearance of‘‘abortive floret primordia”.In fact, these ‘‘abortive floret primordia” did degenerate or disappear (Fig.2) and did not contribute to an increase in NFFs.Thus,although the process of‘‘abortive floret primordia” degeneration or disappearance could be delayed by increase of assimilates, the ‘‘abortive floret primordia” finally still degenerate,and this process is a developmentally determined process.This inference is agreement with the theory[9,18,24,25]that it is difficult to reverse the developmental progress of floret primordia once they have reached a certain stage, because cell division has stopped and programmed cell death has begun.
It has been debated [2,5,26,27] whether abortive initiation of floret primordia is due to competition for assimilates or is developmentally pre-determined.Examination of floret primordia development over time revealed that floret primordia fate during dimorphism phase was determined at the pre-dimorphism phase(Fig.2).Cellular observation revealed that all floret primordia that could reach or exceed the meiotic stage at pre-dimorphism phase would continue to develop into ‘‘fertile floret primordia”, whereas other floret primordia that had not reached the meiotic stage would stop development and then become‘‘abortive floret primordia” and ‘‘short-lived floret primordia” in the subsequent dimorphism phase.The present study suggests that meiotic ability is the threshold or criterion determining whether or not floret primordia at pre-dimorphism phase have fertile potential.Although N fertilizer increased assimilate availability in the spike, the abortion of floret primordia at basal spikelet in all N treatments initiated rapidly (Fig.S2).Among all floret primordia without meiotic ability, even though F1–F3 had higher assimilates than F6–F7 in the basal spikelet, they all initiated abortion.Clearly, in floret primordia without meiotic ability, the initiation of floret primordia abortion may be attributed to developmental cues instead of assimilates.The number of floret primordia with meiotic ability in N1 and N2 was higher than in the N0 treatment at predimorphism phase(Fig.S6A),implying that increasing spike assimilates increased the number of floret primordia that acquired meiotic ability.The F4 in the central spikelet was aborted in N0,whereas it acquired meiotic ability and developed into fertile florets in N1 and N2 treatments (Fig.2), indicating that assimilates were critical in determining whether floret primordia acquired meiotic ability.The effect of assimilates and meiotic ability on floret primordia may be summarized as follows: a) all floret primordia without meiotic ability initiate abortion in response to a developmental cue and not to assimilates, and b) whether floret primordia can acquire meiotic ability is strongly influenced by assimilates.
4.2.The number of fertile florets increased as the duration or rate of floret primordia development were extended or accelerated
The maximum number of floret primordia does not accurately reflect the number of fertile florets.Although in a previous study[28], differences in the number of fertile florets were partially explained by the maximum number of floret primordia, the relationship varies with genotype, sowing date, and temperature[15,29,30].In the present study, the central spikelet carried the maximum number of floret primordia and consequently of fertile florets.Although the maximum number of floret primordia in the basal and apical spikelets was identical, the number of fertile florets in the apical spikelet was higher than that in the basal spikelet(Figs.S2,S3,S6).The reason is that although the basal spikelet had a faster differentiation rate and reached its maximum potential sooner than the apical spikelet, the floret primordia in the basal spikelet had little time to acquire meiotic ability (Fig.3A–H).The apical spikelet differentiated its maximum number of floret primordia earlier than the basal spikelet, giving it enough time to acquire meiotic ability, in agreement with a previous finding[22].These results suggest that extending the developmental duration of floret primordia instead of the maximum number of floret primordia determines the number of fertile florets.Accordingly,the best strategy for increasing the number of fertile florets would be to increase the number of high-quality floret primordia (those acquiring meiotic ability).Developmental rates of floret primordia in the central spikelet were markedly faster in N1 and N2 treatments than in N0 treatment(Fig.2),contributing to the acquisition of meiotic ability by more floret primordia in the central than in the basal spikelet.This finding implies that accelerating the developmental rate of floret primordia would be another way to increase floret primordia fertility.Differences among cultivars in the number of fertile florets have been attributed[9,10]to differing developmental rates of floret primordia.However, other studies[27,31] have found that accelerated development of primordia leads to more rapid degradation of these florets.Thus, the effect of developmental rate on floret fertility has been disputed.We expected that NFF would increase with the developmental rates of floret primordia, but this increase also accelerated the abortion of floret primordia lacking meiotic ability.In other words, rapid developmental rates helped those early-developing floret primordia acquire meiotic ability and then develop preferentially into fertile florets.It also helped those late-developing floret primordia develop,but they did not acquire meiotic ability and finally preferentially initiated abortion.The developmental rate of F4 in the central spikelet in N1-N2 was faster than that in N0 treatment(Fig.2);correspondingly,F4 in N1 and N2 acquired meiotic ability and then became a fertile floret, whereas F4 in N0 aborted.Thus, compared to N0,the central spikelet in N1 and N2 hosted approximately one more fertile floret(Fig.S6).Although the developmental rate of F5(late-developing floret primordia)in the central spikelet in N1–N2 was faster than in N0,F5 in each treatment did not acquire meiotic ability at the pre-dimorphism phase; F5 in N1 and N2 initiated abortion earlier than in N0 at 27 DAT(Fig.2).The number of abortive floret primordia per spike was increased in N0 relative to N1 and N2 treatments (Fig.S4C).Thus, to increase the number of fertile florets, floret primordia must acquire meiotic ability by an acceleration of their developmental rate, an extension of their developmental time, or both.
4.3.Increase in the number of fertile florets is limited by the leaf’s ability to use N
N influences wheat growth and assimilate accumulation[32,33].The gradual decrease in N concentration in each organ over the developmental times (Fig.S12A–H) and the increase in dry matter weight in each organ(Fig.S7B–E),indicate that N was used by the plant to promote assimilate accumulation.The finding of little difference among N treatments in spike N concentration(Fig.S12A), whereas spike dry matter weight was higher in the N1 and N2 treatments than in the N0 treatment(Fig.S7B),suggests that the increase of assimilate accumulation in the spike was due to transport from non-spike organs rather than synthesis in the spike.The finding that leaf N concentration and dry matter weight were higher in N1 than in N0 (Figs.S7D, S12C), indicates that a rapid increase in leaf N concentration promoted the synthesis of assimilates.These results suggest that the increase in assimilate accumulation in the spike may be attributed primarily to assimilates transported by leaves rather than to spike N.This suggestion agrees with previous reports[34,35]that wheat leaves are the key source organs of assimilate supply.Thus, the leaf was the major N sink and the source of assimilate supply.There were few differences between N1 and N2 treatments in organ dry matter or in N concentration and content (Figs.S7, S12), implying that optimal N-fertilizer application (N1) increased the translocation of assimilates from leaf to spike, whereas excess N-fertilizer application(N2) hardly further increased leaf N concentration or assimilate availability.In established populations,leaf development is limited by population architecture[36].Three days after N-fertilizer application,soil NO3--N content in the 20–100 cm soil layer was higher in N1 and N2 than in N0(Fig.5E,F);moreover,soil NO3–-N content was also higher in N2 than in N1 (Fig.5D, F).At anthesis stage, a difference in NO3--N content between N0 and N1 treatments was found only during 2019/2020, whereas this content was higher in the 20–60 cm soil layer under N2 than under the N0 and N1 treatments during 2018/2021(Fig.5G,I).These results suggest that optimal N fertilizer was rapidly absorbed and used by wheat from terminal spikelet stage to anthesis stage, whereas part of N fertilizer in N2 remained mainly in the 0–60 cm soil layer.Indeed,application of N fertilizer has a threshold effect on crop growth;excessive N-fertilizer application increases leaching and runoff loss, impairing wheat growth [37,38].
We propose a new model for the physiological regulation pathway, in which optimal N-fertilizer application promotes more floret primordia development into fertile florets, whereas high N fertilization does not further increase the number of fertile florets(Fig.6).Optimal N fertilizer in the main form of NO3--N is carried by irrigation water to the 0–100 cm soil layer, from which it is absorbed and used by the leaf to increase N concentration,accelerating assimilate accumulation.Part of these assimilates are then transported into the spike to improve N and assimilate accumulation,which allows more distal floret primordia in the central spikelet to develop and acquire meiotic ability, leading finally to an increased number of fertile florets.Under high N application, the extra N-fertilizer is not further used by leaves and remains mainly in the 0–60 cm soil layer,so that the number of fertile florets does not further increase.
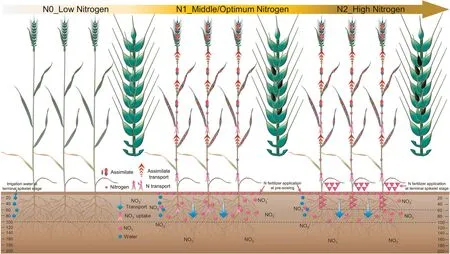
Fig.6.Schematic diagram of synergetic use of aboveground and underground N in wheat to increase the number of fertile florets.
CRediT authorship contribution statement
Zhen Zhang:Writing – original draft, Supervision.Yujing Li:Writing–review&editing.Yuxun Wu:Writing–review&editing.Xiaoyu Zheng:Writing–review&editing.Xiaolei Guo:Writing–review & editing.Wan Sun:Writing – original draft, Supervision.Zhencai Sun:Writing—original draft,Supervision.Zhimin Wang:Writing – original draft, Supervision.Yinghua Zhang:Writing –original draft, Supervision.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This study was supported by the National Key Research and Development Program of China (2022YFD1900703,2022YFD2300802), the Earmarked Fund for CARS (CARS-3), the National Natural Science Foundation of China (31871563), and China Postdoctoral Science Foundation(2022M723437).We thank Profs.Qingwu Xue(from Texas A&M AgriLife Research of USA)and James C.Nelson(Kansas State University)for their careful language revisions on this manuscript.We also thank the editor, and the anonymous reviewers for their valuable suggestions that improved the manuscript.
Appendix A.Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2023.10.009.
杂志排行

The Crop Journal的其它文章
- Flowering-time regulation by the circadian clock: From Arabidopsis to crops
- Global characterization of OsPIP aquaporins reveals that the H2O2 transporter OsPIP2;6 increases resistance to rice blast
- Drought-triggered repression of miR166 promotes drought tolerance in soybean
- The OsBSK1-2-MAPK module regulates blast resistance in rice
- Natural variation of an autophagy-family gene among rice subspecies affects grain size and weight
- Rice gene OsUGT75A regulates seedling emergence under deep-sowing conditions