circMYO9A_006通过翻译蛋白MYO9A-208aa发挥抑制心肌细胞肥大的作用*
2024-03-05姜佳雪苏金凤徐金东刘宇鹏方咸宏单志新
姜佳雪, 苏金凤, 王 娅, 欧 涛, 李 晖, 徐金东,刘宇鹏, 方咸宏, 单志新,5△
[1华南理工大学医学院,广东 广州 510006;2南方医科大学附属广东省人民医院(广东省医学科学院)检验科,广东 广州 510080;3南方医科大学附属广东省人民医院(广东省医学科学院)麻醉科,广东 广州 510080;4南方医科大学附属广东省人民医院(广东省医学科学院)心内科,广东 广州 510080;5南方医科大学附属广东省人民医院(广东省医学科学院)医学研究部,广东 广州 510080]
心血管疾病已成为全世界范围内人口死亡的主要原因之一[1]。正常心脏在受到外界刺激后,会发生生理性或病理性心肌肥大,生理性肥大通常发生在运动训练后、心脏病早期、妊娠末期或甲状腺功能亢进时,是心脏发生的一种良性的适应性反应[2];但在长期的高血压或超负荷血流灌注刺激下,这种生理性适应会演变成病理性心肌肥大,并进一步引发心力衰竭[3-4]。有研究证实,包括非编码RNA 在内的多种表观遗传方式在多种心血管疾病的发生中发挥重要的调节作用[5-6]。
环状RNA(circular RNA, circRNA)为单链闭合的非编码RNA 分子,在病毒、原核生物、单细胞真核生物和哺乳动物(也包括人类基因转录组)中均有表达[7]。多数circRNA 来源于其宿主基因的外显子序列,可通过不同的剪接环化方式形成[8]。与线性mRNA 分子相比,circRNA 转录效率较低,但却有更好的RNA 稳定性。circRNA 可做为微小RNA 或某些RNA 结合蛋白的分子海绵,对其起到特异性吸附作用[9-10],而一些circRNA 具有翻译蛋白/多肽的能力,并通过其翻译产物来发挥相应的生物学功能[11]。
circMYO9A_006 是由其宿主基因肌球蛋白IXA(myosin IXA,MYO9A)的第2~4 个外显子序列,通过反向剪接形成的circRNA,全长为1 126 nt。我们前期的circRNA 表达谱分析结果显示,circMYO9A_006在心衰患者心肌标本中表达显著升高[12],但其对心肌细胞肥大表型的调控作用尚不清楚。序列分析结果提示,该circRNA 包含1 个可能的开放阅读框(open reading frame, ORF)及2 个内部核糖体进入位点(internal ribosome entry site, IRES)。本项工作拟研究circMYO9A_006 对原代分离培养的大鼠和小鼠心肌细胞肥大表型的调节作用,并明确其是否具有翻译蛋白作用,并通过翻译的蛋白来发挥对心肌细胞肥大表型的调节作用。
“移就”又称“转借”、“移状”、“移用”,就是当甲乙两事物连在一起时,把原来属于甲事物的性状词语移用到乙事物上的一种修辞格。
农户是使用机械设备进行农业生产的主要力量,在农机深松整地技术推广过程中,首先需要面对的就是广大农户群众,农机深松整地技术推广需要农户群众的积极配合。因此,在推广过程中必须要加强对农户的教育,引导农户改变传统的思想观念,让农户认识到农机深松整地技术的重要性;学习专业的设备操作技巧,从而在日常生产过程中加强对机械设备的利用,提高生产水平[3]。
材 料 和 方 法
1 实验动物
本研究所使用的实验动物有出生1~3 d的SPF级C57BL/6 乳小鼠和Sprague-Dawley (SD)乳大鼠,雌雄不限,分别用于分离原代乳小鼠心室肌细胞(neonatal mouse ventricular cardiomyocytes, NMVCs)和乳大鼠心室肌细胞(neonatal rat ventricular cardiomyocytes,NRVCs)。以上实验动物均购于广州省医学实验动物中心[生产许可证号为SCXK(粤)2022-0002]。
2 细胞系和主要试剂
1.1 研究对象 回顾性分析2016年10月-2018年7月来我院(萍乡市胎儿畸形产前筛查中心)行III级产前超声检查孕妇23835例,经产后超声心动图及引产后胎儿尸解证明上腔静脉先天性变异的72例患者。
抗体均在5%脱脂乳充分溶解后用于Western blot:鼠抗Flag-M2 型抗体(F3165,稀释比例1∶1 000)购自Sigma;兔抗β-肌球蛋白重链(β-myosin heavy chain, β-MHC)抗体(A7564,稀释比例1∶3 000)和兔抗骨骼肌肌动蛋白α1(skeletal muscle actin alpha 1,ACTA1)抗体(ab197345,稀释比例1∶5 000)购自ABclonal;兔抗心房钠尿肽(atrial natriuretic peptide,ANP)抗体(19H19L1,稀释比例1∶1 000)购自Invitrogen。
2.1.9 锰。2012年全市叶片锰平均含量为167.40 mg/kg(表1),说明锰的含量在较高的水平,这与果园较低的土壤pH和土壤透气性不良有关。果树锰含量过高易产生毒性,表现为粗皮病,韧皮部坏死枯斑,通常土壤pH 5.0以下尤为明显。高的土壤 pH(>6.3)易导致锰缺乏,相反,低的 pH(<5.6)会导致锰过剩。土壤 pH 4.5提高到pH 6.5,土壤锰的有效性降低30 ~50倍。如果锰缺乏,在落花后10~14 d喷布一次硫酸锰(浓度约4 g/L)即可。
Trizol Reagent 购 自Invitrogen;5× PrimeScript ™RT Master Mix 和预混型qPCR 试剂盒购自TaKaRa;胶原酶粉末购自Sigma;Dual-Luciferase®Reporter Assay System 购自Promega;鬼笔环肽(phalloidin)购自翌圣生物科技有限公司;BCA 蛋白定量试剂盒和蛋白Marker购自Thermo;PVDF 膜和Immobilon ECL Ultra Western HRP 试剂购自Millipore;超敏发光液购自广州创伟生物科技有限公司;脱脂奶粉购自Bio-Rad。
3 主要实验方法
所有数据的统计均用GraphPad Prism 9.5 进行分析,表示为三个独立实验的均数±标准差(mean±SD)。多组间均数比较用采用单因素方差分析,进一步两两比较采用Bonferroni校正的t检验。以P<0.05为差异有统计学意义。
资金来源渠道的单一严重限制了我国互联网汽车金融行业的发展,互联网汽车金融企业要运用多种方式筹集资金,拓宽筹资渠道。比如发行金融债券,向同业机构进行拆借等方式。这可以有效地克服先前资金来源期限短、金额少等缺点,促进互联网汽车金融行业的发展。
3.5 双萤光素酶报告基因实验 通过DNA 合成出circMYO9A_006 的2 个可能的IRES 序列,并定向克隆到pGL3-Promoter 萤光素酶载体中。将1 μg 海肾萤光素酶质粒(作为内部对照)与构建好的萤光素酶质粒共转染293T 细胞。转染后24 h 后,用购自Promega的双萤光素酶报告基因测定试剂盒中的裂解缓冲液裂解细胞,按照试剂盒指示测量萤火虫萤光素酶和海肾萤光素酶活性。相对萤光素酶活性即为被海肾萤光素酶标准化的萤火虫萤光素酶的值。
3.4 鬼笔环肽染色 参照我们已报道的方法[13],预先在激光共聚焦细胞培养皿中铺适当鼠尾胶原,使胶原均匀盖满皿底。于37 ℃细胞培养箱孵育1 h 以上,吸出胶原,将适量密度的乳小鼠心肌细胞悬液接种到皿中,置于细胞培养箱中培养。次日,用高压灭菌后的PBS 反复冲洗细胞,弃去废液。每皿加入1 mL 含2%血清的F12 培养液,将细胞同步化处理12 h。对照组细胞以带有绿色荧光蛋白(green fluorescent protein, GFP)的空载腺病毒感染,实验组细胞分别感染含不同目的基因的重组腺病毒,每组均做3个复孔处理。充分感染24 h 后弃原有细胞培养液,用PBS 漂洗细胞3 次。加入4%多聚甲醛室温静置固定20 min。回收多聚甲醛,PBS 洗涤细胞3 次(每次10 min),用提前2 h 配好的0.1% Triton X-100 透化细胞。弃透化液,加入500 μL 用3%牛血清白蛋白溶液配好的鬼笔环肽工作液(iFluor™ 647 标记的鬼笔环肽)避光孵育90 min。回收工作液,用预先配好的PBST 漂洗细胞3 次,每次10 min。加入3~5 滴DAPI 染色剂染色15 min 后,用PBS 漂洗5 min 后即可用于共聚焦显微镜拍照。使用ImageJ 软件进行细胞表面积的测量。
3.3 Western blot 事先用PBS 轻轻漂洗掉孔板内原有的细胞培养液,加入适量的RIPA 裂解液置于冰上充分裂解15 min。事先预冷离心机至4 ℃,将细胞裂解液于12 000×g离心15 min。取上清液,按照说明书指示,利用标准曲线法检测蛋白浓度。接着加入适量4×蛋白上样缓冲液于99 ℃变性10 min。利用SDS-PAGE 分离样品中不同分子量大小的蛋白,并将其转印到PVDF 膜上。将含有转印蛋白的PVDF膜在室温下用封闭液(含0.1% Tween 20和5%脱脂奶粉的TBST 缓冲液)封闭1 h 后,加入相应的Ⅰ抗稀释液,4 ℃过夜孵育。次日用TBST 缓冲液漂洗膜3次,每次6 min。用相应的Ⅱ抗稀释液,于室温孵育1 h 后,TBST 缓冲液漂洗膜3 次,每次10 min。使用LAS 500进行化学发光,后续用ImageJ软件进行灰度值分析。
3.2 RT-qPCR 使用Trizol 试剂从培养的细胞中提取总RNA,按照PrimeScript™ Master Mix 逆转录试剂说明书指示,将总RNA 逆转录为cDNA。使用2×SYBR Green Mix 和相应引物进行RT-qPCR 检测,用2-ΔΔCt公式计算相对基因表达水平。circMYO9A_006的上游引物序列为5'-AGGAGCAAGTGAAGATGAGAGA-3',下游引物序列为5'-TTCAAAGCGTCGTCTTCCTC-3',内参照GAPDH 的上游引物序列为5'-CAAGAAGGTGGTGAAGCAGG-3',下游引物序列为5'-CCACCCTGTTGCTGTAGCC-3'。PCR 条 件 如下:一个初始变性周期(95 ℃ 3 min),40 个扩增循环(95 ℃ 10 s,60 ℃ 20 s,72 ℃ 15 s),熔解曲线分析(95 ℃ 5 s,65 ℃ 1 min,97 ℃连续),冷却期(40 ℃30 s)。
4 统计学处理
3.1 分离和培养乳鼠心肌细胞 采用课题组以往做法分离并培养乳鼠心肌细胞[12]。取1~3 日龄的C57BL/6乳小鼠或SD 乳大鼠,拉紧其背颈部皮肤,持眼科剪沿腋下横向剪开,取出心脏置于提前预冷的PBS 中,将心脏周围血管和结缔组织用镊子轻轻剥离,置于7 mL 胰蛋白酶(0.1% trypsin-EDTA)中,4 ℃摇床上消化过夜。次日,加入14 mL 完全培养液并置于37 ℃水浴锅中孵育10 min终止消化。弃去试管中原有液体,加入10 mL事先用F12培养液配好的胶原酶溶液,37 ℃孵育10 min 后用巴氏管不断吹打直至吹碎整个心脏,用70 μm 细胞滤网过滤后将细胞悬液置于细胞培养瓶中,于37 ℃、5% CO2培养箱中培养,约1.5 h 后部分细胞贴壁,吸出上层细胞悬液即为乳鼠心肌细胞,置于已提前铺好鼠尾胶原的细胞板中。用含10% FBS 的完全培养液培养,使细胞稳定生长24 h后进行后续实验。
结 果
1 过表达circMYO9A_006 可抑制NMVCs 中心肌肥大相关蛋白表达
circMYO9A_006来源于其宿主基因MYO9A的第2~4 个外显子反向剪接环化形成,由1 226 个碱基组成(图1A)。通过分子克隆技术构建rAd-circ-MYO9A_006 重组腺病毒,其载体本身带有GFP 标签,用其感染NMVCs 24 h 后,观察到GFP 荧光充分表达;RT-qPCR 结果显示腺病毒可介导circMYO9A_006 在NMVCs 中有效表达(图1B)。DNA 测序结果证实,在NMVCs 中过表达的circMYO9A_006 包含正确的接头序列,见图1C。Western blot 结果显示,与感染空载体重组腺病毒组相比,过表达circMYO9A_006 可有效抑制NMVCs 中肥大相关蛋白β-MHC、ACTA1和ANP的表达(P<0.01),见图1D。
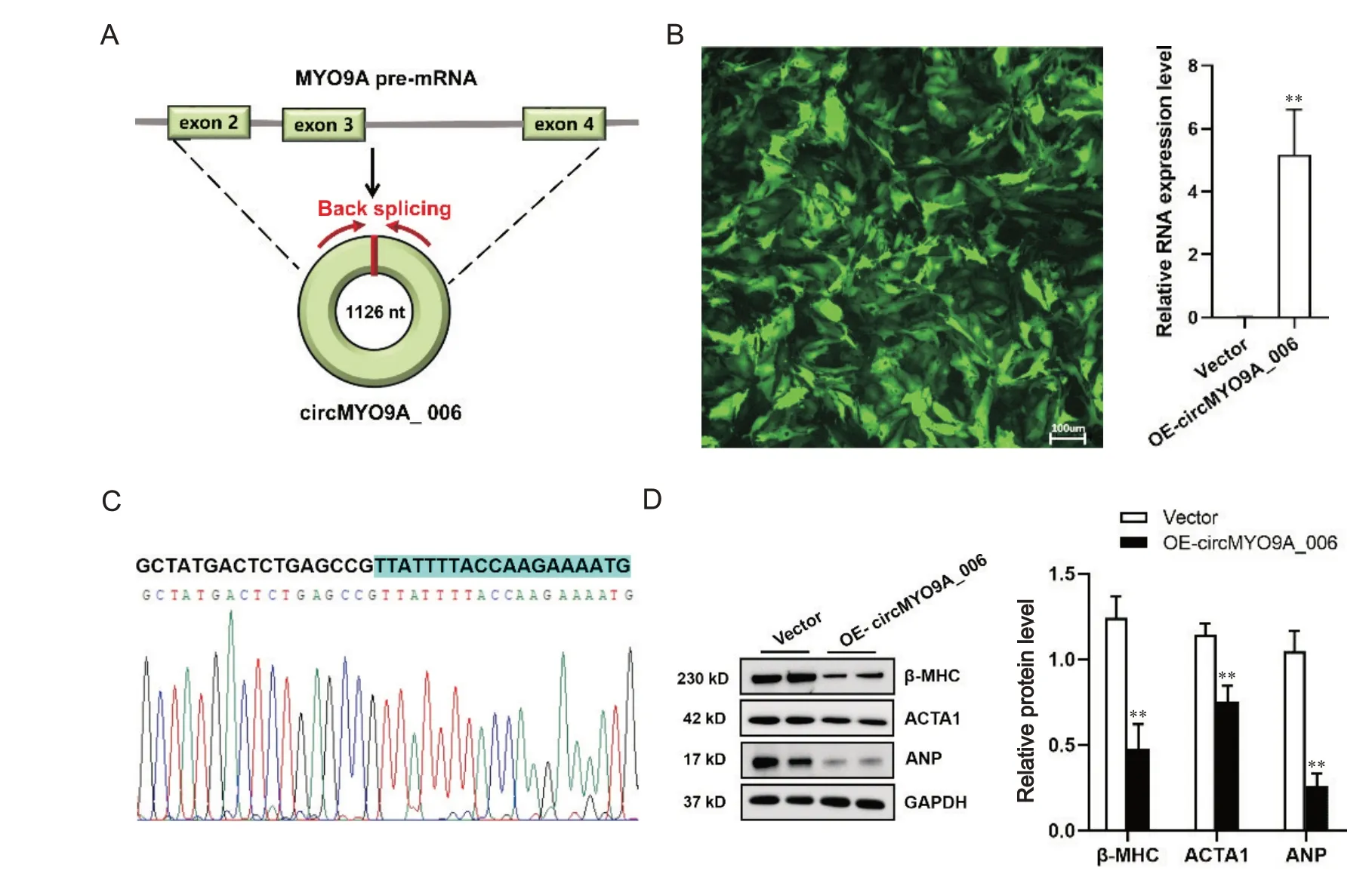
Figure 1. Overexpression of circMYO9A_006 inhibited the expression of hypertrophy-related proteins in neonatal mouse ventricular cardiomyocytes (NMVCs). A: circMYO9A_006 sequence is derived from exon 2 to exon 4 of MYO9A gene; B: the infection efficiency of rAd-circMYO9A_006 was monitored by the co-expressed marker green fluorescent protein, and the expression of circMYO9A_006 was determined by RT-qPCR; C: DNA Sanger sequencing results showed the correct junction sequence of the exogenously overexpressed circMYO9A_006 in NMVCs; D: the protein expression of β-myosin heavy chain(β-MHC), skeletal muscle actin alpha 1(ACTA1) and atrial natriuretic peptide (ANP) was decreased in NMVCs with overexpression of circMYO9A_006. Mean±SD. n=3. **P<0.01 vs vector group.图1 过表达circMYO9A_006抑制NMVCs中肥大相关蛋白表达
2 过表达circMYO9A_006 可抑制NRVCs 的肥大表型
利用腺病毒介导在分离培养的NRVCs 中过表达circMYO9A_006。Western blot 结果显示,与感染对照腺病毒组相比,过表达circMYO9A_006 可有效抑制NRVCs 中肥大相关蛋白β-MHC、ACTA1 和ANP的表达(P<0.01),见图2A。建立去氧肾上腺素(phenylephrine, PE)诱导的NRVCs 肥大模型。鬼笔环肽染色结果显示,过表达circMYO9A_006 可显著缩小PE 诱导的NRVCs 细胞表面积(P<0.05),表明过表达circMYO9A_006 可抑制PE 诱导的NRVCs 肥大表型。
人胚肾293T 细胞株来源于美国细胞培养物收藏中心(ATCC)细胞库。细胞在补充有10%澳洲胎牛血清(fetal bovine serum, FBS)的DMEM/F12 培养液(Gibco)中培养,培养条件为37 ℃、5% CO2的加湿细胞培养箱。
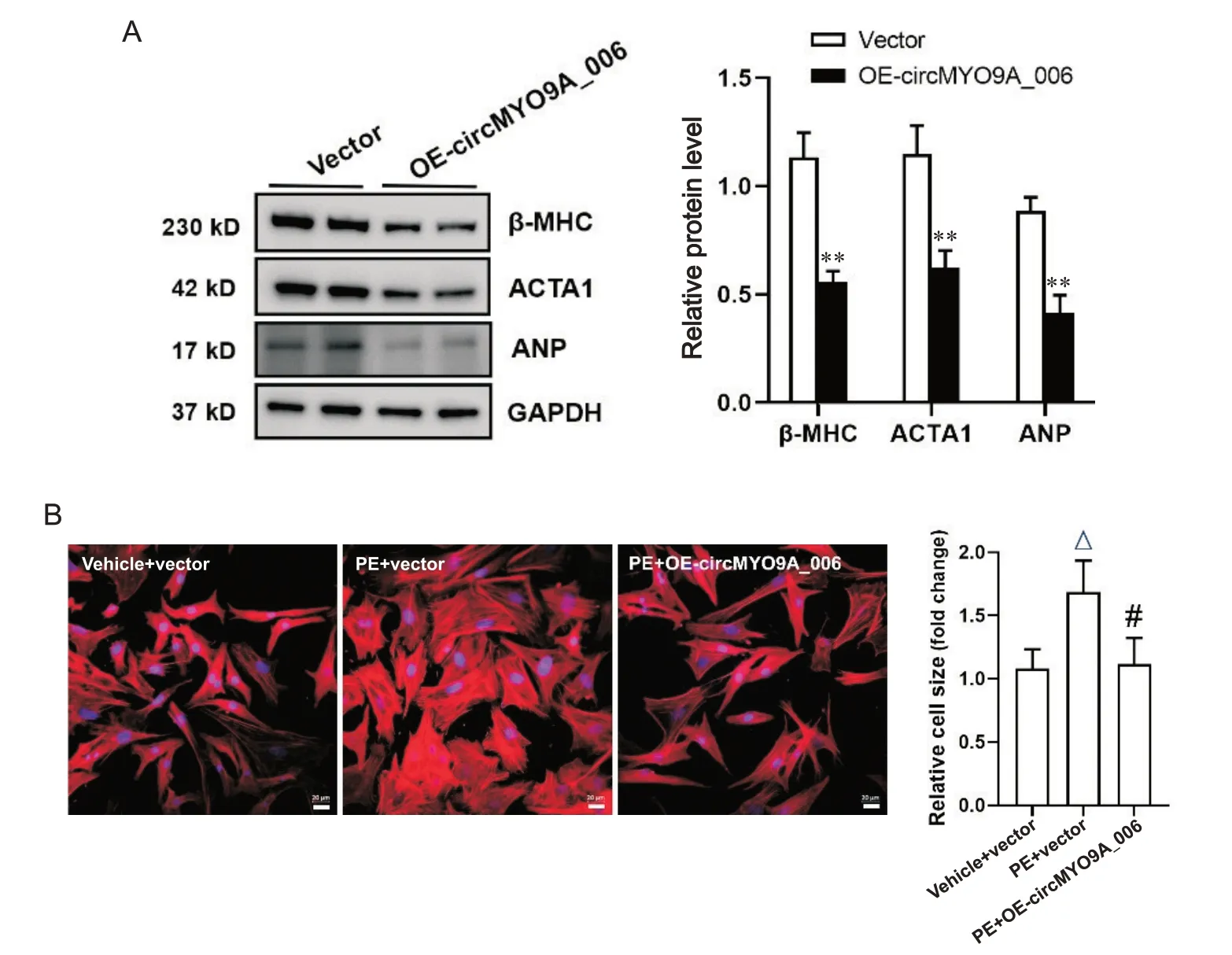
Figure 2. Overexpression of circMYO9A_006 inhibited the expression of hypertrophy-related proteins in neonatal rat ventricular cardiomyocytes (NRVCs). A: the protein expression of β-myosin heavy chain (β-MHC), skeletal muscle actin alpha 1 (ACTA1) and atrial natriuretic peptide (ANP) was suppressed in NRVCs with overexpression of circMYO9A_006; B: the morphological changes of NRVCs observed by Phalloidin-iFluor 647 staining showed that overexpression of circMYO9A_006 reduced phenylephrine (PE)-induced NRVC size (scale bar=20 μm). Mean±SD. n=3. **P<0.01 vs vector group; △P<0.05 vsvehicle+vector group; #P<0.05 vsPE+vector group.图2 过表达circMYO9A_006抑制NRVCs中肥大相关蛋白表达
3 circMYO9A_006 可翻译出含208 个氨基酸的蛋白
通过在线程序circBank (http://www. circbank.cn/searchCirc.html)进行序列分析提示,circMYO9A_006包含2个潜在的IRES和1个可翻译出含208个氨基酸蛋白(命名为MYO9A-208aa)的ORF(图3A、B)。我们将两个潜在IRES 的DNA 序列定向克隆到萤光素酶报告基因载体上,双萤光素酶报告基因实验结果显示,与阳性对照circ_0001742 相似,circMYO9A_006所包含的两个IRES序列具有介导下游ORF翻译的作用(图3C)。Western blot结果显示,在NRVCs中过表达circMYO9A_006后,在28 kD位置有特异表达的蛋白条带(图3D)。通过对成功过表达MYO9A-208aa 的NRVCs 进行蛋白核质分离实验,发现该翻译蛋白主要定位于细胞质中(图3E)。

Figure 3. Identification of circMYO9A_006-translated MYO9A-208aa protein. A: the IRES and ORF sequences in circMYO9A_ 006;B: the predicted amino acid sequences of circMYO9A_006-translated MYO9A-208aa protein; C: detection of the activity of 2 IRES sequences in circMYO9A_006 by dual-luciferase reporter assay (FL: firefly luciferase; RL: Renilla luciferase);D: identification of circMYO9A_006-translated MYO9A-208aa in neonatal rat ventricular cardiomyocytes (NRVCs) by Western blot; E: identification of the intracellular localization of MYO9A-208aa in NRVCs by Western blot. MYO9A-208aa was shown accumulated in the cytoplasm.图3 circMYO9A_006翻译蛋白MYO9A-208aa的鉴定
4 MYO9A-208aa 介导了circMYO9A_006 对心肌细胞肥大的抑制作用
为了进一步探究circMYO9A_006 翻译蛋白的生物学功能,本研究分别构建了circMYO9A_006 的ORF 中ATG 突变(circMYO9A_006-ATG-mut,ATG 变为AG,不能起始翻译)和表达circMYO9A_006-ORFflag 的载体,见图4A。Western blot 结果显示,circ-MYO9A_006-ATG-mut 不能翻译产生MYO9A-208aa,只有circMYO9A_006-flag 和circMYO9A_006-ORFflag 可一致表达MYO9A-208aa 蛋白(图4B)。同时,只有过表达circMYO9A_006-flag 和circMYO9A_006-ORF 可一致地下调NRVCs 中心肌细胞肥大相关蛋白β-MHC、ACTA1 和ANP 的表达(P<0.01),并可逆转PE 诱导的NRVCs 肥大反应(P<0.05),而过表达circMYO9A_006-ATG-mut没有上述作用,见图4C、D。
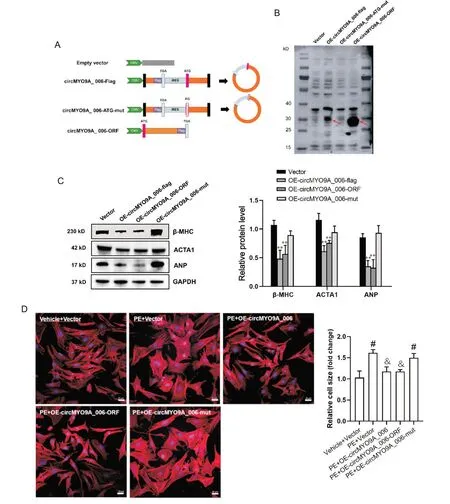
Figure 4. circMYO9A_006 inhibited the hypertrophic phenotype via translating MYO9A-208aa protein. A: sequence characteristics of empty vector, circMYO9A_006-flag, circMYO9A_006-ATG-mut and circMYO9A_006-ORF; B: the expression of MYO9A-208aa-flag protein in neonatal rat ventricular cardiomyocytes(NRVCs) detected by Western blot; C: overexpression of circMYO9A_006-flag and circMYO9A_006-ORF could consistently inhibit β-myosin heavy chain (β-MHC), skeletal muscle actin alpha 1 (ACTA1) and atrial natriuretic peptide (ANP) expression in NRVCs; D: the morphological changes of NRVCs exposed to phenylephrine (PE) treatment were observed by Phalloidin-iFluor 647 staining, and the results showed that overexpression of circMYO9A_006-flag and circMYO9A_006-ORF could markedly reverse PE-induced increase in size of NRVCs (scale bar=20 μm). Mean±SD. n=3. **P<0.01 vs vector group; #P<0.05 vsvehicle+vector group; &P<0.05 vsPE+vector group.图4 circMYO9A_006通过翻译MYO9A-208aa发挥抑制NRVCs肥大表型的作用
讨 论
心肌肥大是一种对左心室壁应力升高的代偿性心肌增厚反应[14-16]。随着长期处于持续的压力刺激,这种病理性心肌肥大会进一步加重演变成不可逆转的心肌损伤[17]。在本研究中,我们证实circMYO9A_006具有抑制NMVCs和NRVCs中肥大相关蛋白表达的作用,以及抑制PE 诱导的NRVCs 肥大的作用,即circMYO9A_006具有抑制心肌细胞肥大的作用。
绝大多数circRNA 是由宿主基因的相关外显子反向剪接环化构成[18-19],虽然circRNA 通常要低于其线性宿主基因mRNA 的表达水平,但共价闭合的环状结构可使其免于核酸内切酶的降解,导致其在体内半衰期通常较长,进而可能发挥更为持久的生物学功能[20]。近年来的研究证实一些circRNA 具有翻译蛋白的能力[21-23]。由于circRNA 缺乏5′端帽子结构,故其翻译主要依赖于潜在ORF 前短元件的m6A修饰或者IRES的驱动来实现[24]。
序列分析提示,circMYO9A_006具有2个潜在的IRES 序列。我们分别构建萤光素酶报告基因载体,并通过双萤光素酶报告基因实验来检测证实2 个IRES 序列均具有介导核糖体与circMYO9A_006 结合,并启动蛋白翻译的能力。蛋白表达结果显示,circMYO9A_006 可翻译出蛋白MYO9A-208aa,且该蛋白主要定位于细胞质中。
为明确MYO9A-208aa 对心肌细胞肥大表型的调节作用,我们分别构建了编码MYO9A-208aa 的ORF 表达载体和circMYO9A_006-ATG-mut 表达载体。功能实验结果显示,circMYO9A_006 和circ-MYO9A_006-ORF 均可表达MYO9A-208aa 蛋白,一致性地抑制心肌细胞肥大相关蛋白β-MHC、ACTA1和ANP 的表达,并可逆转PE 诱导的NRVCs 肥大反应,而circMYO9A_006-ATG-mut 不具有上述作用。上述结果表明翻译蛋白MYO9A-208aa 介导了circ-MYO9A_006对心肌细胞肥大的抑制作用。
借鉴游客感知价值测量相关研究,依据“Means-End”理论,参考我国《中华人民共和国旅游行业规范——旅游景区游客中心设置与服务规范》(LB/T011-2011)对于游客中心相关建设标准、临安大峡谷村游客中心现状,设计游客中心属性层与结果层题项;参考KAHLE[23]的LOV价值观列表,制定使用游客中心可能满足的游客最终目的题项,量表指标来源如表1所示。设计访谈问卷如表2所示。
综上所述,本研究证实了circMYO9A_006 通过其翻译蛋白MYO9A-208aa 发挥抑制心肌细胞肥大的作用。在后续研究中,我们将在整体动物水平来明确circMYO9A_006 抑制心肌肥大的作用,阐明翻译蛋白MYO9A-208aa 介导circMYO9A_006 发挥抑制心肌肥大作用的分子机制。
