三裂棉不育胞质杂交组合F1、F2 产量优势分析
2024-03-04唐会妮张梦王舜戚廷香吴建勇郭立平张学贤陈亮亮王海林乔秀琴梅拥军邢朝柱
唐会妮,张梦,王舜,戚廷香,吴建勇,郭立平,张学贤,陈亮亮,王海林,乔秀琴,梅拥军,邢朝柱
(1.塔里木大学农学院,新疆 阿拉尔 843300;2.中国农业科学院棉花研究所/ 棉花生物育种与综合利用全国重点实验室/ 农业农村部棉花生物学与遗传育种重点实验室,河南 安阳 455000;3.商丘市睢阳区乡村产业发展服务中心,河南商丘 476000)
杂种优势是自然界中普遍存在的一种生物学现象,其利用已被证明可显著提高棉花产量、改善纤维品质以及增强抗病虫性等[1]。 目前,生产上大面积推广种植的棉花杂交种主要是由人工去雄授粉获得的。 随着劳动力成本日益上涨,通过人工去雄实现杂种一代大规模生产难以为继。为解决这一问题,基于“三系”(不育系、保持系和恢复系)的细胞质雄性不育系(cytoplasmic male sterility, CMS)杂交制种技术由于操作简单、节约成本和制种纯度高等优点,已成为棉花杂种优势利用的重要手段[2]。近年来,国内育种家已成功选育出一系列强优势棉花胞质不育“三系”杂交种,如邯杂301[3]、中棉所99[4]和鲁杂2138[5]等。 然而,由于所用的陆地棉胞质和哈克尼西棉胞质不育系均为孢子体不育类型,其杂种F2会出现1/4 的不育株[6],从而导致产量显著降低,因此无法在生产上大面积利用。
三裂棉CMS(CMS-D8)系是新的棉花配子体不育类型,目前也已实现了“三系”配套,其与恢复系杂交产生的杂种F1自交所得F2群体中无任何配子不育或育性特征的分离[6]。 因此,将CMS-D8 系应用于棉花三系杂交种子生产,不仅能够免去人工去雄成本,而且可以大大提高D8 不育胞质杂种二代制种产量[7-8]。 关于CMS-D8 胞质对棉花主要经济性状的影响,目前已有一些研究报道。王江林等[9]研究发现D8 不育胞质对杂交种F1产量性状具有极显著的负效应;对纤维长度和长度整齐度则无明显影响,可使断裂比强度提高1.0~1.1 cN·tex-1,降低马克隆值1.4~1.5, 并增加断裂伸长率0.4 百分点。 Zhang 等[10]认为与D2 不育胞质相比,D8 不育胞质对棉花产量性状尤其是皮棉产量具有更大的负效应。 另有研究表明,一些陆地棉人工去雄杂交F2的产量仍保持了一定的剩余优势[11],说明繁殖简单、 种子成本低廉的F2在当前新疆主产棉区机械化密植条件下具有潜在的应用价值[7,11]。 然而,D8不育胞质杂种二代能否在生产上利用目前还未见相关报道。
本研究结合强优势杂交棉品种选育实践,通过在2 个不同的生态点对7 个棉花CMS-D8 胞质杂交组合F1、F2的产量优势表现进行系统分析, 旨在为生产上利用D8 不育胞质杂种二代提供一定的理论依据。
1 材料与方法
1.1 供试材料
以7 个细胞核背景不同的CMS-D8 不育系(多代回交创制) 为母本、1 个综合性状优良且具有强恢复力的哈克尼西棉雄性不育(CMS-D2)恢复系ZHR142 (含Rf1基因的恢复系可同时恢复棉花CMS-D2 和CMS-D8 的育性[8])为父本,配制了7 个杂交组合,编号为1~7(表1)。
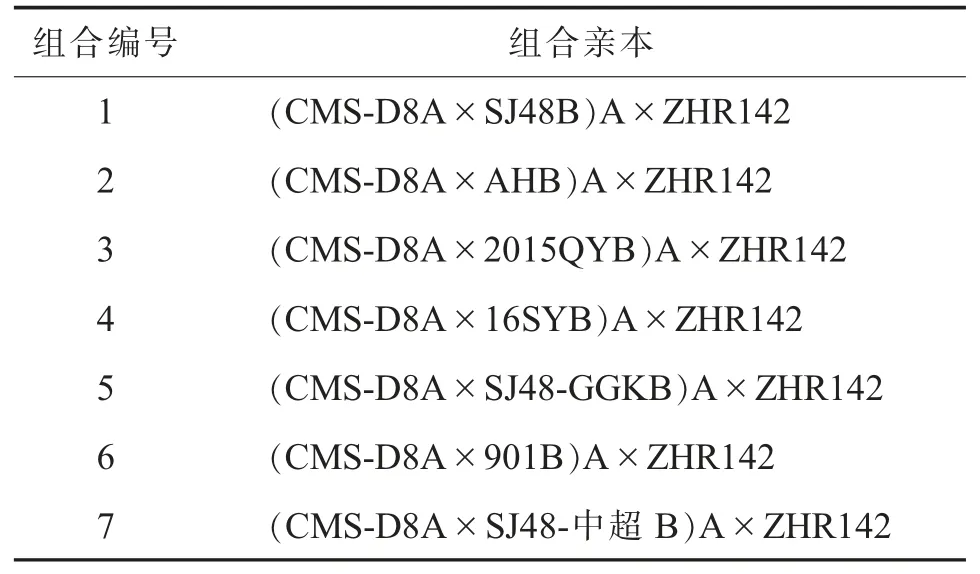
表1 CMS-D8 胞质杂交组合亲本与代号
2020 年4 月下旬, 将7 个CMS-D8 系和恢复系ZHR142 材料种植于河南安阳白璧镇中国农业科学院棉花研究所东场试验基地, 杂交获得F1种子。 同年10 月份将7 个F1材料种植于海南三亚中国农业科学院棉花研究所崖城试验基地,自交获得F2种子。 2021 年4 月下旬,将7 个F1、F2及其亲本(由于不育系无法自交结铃, 在本研究中使用其同核异质保持系SJ48B、AHB、2015QYB、16SYB、SJ48-GGKB、901B 和SJ48- 中超B 的产量性状数据来代替对应的不育系计算中亲和超亲优势)和对照品种分别种植在西北内陆棉区的新疆阿拉尔 (E1)和黄河流域棉区的河南安阳(E2)2 个生态点。 其中,阿拉尔点和安阳点分别选用国家区域试验对照品种中棉所49(CK1)和瑞杂816(CK2)。
1.2 试验设计
采用随机区组设计,3 次生物学重复。 其中,安阳点采用裸地直播,每个材料3 行区,行长9 m,行距0.8 m;阿拉尔点采用覆膜直播,每个材料1 膜3行,每个小区面积约为9.6 m2,均设置保护行。 种植密度按照不同生态区设置,其中安阳生态点为每666.7 m23 000 株,阿拉尔生态点为每666.7 m210 000 株。试验材料管理措施与当地普通大田一致。
1.3 性状调查
于吐絮前(9 月下旬),分别在2 个生态点进行单株结铃数调查,具体计算方法参照前人的研究[12]。于吐絮期(10 月上旬),在安阳点从每个小区随机收取棉株中部正常吐絮的30 个铃; 在新疆阿拉尔点从每个小区收取棉株中部完全吐絮的50 个棉铃,进行室内考种,计算铃重、衣分等产量性状。 之后,计算每个小区的籽棉产量,并按小区面积换算成每666.7 m2籽棉产量,根据衣分计算每666.7 m2皮棉产量。
1.4 数据处理
本试验调查数据用Microsoft Excel 2016 进行整理与分析。 按下列公式计算中亲优势(mid-parent heterosis,MPH)、超亲优势(better-parent heterosis,BPH)、竞争优势(competitive heterosis,CH)和优势衰退率(heterosis decline rate,HDR):MPHi(%)=(X1,i或X2,i-MPi)/MPi×100;BPHi(%)=(X1,i或X2,i-BPi)/BPi×100;CHi(%)=(X1,i或X2,i-CKi)/CKi×100;HDRi(%)=(X1,i-X2,i)/X1,i×100。其中,X1,i是杂 种 一代产量 性 状i的 平 均 值,X2,i是 杂 种二代产量性状i的平均值,MPi是双亲产量性状i的平均值,BPi为双亲中表现较好亲本的产量性状i的平均值,CKi为生态点相应对照品种产量性状i的平均值。
2 结果与分析
2.1 中亲和超亲优势分析
对阿拉尔点7 个CMS-D8 胞质杂交组合不同世代F1、F2的产量及其构成因素中亲及超亲优势进行分析,结果(表2)显示:F1铃重、籽棉产量和皮棉产量的平均中亲和超亲优势均为正值;F2铃重的平均中亲和超亲优势分别为11.21%和9.62%, 籽棉产量表现出0.37%的平均中亲优势,其他产量性状的平均中亲和超亲优势均为负值。在籽棉产量和皮棉产量方面,除组合2 外,其他组合F1均具有正向中亲和超亲优势; 组合3、6 和7 的F2中亲和超亲优势仍为正值(表2)。 上述结果说明这3 个组合的杂种二代在阿拉尔点仍具有一定的产量剩余优势。
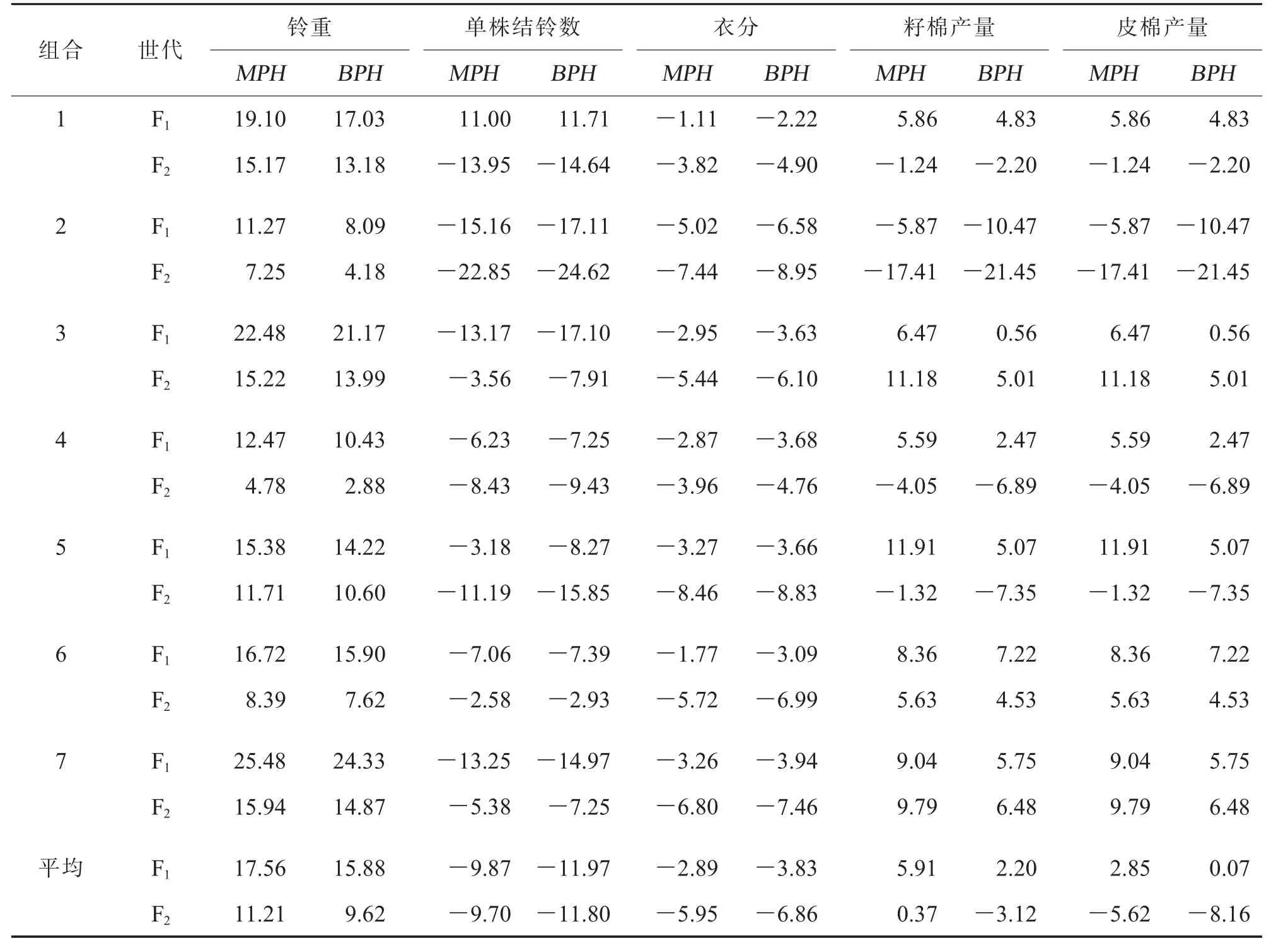
表2 阿拉尔点7 个组合F1 和F2 产量性状中亲和超亲优势表现 %
在安阳点,F1铃重、 籽棉产量和皮棉产量的平均中亲优势为正值; 除铃重外,F1其他产量性状的平均超亲优势均为负值(表3)。 F2单株结铃数的平均中亲优势为0.60%,其他产量性状平均中亲优势和超亲优势均为负值。 就籽棉产量和皮棉产量而言,组合1、6 和7 的F1和F2中亲优势均为正值,但仅组合6 的F1和F2籽棉产量具有正向超亲优势,分别为4.01%和3.25%(表3)。 综合来看,组合6 和7 的F1、F2在2 个生态点较亲本仍具有一定产量优势。
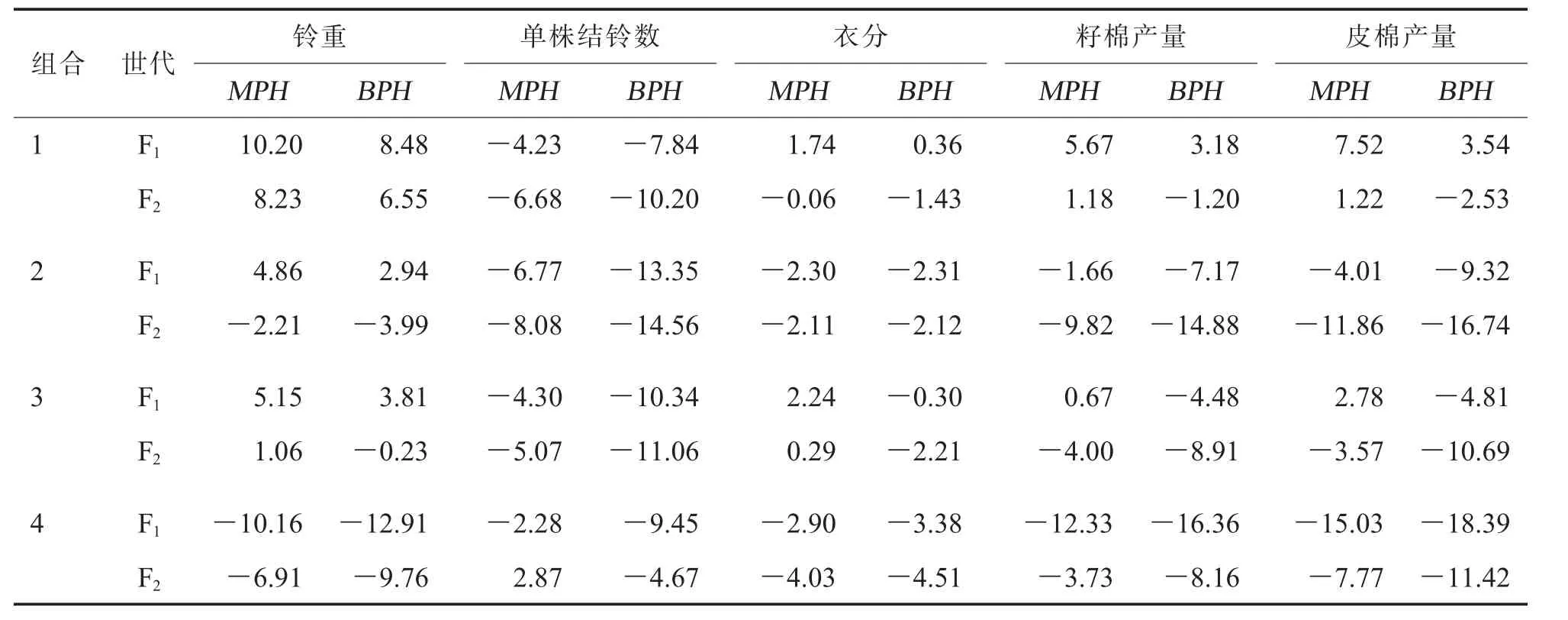
表3 安阳点7 个组合F1 和F2 产量性状中亲和超亲优势表现 %
2.2 竞争优势分析
对阿拉尔点7 个CMS-D8 胞质杂交组合F1、F2和对照品种CK1(中棉所49)的产量及其构成因素进行差异比较分析,结果见表4。 在铃重上,所有7个组合的F1均表现为正向竞争优势, 其中组合3、4、6 和7 的铃重较对照差异达显著或极显著水平;各组合F2铃重均较F1有所降低, 但除组合2 和4外,其他组合仍具有正向竞争优势,尤其是组合3和7 的铃重显著或极显著高于对照。就单株结铃数而言,7 个杂交组合的F1、F2均表现为负向竞争优势,其中组合2 的F1、F2以及组合3 和7 的F1单株结铃数显著少于对照。 所有组合的F1衣分均具有正向竞争优势, 其中组合1 极显著高于对照;F2除组合5 的衣分显著低于对照外,其余组合均表现为正向竞争优势。 对于籽棉产量和皮棉产量来说,除组合2 的F2显著低于对照外, 其余组合在不同世代与对照无显著性差异, 组合6 和7 的F1、F2均表现为正向竞争优势(表4)。
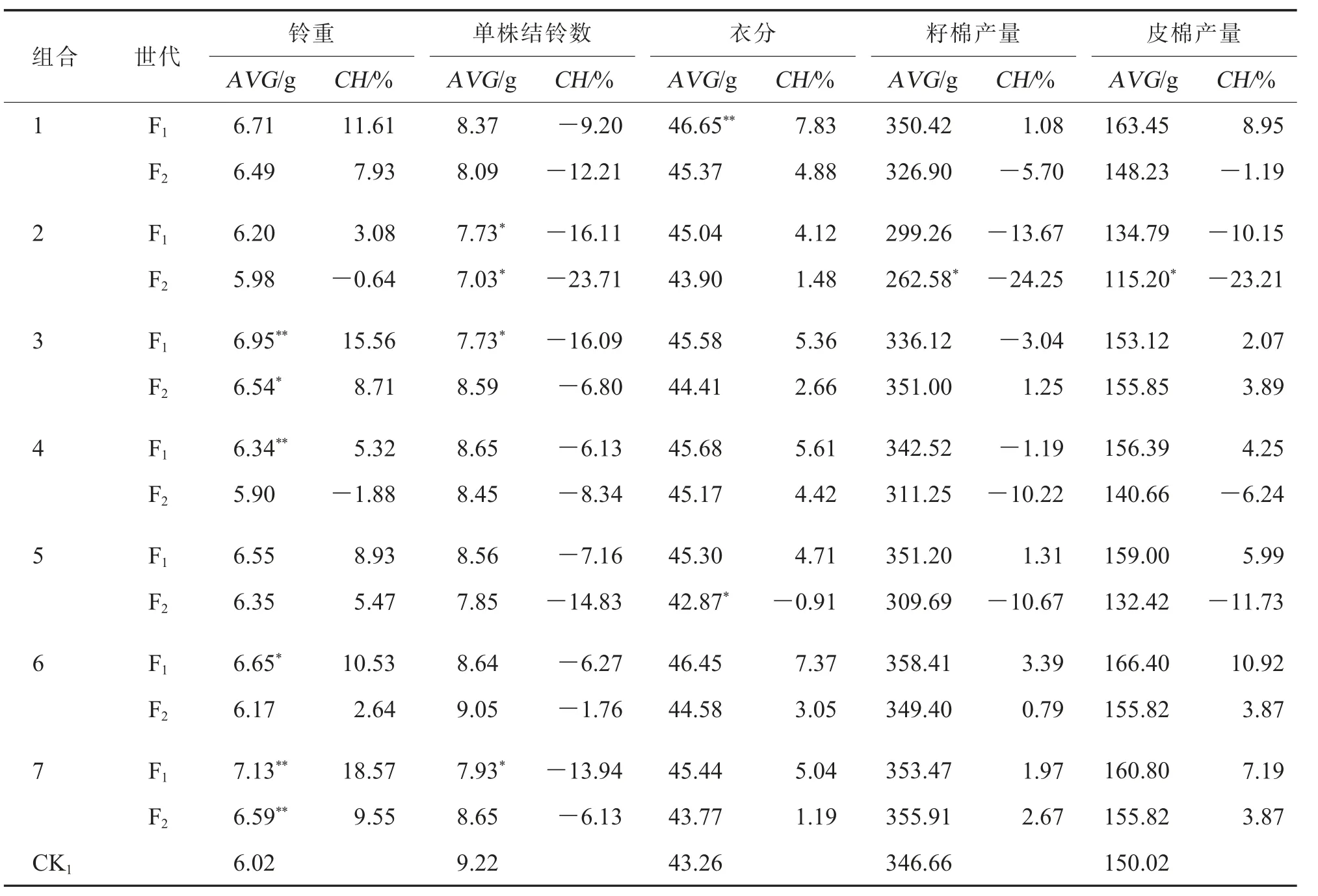
表4 阿拉尔点7 个组合F1 和F2 产量性状竞争优势分析
在安阳点,就铃重来看,各杂交组合不同世代F1、F2均表现出不同程度的负向竞争优势, 其中组合4 和5 的F1以及组合2、3、4、5、7 的F2与对照瑞杂816 差异达显著水平(表5)。 除组合2 和3 外,其他组合F1的单株结铃数均多于对照,但仅组合6达到差异显著水平; 组合4、5、6 和7 的F2单株结铃数表现为正向竞争优势, 同时多于对应的F1,其中组合6 的F2单株结铃数显著多于对照。 对于衣分性状,除组合1 的F1呈显著的正向竞争优势外,其余组合不同世代与对照相比无显著差异。在籽棉产量和皮棉产量方面, 只有组合1、6 和7 的F1表现为正向竞争优势,但差异均不显著;除组合6 的F2籽棉产量高于对照外, 其余组合均低于对照,但仅组合2 的籽棉产量和皮棉产量较对照差异达显著或极显著水平(表5)。 由此可见,在安阳点,组合1、6 和7 的F1产量具有一定的竞争优势,F2与对照相比差别不大; 大部分F2产量竞争优势减退主要是由于铃重的降低。
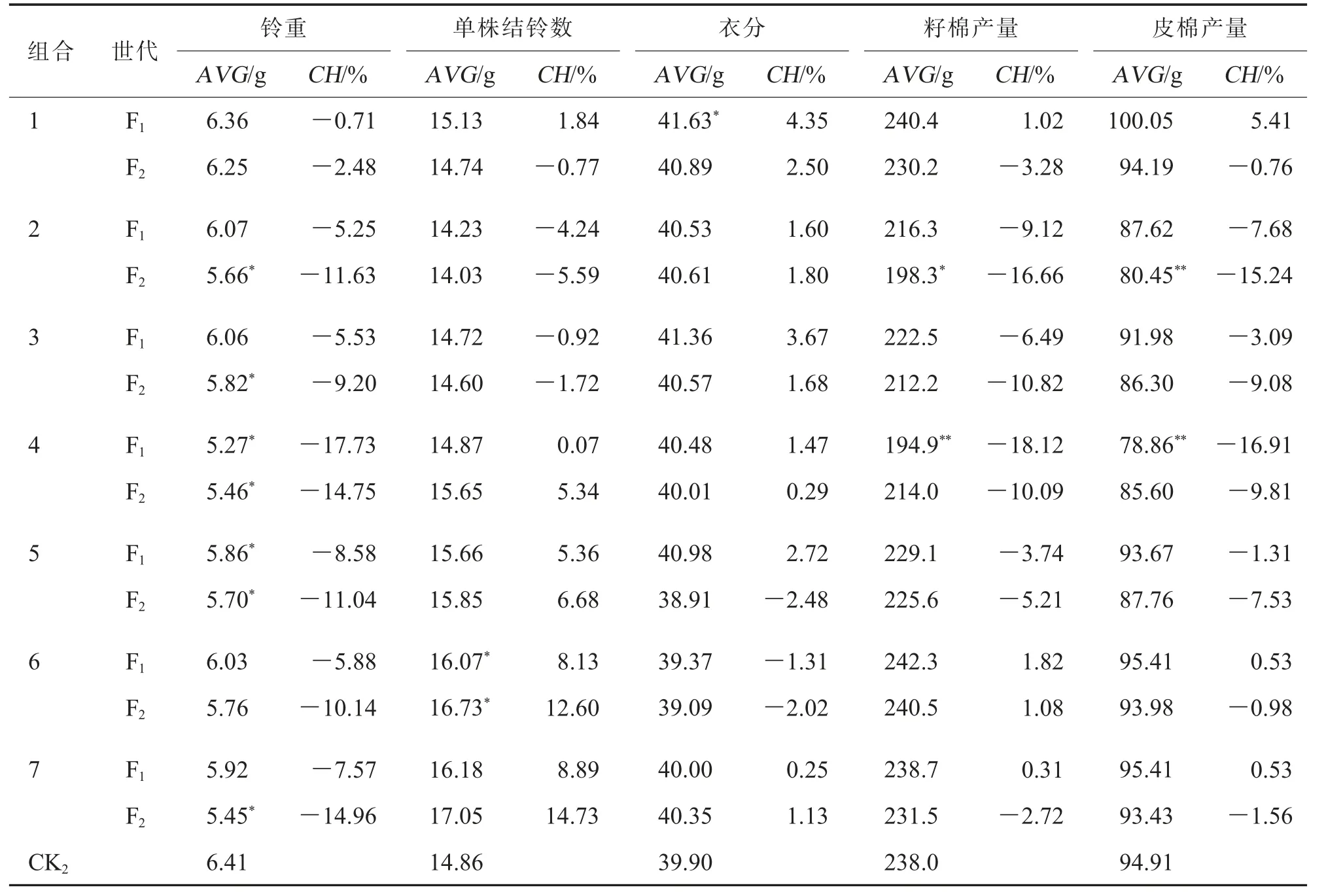
表5 安阳点7 个组合F1 和F2 产量性状竞争优势分析
2.3 优势衰退分析
对7 个CMS-D8 胞质杂交组合F2的5 个产量性状在2 个生态点进行优势衰退分析,结果(表6)表明:F2除单株结铃数外其他产量性状在2 个生态点均表现出一定程度的优势衰退,整体上阿拉尔点优势衰退率高于安阳点。在阿拉尔点,铃重、籽棉产量和皮棉产量的平均优势衰退率分别为5.37%、5.33%和8.33%;在安阳点,铃重和皮棉产量的优势衰退表现较为明显,平均优势衰退率分别为3.43%和3.08%。
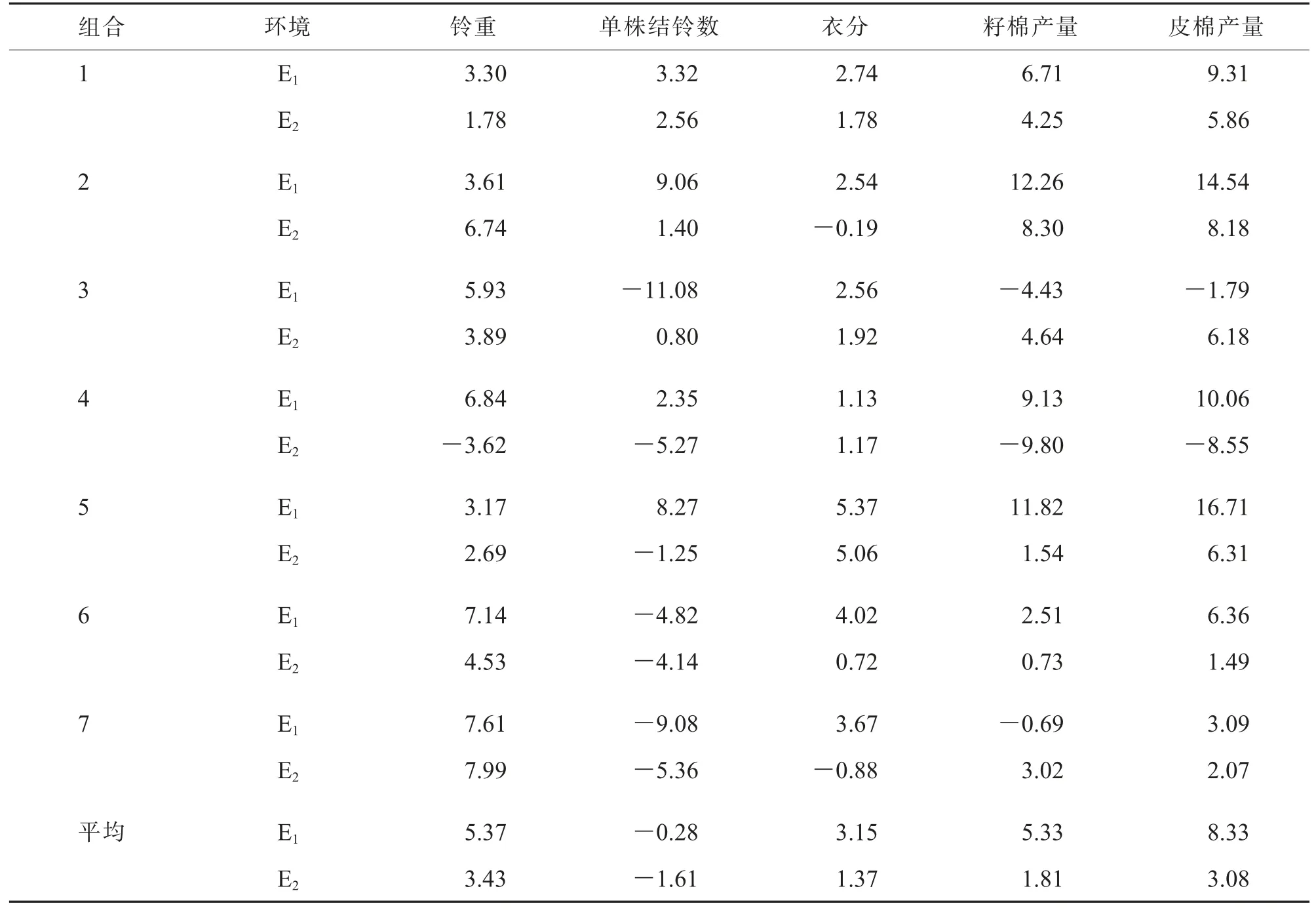
表6 在2 个生态点F2 产量性状优势衰退表现%
具体来看,各杂交组合大部分产量性状表现在F2中有所衰退, 且不同性状的优势衰退程度不同,其中籽棉产量和皮棉产量优势衰退率相对较大,分别为-9.80%~12.26%和-8.55%~16.71%。 组合3、6 和7 的F2在2 个试点的籽棉产量和皮棉产量优势衰退不明显,衰退率相对较小(表6)。 说明这些D8 不育胞质F2在产量方面可能仍具有一定的剩余优势。
3 讨论与结论
3.1 棉花CMS-D8 胞质杂交组合不同世代产量优势表现
棉花具有明显的杂种优势,一般来说杂交棉相比常规棉的产量更高、品质更优[1]。虽然已有学者研究发现CMS-D8 胞质对棉花主要产量性状具有一定的负效应[9-10],但目前关于D8 不育胞质F1、F2的产量优势表现仍缺乏系统研究,其杂种二代的潜在育种价值也未受到足够重视。 本研究发现,组合1、3、6 和7 的不同世代F1、F2在2 个生态点相比亲本来说具有一定产量优势(表2、表3),尤其组合6和7 的F1、F2产量在阿拉尔点均表现为正向竞争优势(表4),说明这2 个组合的F2在产量方面具有一定的剩余优势,可以利用其F2的杂种优势。 大部分D8 不育胞质杂交F2产量竞争优势减退主要是由于铃重的降低(表4、表5),这与裴小雨等[13]以及Chen 等[11]关于人工去雄授粉杂交F2产量竞争优势的研究结果基本一致。 此外,F2单株结铃数在2 个生态点平均表现为无优势衰退(表6),这可能是由于D8 不育胞质F2群体中完全可育株的花粉育性好于部分可育的F1,最终导致成铃数表现出一定程度的增加[6]。
3.2 D8 不育胞质杂种二代育种应用前景初步探讨
陆地棉属于异源四倍体, 其F2分离现象不如二倍体作物水稻、玉米那么严重,而且棉花的收获期比较长,F2株型及熟性分离对产量和纤维品质不构成直接影响,为棉花杂种二代利用提供了可能[7]。与常规棉品种相比, 优势表现显著的杂种F1单产能提高10%~30%,而一些F2产量仍可提高5%~10%[11,14-15]。目前,新疆已成为我国棉花主产区,国家统计局数据显示2022 年新疆棉花产量为539.1 万t,占全国棉花总产量的90.2%。 然而,由于西北内陆棉区主要采用机械直播方式,用种量大,大面积推广利用人工去雄获得的杂交种,成本高昂,难以实现[8]。 利用CMS-D8 配子体不育系选育强优势“三系”杂交棉品种应用于生产可以很好地解决这一问题,不仅能省去人工去雄成本、提高杂交F1纯度,而且能显著提高D8 不育胞质F2制种产量。在本研究中,尽管大部分D8 不育胞质杂交组合F2产量优势较F1有所衰退, 但仍有部分组合如3、6 和7 的F2籽棉产量和皮棉产量优势衰退不明显,衰退率较小(表6),且组合6 和7 的F2产量在阿拉尔点具有一定的正向竞争优势(表4),说明部分D8 不育胞质F2仍具有一定的产量剩余优势, 理论上可利用其F2的杂种优势。 有研究报道,D8 不育胞质对纤维品质性状无明显不利影响,在一定程度上还可以明显改善棉花品种的纤维细度[9]。 因此,通过对棉花CMS-D8“三系”亲本进行回交改良和合理选配,并结合多年、多生态点交叉选择,选育出一些产量性状衰退较小或无衰退的D8 不育胞质杂种二代在我国主产棉区推广应用是切实可行的[7]。
