花生籽仁蔗糖含量遗传模型分析
2024-02-14郅晨阳薛晓梦李雄才晏立英陈玉宁康彦平王志慧淮东欣洪彦彬姜慧芳廖伯寿
郅晨阳 薛晓梦 吴 洁 李雄才 王 瑾 晏立英 王 欣 陈玉宁 康彦平 王志慧 淮东欣,* 洪彦彬 姜慧芳 雷 永,* 廖伯寿
1中国农业科学院油料作物研究所 / 农业农村部遗传育种重点实验室, 湖北武汉 430062; 2 襄阳市农业技术推广中心, 湖北襄阳 441021; 3 河北省农林科学院粮油作物研究所, 河北石家庄 050035; 4 广东省农业科学院作物研究所, 广东广州 510640
花生是世界范围内重要的油料作物和经济作物之一, 我国是世界花生生产和消费大国, 其产量和消费量持续增长[1]。近年来, 随着人们消费需求的改变, 高品质食用花生在消费市场中崭露头角[2-3], 对其风味品质也提出了更高的要求。研究表明, 甜味与花生口味品质呈显著正相关, 相关系数高达0.88[4], 甜味较高时, 花生口感较好。可溶性糖是花生甜味的主要来源, 其中蔗糖所占比例最高, 约占可溶性糖的90%, 与总糖含量极显著正相关[5-6],当蔗糖含量高于5%, 花生籽仁可以品尝到明显的甜味[7], 表明蔗糖含量是影响花生甜味的关键因素,因此籽仁蔗糖含量是评价花生风味的关键指标。
研究发现, 花生种质资源中蔗糖含量变异丰富,是可遗传的数量性状[7-9], 改良潜力大。已有研究表明, 花生蔗糖含量可能受两对主基因+多基因调控,主要受母体效应控制[10-11]。通过定位研究, 分别在A06、A08和B06染色体上鉴定到调控花生蔗糖含量的主效QTL[12-14]。通过转录组-代谢组联合分析,推测花生籽仁蔗糖含量可能来源于光合产物的非完全转化[15]。虽然花生籽仁蔗糖含量研究已取得初步进展, 但其调控机制仍不明确, 因此, 开展花生籽仁蔗糖含量的遗传研究分析, 探索其规律, 对指导花生蔗糖含量定位研究及花生品质育种具有重要意义。
本研究利用高糖品种(系) SYT5-1 (6.41%)和冀花甜1号(JHT1, 6.34%)分别构建了2个F2:3群体:NYBP×SYT5-1、19-1934×冀花甜1号, 分别获得了467份和149份F2:3家系, 通过测定2个群体籽仁蔗糖含量, 分析蔗糖含量与含油量、蛋白质含量以及籽仁相关性状之间的相关性, 利用SEA 2.0软件对蔗糖含量进行了遗传模型分析, 推测其遗传规律,为进一步探索蔗糖含量调控机制、指导食用型花生品种培育奠定基础。
1 材料与方法
1.1 群体材料
本研究以4个蔗糖含量存在极显著差异的品种(系): SYT5-1 (6.41%)、南阳白皮(NYBP, 2.53%)、19-1934 (2.61%)和冀花甜1号(JHT1, 6.34%)为亲本,配置2个杂交组合: NYBP×SYT5-1、19-1934×冀花甜1号, 种植于中国农业科学院油料作物研究所阳逻基地, 按组合收获F1、F2代种子, 分别获得467份和149份F2:3家系的花生籽仁, 以其为试验材料开展研究。其中, SYT5-1和19-1934分别为中国农业科学院油料作物研究所选育的高糖品系和高油品系,南阳白皮为本单位保存的种质资源, 冀花甜1号由河北省农林科学院育成并提供。
1.2 表型分析
1.2.1 花生籽仁蔗糖含量及蛋白质含量测定 利用近红外模型测定花生籽仁蔗糖含量和蛋白质含量。每份材料选取15~20粒饱满无病变的花生种子,将其放置于小样品杯中, 利用Sperctra Star XL近红外光谱仪(美国Unity科技公司)采集光谱并预测蔗糖含量和蛋白质含量, 每份材料重复3次, 分别计算蔗糖含量和蛋白含量的平均值。
1.2.2 花生籽仁含油量测定 利用脉冲核磁共振仪分析法, 使用核磁共振仪(PQ001)对花生籽仁含油量进行测定。每份材料选用10粒饱满无病变的花生籽仁称取其质量, 利用NMR (Nuclear Magnetic Resonance)技术得到的信号量与含油量的线性方程,计算样品含油量, 每份材料重复3次, 以平均值作为该材料的含油量。
1.2.3 花生籽仁长、籽仁宽及百仁重测定 花生籽仁产量性状统计: 选取15粒均匀饱满的花生籽仁,利用万深SC-G型自动考种仪及千粒重仪, 分别获得籽仁长、籽仁宽和百仁重, 每份材料重复3次, 取平均值。
1.2.4 数据分析 利用SPSS 19统计F2:3群体花生蔗糖含量、含油量、蛋白质含量和籽仁性状数据,并计算各性状间的相关系数。
应用SEA 2.0软件程序[16]对花生蔗糖含量进行主基因+多基因遗传模型分析。依据AIC准则和极大似然值筛选AIC值最小及极大似然值较大的遗传模型作为备选模型, 并对获得的所有模型进行适合性检验, 包括: 均匀性检验(U12、U22、U32)、Smirnov检验(nW2)、Kolmogorov检验(DN), 筛选适合性检验参数均未达到显著或极显著水平、具有其统计学意义的模型进行遗传参数分析。使用最小二乘法对蔗糖含量遗传模型进行遗传参数估算确定最佳遗传模型。
2 结果与分析
2.1 2个群体F2蔗糖含量频数分布特征
对两群体花生籽仁蔗糖含量频数分布进行统计(图1)发现, 在NYBP×SYT5-1中, 蔗糖含量的变异范围为0.19%~8.20%, 呈现连续分布的特点, 且群体蔗糖含量偏度与峰度绝对值均小于1 (附表1), 总体呈现近似正态分布, 表明蔗糖含量可能受多基因调控; 19-1934×JHT1群体中, 蔗糖含量变异范围更大, 为0.09%~9.51% (附表1), 同样呈现连续分布,但是表现明显的双峰分布趋势, 表明蔗糖含量可能由少量主基因调控。在2个群体中都观察到明显的超亲分离现象, 其中在NYBP×SYT5-1中蔗糖含量高于亲本SYT5-1的家系共27个, 在19-1934×JHT1群体中蔗糖含量高于亲本JHT1的家系共9个, 这些优异家系为花生高糖育种提供了材料基础。
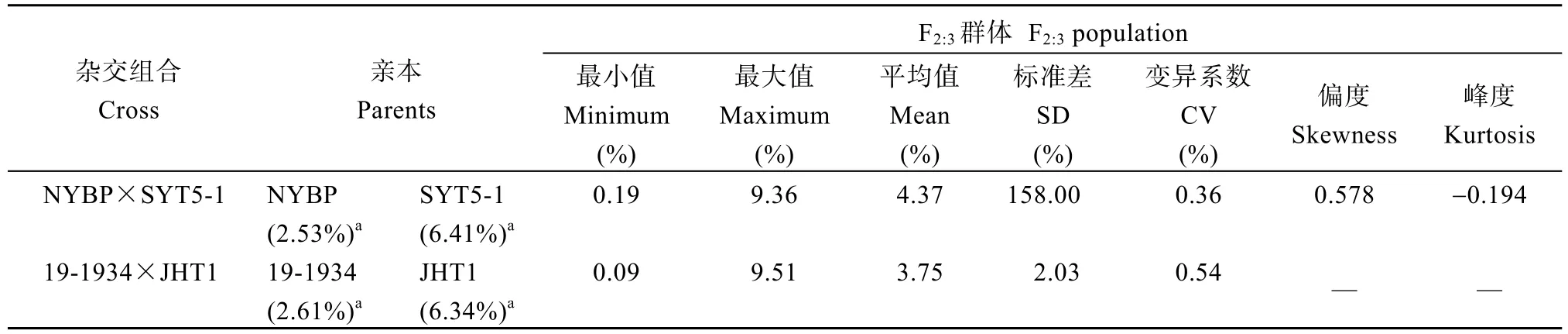
附表1 F2:3群体蔗糖含量表型统计Table S1 Phenotypic statistics of sucrose content in F2:3 population
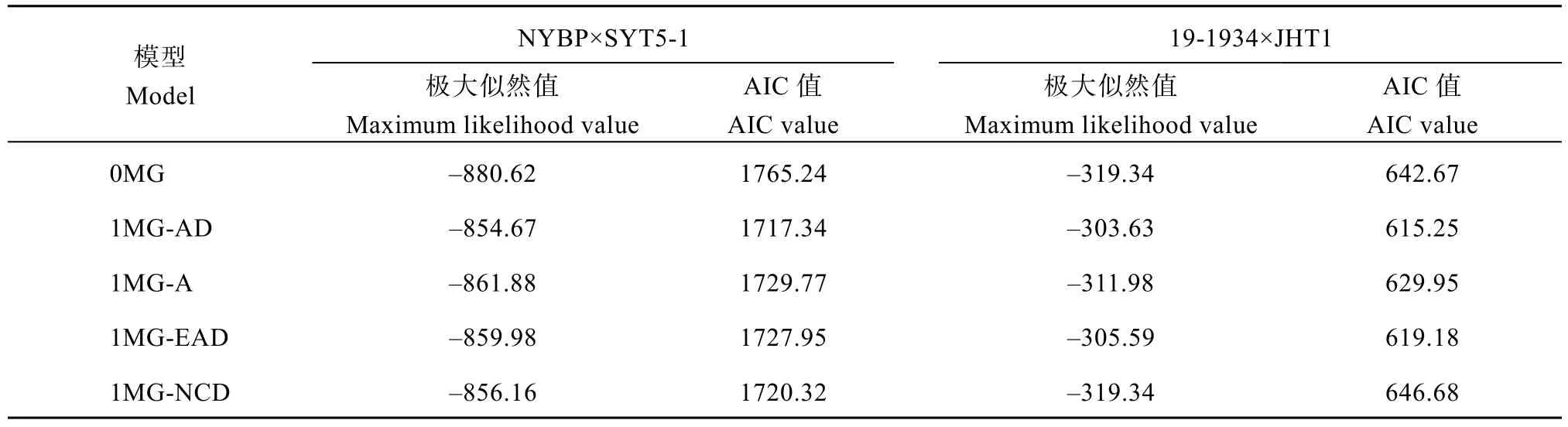
表1 花生籽仁蔗糖含量遗传模型分析Table 1 Genetic model analysis of sucrose content in peanut seeds

图1 花生籽仁蔗糖含量频数分布图Fig. 1 Frequency distribution of sucrose content in peanut kernel
2.2 花生籽仁蔗糖含量与含油量、蛋白质含量及产量相关性状之间的相关性分析
由图2可知, 在NYBP×SYT5-1群体中, 花生籽仁蔗糖含量与含油量呈极显著负相关(r= -0.57**),与蛋白质含量显著正相关(r= 0.52**), 但与籽仁长、籽仁宽和百粒重间无明显相关关系。在19-1934 ×JHT1中, 籽仁蔗糖含量与含油量同样显著负相关(r= -0.80**), 与蛋白质含量也显著正相关(r=0.75**),而与籽仁长、籽仁宽和百仁重间都表现为显著负相关, 相关系数分别为-0.41**、-0.39**、-0.42**。同时发现, 在2个群体中, 籽仁含油量与蛋白质含量显著负相关, 相关系数在2个群体间分别为-0.47**、-0.62**。

图2 花生籽仁蔗糖含量与其他性状之间相关性Fig. 2 Correlations between sucrose content and other traits in peanut kernel
综上, 在两群体间, 籽仁蔗糖含量与含油量均呈显著负相关, 与蛋白质含量均显著正相关, 同时发现, 籽仁含油量与蛋白质含量间也存在极显著的负相关。但在2个群体中, 蔗糖含量与籽仁长、籽仁宽和百仁重3个性状间的相关性不一致, 在19-1934 × JHT1中蔗糖含量与三者显著负相关, 但在NYBP×SYT5-1群体中不存在相关关系。
2.3 花生籽仁蔗糖含量的遗传模型分析
使用SEA 2.0软件中SEA-F23程序对花生籽仁蔗糖含量进行主基因+多基因模型分析发现, NYBP×SYT5-1与19-1934×JHT1群体均获得11种遗传模型(表1), 模型适合性检验结果显示, 所有模型均为达到显著或极显著水平, 具有其统计学意义(表2)。
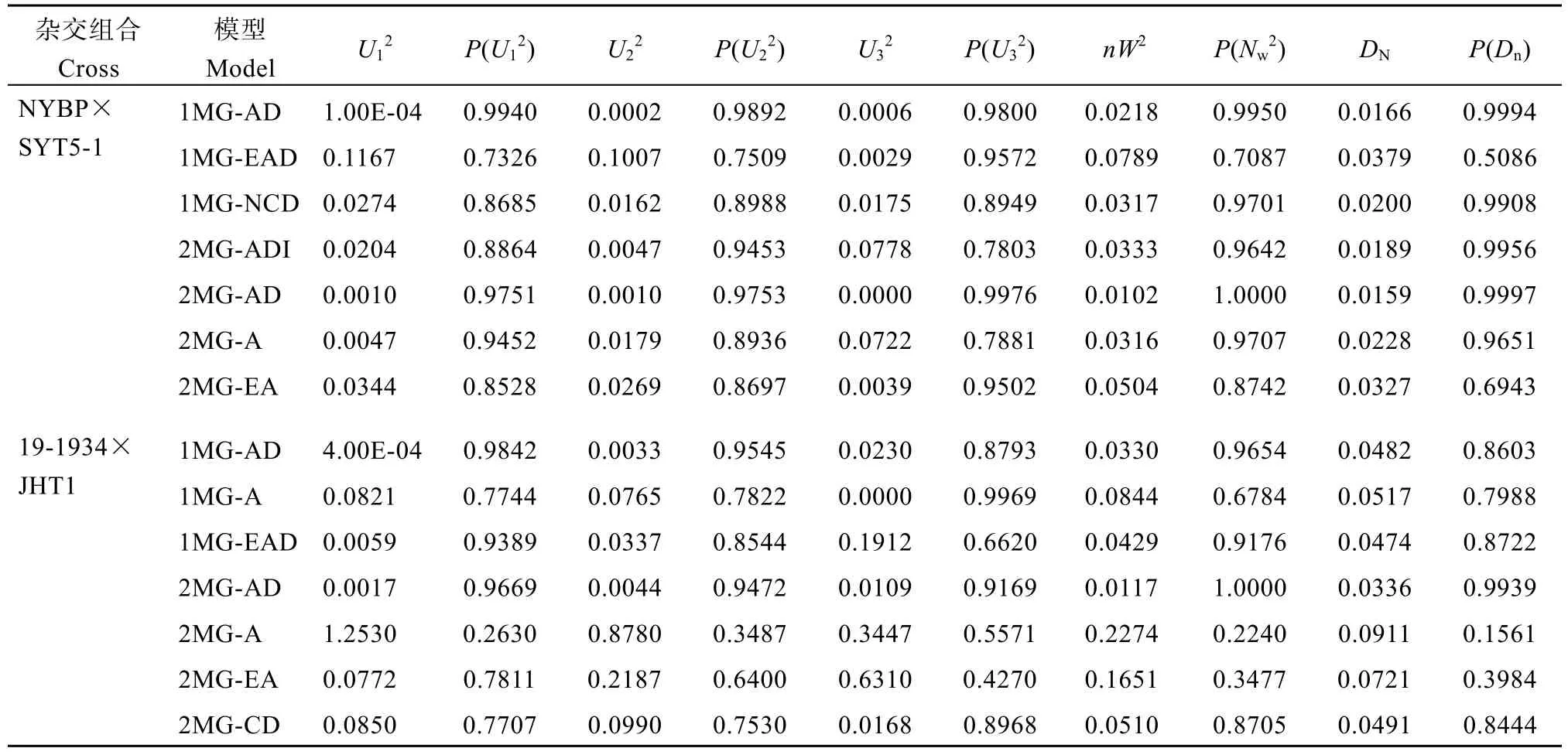
表2 花生籽仁蔗糖含量候选模型的适合性检验Table 2 Suitability test of candidate models for sucrose content in peanut seeds
依据极大似然值最大及AIC值最小原则对2个组合间所有遗传模型进行筛选(表1)发现, 在NYBP×SYT5-1群体中, 2MG-AD极大似然值为–847.85,AIC值为1707.69, 是最优候选模型; 在19-1934×JHT1中, 2MG-AD同样具有最大的极大似然值及最小AIC值, 分别为-298.02与608.03, 是最优候选模型。综上, 两群体最优候选模型均为2MG-AD, 即两对主基因控制加性显性效应是最优遗传模型。
一阶参数结果表明(表3), 在NYBP×SYT5-1群体中, 第1对主基因与第2对主基因加性效应分别为da=1.6676、db=1.2171, 显性效应分别为ha=-0.6996、hb= -0.3973; 19-1934×JHT1群体中, 两对主基因的加性效应分别为da=1.7948、db=0.7131, 显性效应分别为ha=1.9106、hb= -0.1276。

表3 花生籽仁蔗糖含量一阶参数估计值Table 3 Estimates of first order genetic parameters for sugar content in peanut kernels
两群体间, 两对主基因的加性效应均表现为|da|>|db|, 即表明两对主基因调控蔗糖含量表型间,以其中一对主基因的加性效应为主; 显性效应|ha|>|hb|, 表明组合中控制籽仁蔗糖含量的其中一对主基因显性效应大于另一对主基因, 显性效应更加明显。因此, 调控花生籽仁蔗糖含量的主基因间存在明显的互作, 且其中一对主基因的加性效应和显性效应大于另一对主基因, 对控制花生籽仁蔗糖含量的效应更加明显。
二阶遗传参数结果显示(表4), NYBP×SYT5-1中, 最优模型2MG-AD的主基因方差与主基因遗传率最高, 分别为2.2810和85.9102; 19-1934×JHT1组合中, 主基因方差与遗传率为4.0724和97.7852。在两群体间蔗糖含量的主基因遗传率很高, 表明蔗糖含量主要受遗传效应控制, 受环境影响较小。
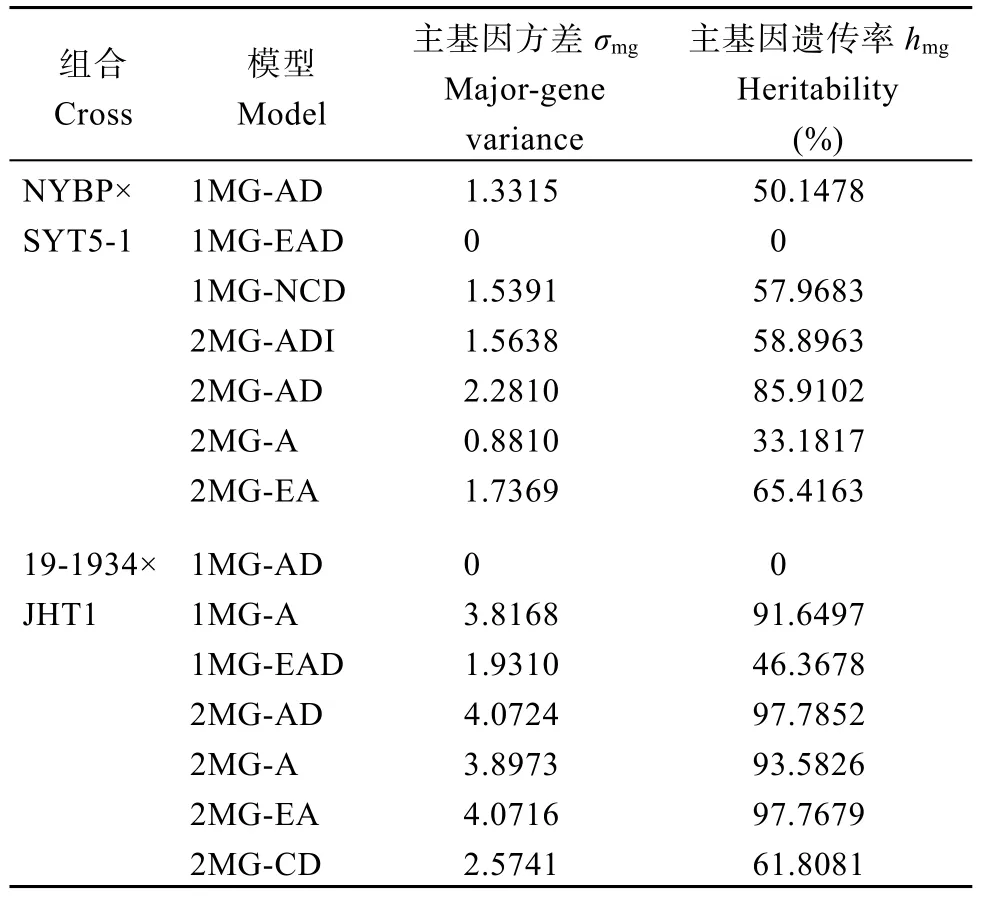
表4 花生籽仁蔗糖含量二阶遗传参数估计值Table 4 Estimated values of genetic parameters of sucrose content in peanut kernels
3 讨论
在我国, 花生是主要的油料和经济作物之一,我国花生总产的约40%用于食用加工, 食用花生的消费量也在逐年增长[17]。蔗糖含量是影响花生及其加工食品风味的重要因素, 已成为食用花生品质遗传改良的重要方向, 研究蔗糖含量的遗传规律将为开展高糖花生育种奠定重要的理论基础。本研究利用2个高糖材料SYT5-1和JHT1, 分别构建了F2:3群体, 杂交后代中蔗糖含量呈连续分布, 变异范围分别为0.19%~8.20%和0.09%~9.06% (图1), 与Bishi等在60份珍珠豆型资源材料及41份印度主栽培品种中检测的蔗糖含量(2.44%~7.61%和2.61%~6.50%)相比[7-8], 杂交群体中蔗糖变异范围更大, 创造了更多的变异。同时发现, 本研究2个群体间蔗糖含量表现出显著不同的连续分布特征, NYBP×SYT5-1群体蔗糖含量呈正态分布(图1-A), 但19-1934×JHT1群体的双峰分布趋势明显(图1-B)。对比前人研究结果,同样发现不同材料构建的群体间, 蔗糖含量的分布特征存在差异, NYBP×SYT5-1群体与张晓军等[12]构建的分离群体中, 蔗糖含量均呈近似正态分布, 表明花生蔗糖含量在这些群体中可能受多基因调控[18];而本研究19-1934×JHT1群体与秦利等[11]、Guo等[14]的试验结果均表现出明显的双峰分布, 可能受少量主基因调控。造成这种差异的原因可能与选用的杂交亲本携带不同的蔗糖含量主效位点相关, 加上不同组合群体间亲本材料遗传背景存在差异, 导致不同群体间蔗糖含量分布特征存在差异。
本研究的2个杂交分离群体中, 花生蔗糖含量与含油量均呈极显著负相关(r= -0.57**,r= –0.80**)(图2)。在大豆、油菜等油料作物中, 蔗糖含量与含油量也表现出类似的负相关性[19-21], 这种负相关性可能与油料作物籽仁中的油脂来源于蔗糖相关, 多项研究结果表明, 油料作物的光合产物以蔗糖形式运输到籽仁, 并被转化分解, 为脂肪酸和三酰甘油的合成积累提供最主要碳源[22-24]。在油菜种子胚中,随着胚发育, 蔗糖含量逐步降低, 而油脂含量不断积累增加[24]。同时, 蔗糖还作为一种重要的信号物质调控糖酵解途径, 影响该途径中多种基因的表达,对下游油脂合成途径产生影响[25]。蓖麻种子中糖代谢相关基因表达量下调引起油脂积累相关基因表达量的明显提高[26]。在以淀粉作为主要储藏物质的玉米、水稻等作物相关研究中, 与糖代谢相关的基因的突变造成糖分积累并引起淀粉含量的下降或淀粉性质的变化[27-29],OsSWEET11突变体中糖-淀粉代谢途径明显变化, 对水稻颖果发育产生了重要影响[30]。因此推测, 花生籽仁中蔗糖含量的积累可能是油脂合成受阻引起。
在本研究两群体中, 花生籽仁蔗糖与蛋白质含量均呈显著正相关(r= 0.52**,r= 0.75**), 含油量与蛋白质含量显著负相关(r= –0.47**,r= –0.62**, 图2),大豆相关研究存在相似结论[31]。籽仁中油脂和蛋白质的合成主要以蔗糖的分解产物丙酮酸为共同底物[32], 二者存在底物竞争关系, 油脂含量与蛋白质含量通常表现为显著负相关[33-35]。因此推测, 花生籽仁内蔗糖含量与蛋白质含量显著正相关可能与油脂合成受阻、碳源流向蛋白质合成有关。
在19-1934×JHT1群体中, 蔗糖含量与籽仁长、籽仁宽、百仁重显著负相关(图2-B), 这与油菜、大豆中的研究结果相似[36-38], 籽仁中蔗糖含量变化可以引起每角果粒数(荚粒数)和粒重等相关性状的变化。蔗糖含量高低总体体现了植物体内碳源供应水平, 除了为种子油脂积累提供所需的碳源, 蔗糖也为胚组织细胞的发育和分裂提供养分[39-40], 进而影响了籽仁发育。在玉米、水稻、大豆等研究中表明,蔗糖转运蛋白SWEET相关基因在作物驯化过程中受到选择, 对产量和品质都具有重要影响, SWEET基因的突变造成蔗糖转运受阻, 引起胚乳发育不良,籽粒大小及粒重均显著降低[27,41]。膜转运蛋白MFS家族也可通过蛋白互作方式影响水稻光合产物的分布平衡, 调控灌浆、籽粒大小及粒数等重要性状[42]。在本研究2个群体中, 蔗糖含量与籽仁大小相关性状的相关性不一致, 也可能与亲本材料具有不同的遗传背景相关。
本研究遗传分析结果表明, 蔗糖含量具有很高的遗传率, 分别为85.91%与97.79% (表4), 与秦利等[11]、胡美玲等[10]研究结果一致。花生籽仁蔗糖含量遗传率高, 表明其主要受基因型调控, 环境因素对其影响较小, 因此在育种群体低世代即可进行蔗糖含量的选择, 以加快育种进程, 提高育种效率。遗传模型分析表明, 花生籽仁蔗糖含量主要受两对主基因的加性显性效应调控(表3), 与秦利等[11]研究结果一致。两对主基因的加性效应|da|>|db|、显性效应|ha|>|hb|, 表明主基因间存在明显互作, 且以其中一对主基因的加性显性效应为主。同时, 两群体中加性效应da>db>0, 两对主基因的加性效应均为正效应,即基因间的加性效应可以增强, 具有累加效应。花生籽仁蔗糖含量遗传率高且受两对主基因控制, 表明蔗糖含量可以通过开发与蔗糖含量相关的分子标记进行快速选择育种, 利用主基因的累加效应开展超亲育种, 进行种质创新和高蔗糖品种育种具有充分的可行性, 为培育不同蔗糖含量的食用花生品种提供了丰富的亲本材料和选择空间, 对花生籽仁蔗糖含量的改良具有重要指导意义。
4 结论
本研究利用高糖材料SYT5-1和JHT1分别构建了2个F2:3群体, 花生籽仁蔗糖含量在两群体中都呈现连续分布特点, 超亲分离现象明显。在两群体中, 花生籽仁的蔗糖含量均与含油量显著负相关,与蛋白含量显著正相关; 但与籽仁长、籽仁宽和百仁重的相关性不一致。花生蔗糖含量主要受两对主基因控制, 两对主基因间存在明显互作, 累加效应明显, 且其中一对主基因的加性效应和显性效应大于另一对主基因, 对花生蔗糖含量的调控效果更加显著。
