保活运输应激对水产动物肌肉品质影响的研究进展
2024-01-26张亮子黄泽南马华威
张亮子,黄泽南,尤 娟,3,刘 茹,3,尹 涛,3, ,马华威
(1.武汉商学院,湖北 武汉 430056;2.华中农业大学食品科学技术学院,湖北 武汉 430070;3.国家大宗淡水鱼加工技术研发分中心,湖北 武汉 430070;4.广西壮族自治区水产科学研究院,广西 南宁 530021)
《中国居民膳食营养指南》指出水产动物是人类蛋白、脂类、B 族维生素和矿物质的良好来源,是平衡膳食的重要组成部分[1]。中国是一个有14亿多人口的大国,要解决好吃饭问题,要树立“大食物观”,既向陆地要食物,又要向江河湖海要食物,这更凸显水产动物的重要性。物流运输是水产品全产业链的关键环节之一,可实现不同地域资源配置,满足消费和加工需求。水产动物的活运形式包含高密度充氧运输(鱼、虾等)、低密度保温运输(虾、蟹、贝等)、单体充氧运输(鱼)、无水运输(鱼、虾、蟹、贝)等[2]。暂养[3]、生态休眠[4]等技术也逐渐在水产动物运输中应用,对减缓应激和提升存活率起到重要作用。近几年,运输应激对肌肉品质的影响成为研究的热点[5-7]。然而,目前已有的关于水产动物运输方面的综述主要是关于应激、存活率、生理特性和调控措施等[8,9],而关于运输应激对肌肉品质影响的系统性综述尚未见报道。因此,本研究系统论述运输中主要应激源和应激减缓措施对水产动物肌肉品质的影响,并介绍组学技术在运输应激对水产动物肌肉品质影响机制研究中的应用动态,以期为提升水产动物品质和开展相关研究提供参考。
1 保活运输中应激对水产动物肌肉品质的影响
水产动物应激反应是指受到外界刺激(应激源)后所产生的异常跳跃、呼吸速率加快,心跳加快,游动速度加快等生理反应[10]。运输过程中温度、低氧、挤压、振动、氨氮和空气暴露等是主要应激源,它们刺激感受器,使脑部产生激素和电信号来激活神经-内分泌反应系统(例如:HSC 轴即下丘脑-交感-嗜铬细胞轴和HPI轴即下丘脑-垂体-肾间组织轴)[11]。神经-内分泌反应系统产生儿茶酚胺类、皮质类固醇激素等化合物,调节能量的消耗和分布,以抵抗或适应应激。正常的鱼体生理处于平衡或者稳态,能量和应激可以看成稳态的两级,当能量作为货币消耗后能够克服应激,重建稳态,那么水产动物能够恢复到正常状态;反之,水产动物则趋近于疾病或者死亡。水产动物在应激反应过程中,生命活动如基因表达、代谢和免疫等都会受到影响,使储能化合物、代谢物质和细胞形态等发生变化,最终可能导致营养组成、质构、色度、蒸煮损失、滴落损失、风味等肌肉品质的变化(表1)。
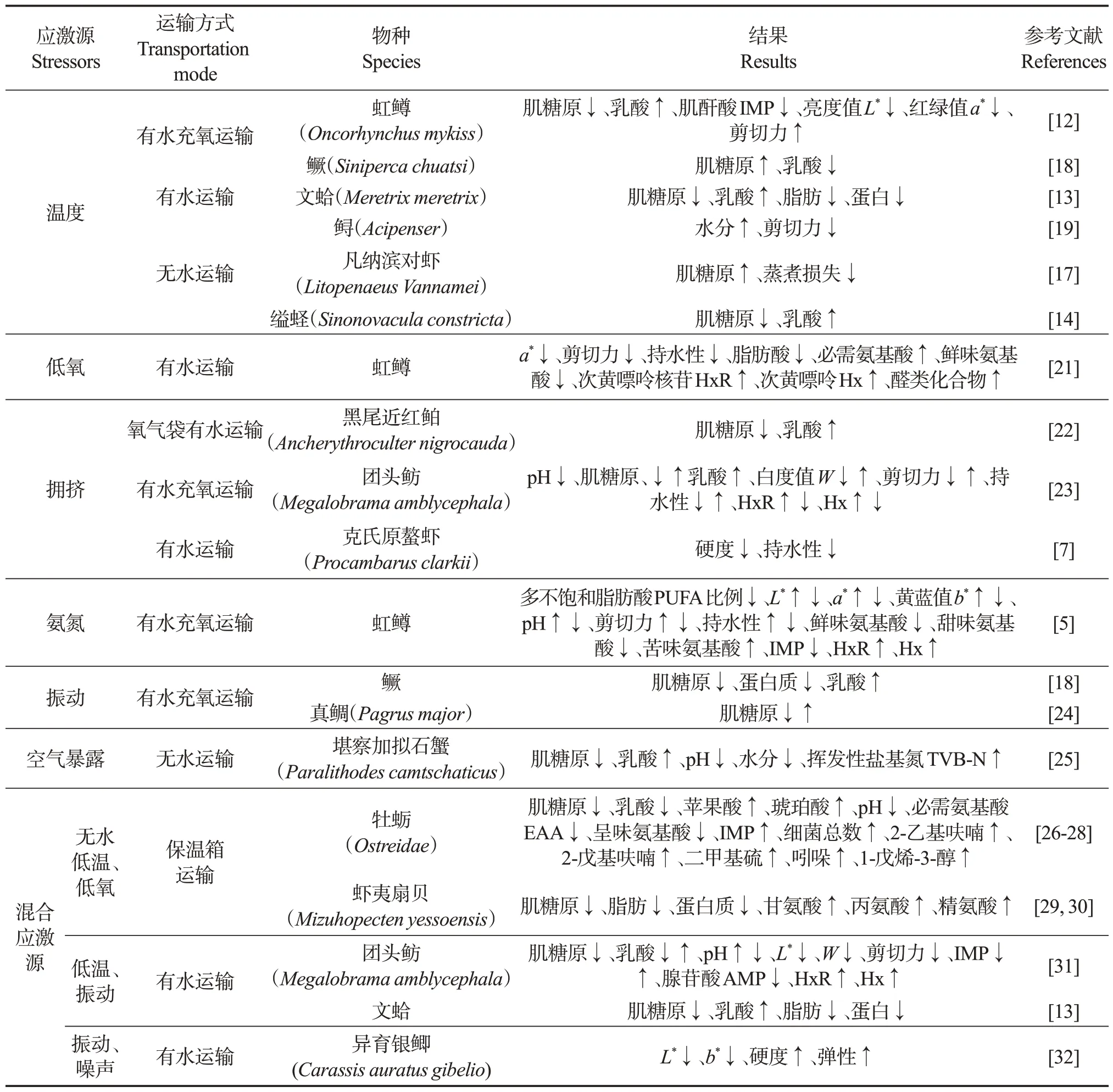
表1 运输中应激对水产动物肌肉品质的影响Table 1 Effects of stress during live transportation on muscle quality of aquatic animals
1.1 温度应激对水产动物肌肉品质的影响
当水产动物从养殖环境转入运输环境,由于密度增加、振动等因素,环境温度会增加,水产动物产生热应激,引起其代谢速率增加,导致在运输中消耗更多的能量。因此在更高的温度下,虹鳟(Oncorhynchus mykiss)[12]、文蛤(Meretrix meretrix)[13]、缢 蛏(Sinonovacula constricta)[14]、鲈(Lateolabrax maculatus)[15]、珍珠龙胆石斑鱼(Epinephelus fuscoguttatus♀×Epinephelus lanceolatus♂)[16]等水产动物在运输过程中肌糖原的消耗增加,并导致乳酸水平上升。除糖原外,脂肪也是水产动物重要的储能物质。随着运输温度的增加,文蛤[13]和鲈[15]的肌肉脂肪含量显著降低。在长时间和极端条件下,水产动物还会降解蛋白质以补充能量的不足。秦小明等[13]等发现在5~20 ℃范围内,当时间超过2 d,高温保活运输文蛤的蛋白质含量明显低于低温下的。
三磷酸腺苷(ATP)是直接供能物质,可降解为具有鲜味的肌苷5ʹ-单磷酸(IMP)。Wu等[12]等发现,在高温运输条件下虹鳟肌肉中IMP 含量较低。相反,在运输过程中加冰、通氧等操作会迅速降低环境温度,水产动物会产生冷应激。因此凡纳滨对虾(Litopenaeus vannamei)[17]和鳜(Siniperca chuatsi)[18]在较低温条件下运输后肌肉中糖原含量显著低于高温下的,而乳酸含量显著较高(P<0.05)。这归因于随着运输时间的延长,一方面鱼和虾逐渐适应了低温环境,另一方面在较低温条件下的代谢速率较低,导致水产动物肌糖原和乳酸含量在不同温度下的差异降低,甚至反转。
除营养品质外,运输温度还影响色度、剪切力和蒸煮损失(表1)。Wu 等[12]等发现,虹鳟在21.61 ℃下运输后肌肉的亮度(L*)、红度值(a*)显著低于在16.54 ℃下;肌红蛋白的氧化和还原形式之间的平衡决定鱼肉的颜色,运输过程中在应激作用下不同状态的肌红蛋白可能是L*和a*值发生变化的原因;另外,他们还发现在高温下运输的虹鳟肌肉的剪切力高于低温下的。然而,Wang等[19]发现高温下鲟(Acipenser schrenckii)肌肉的剪切力低于低温下的。鱼肉剪切力的变化受肌细胞结构、胶原蛋白含量、水分含量、脂肪含量和pH等因素的共同影响。在应激作用下水产动物肌肉细胞结构在凋亡等作用下破裂,会导致剪切力下降[6]。然而,脂肪含量在水产动物应激后被消耗,会增加剪切力[19]。运输过程中,糖原被分解为葡萄糖,葡萄糖可继续被分解为水和二氧化碳,导致肌肉中水分含量增加,从而降低鱼肉剪切力。鲟在高温条件下运输,水分含量显著增加,因此其剪切力下降[19]。蒸煮损失值越高表示营养物质流失越严重。应激改变水产动物的肌肉细胞结构和蛋白质热变性降解程度,可能影响蒸煮损失。在4 ℃运输条件下,凡纳滨对虾受到强烈的冷应激,其蒸煮损失明显高于10 ℃下的[17]。
另外,保活运输温度还可能影响水产动物宰杀后肌肉在贮藏过程中的品质变化。Zhao 等[20]比较16.5 和21.6 ℃运输的虹鳟鱼片在冷藏过程中品质的变化,发现较高的运输温度增加了鱼片的游离水、总挥发性盐基氮(TVB-N)、总活菌数(TVC)和K值,表明运输过程中的热应激加剧虹鳟鱼片在贮藏过程中的质量恶化。
1.2 低氧应激对水产动物肌肉品质的影响
运输过程中由于水产动物密度增加、供氧措施低效和环境中氧气持续消耗等原因,水产动物会出现低氧应激。Wu 等[21]在氧气质量浓度分别为7.5(正常浓度)、5.5(轻度缺氧)和3.3 mg/L(重度缺氧)条件下运输虹鳟,发现低氧应激导致鱼片a*、剪切力、持水性、脂肪酸含量、鲜味氨基酸含量下降,而次黄嘌呤核苷(HxR)含量、次黄嘌呤(Hx)含量、必需氨基酸占总氨基酸比例、醛类化合物含量增加。在低氧应激下,氧合肌红蛋白含量下降,因此a*降低;剪切力的下降主要与低氧应激直接影响胶原的合成及肌节的完整性有关,而持水性降低与肌纤维细胞结构的破坏有关;在对抗低氧应激过程中,能量物质(包含脂肪酸)被大量消耗,必需氨基酸占总氨基酸比例增加是由于组氨酸的含量显著增加导致的,鲜味氨基酸中谷氨酸和甘氨酸含量降低,而ATP 代谢产物中具有苦味物质的HxR、Hx 增加,导致滋味品质下降;虹鳟富含不饱和脂肪酸,在低氧应激后,醛类化合物含量增加导致产生不愉悦的气味,这可能是与n-3 和n-6 等多不饱和脂肪酸的氧化有关。可见,运输过程中的低氧应激对虹鳟鱼片的肌肉品质有负面影响[21]。
1.3 拥挤应激对水产动物肌肉品质的影响
在运输过程中为提升经济效益,水产动物的密度会明显高于养殖过程中的,造成拥挤应激,从而影响肌肉品质(表1)。当运输密度从20 g/L 逐渐升高到40、80 g/L 后,黑尾近红鲌肌糖原含量逐渐下降,而乳酸含量逐渐增加,说明拥挤导致的应激反应逐渐增强[22]。彭玲等[23]也发现,随着运输过程中团头鲂(Megalobrama amblycephala)与水质量比从1∶2.5增加到1∶1.5,肌糖原含量逐渐下降,乳酸含量逐渐增加;当密度进一步增加,肌糖原含量显著增加;另外,在1∶1.5 比例下,鱼肉W、剪切力和持水能力最低,苦味物质的含量(HxR、Hx)最高,意味着在该密度下的应激反应最强烈;当比例从1∶2.5 增加到1∶1.5 时,鱼体间、鱼与水箱壁面间的挤压频率也增加,导致应激反应增强;随着密度的进一步增加,尽管碰撞频率增加,但碰撞距离缩短,碰撞动量反而降低,这可能会减轻应激反应。Shi 等[7]发现,增强运输中拥挤应激反应会降低克氏原螯虾(Procambarus clarkii)虾肉的剪切力和持水性,这主要与蛋白质的降解和肌纤维细胞结构的破坏有关。
1.4 氨氮应激对水产动物肌肉品质的影响
在活鱼运输过程中,氨氮的产生主要是由于鱼类自身的氨转移和脱氨作用,以及厌氧细菌氨化动物粪便残渣、饵残渣等有机物。NH3具有良好的脂溶性,不带电荷,使其更容易与细胞膜上的磷脂双层融合,从而通过细胞膜扩散到血淋巴,增加鱼体内氨氮的浓度,引起应激反应[8,24]。Wu等[5]比较运输过程中不同质量浓度(0.5、10、20、30、45、60 mg/L)氨氮暴露对虹鳟肌肉品质的影响,发现多不饱和脂肪酸(PUFA)占总脂肪酸的比例随着暴露浓度的增加而逐渐下降,其原因是虹鳟血氨积累导致过量活性氧(ROS)的产生,从而诱导氧化应激和脂质氧化;尽管虹鳟在低氨氮浓度暴露(低于30 mg/L)后L*、a*、b*、剪切力、持水性略微增加,但这些肌肉品质参数在高浓度下显著降低,这归因于在强氨氮应激的作用下肌原纤维吸水能力下降,其与胶原和肌原纤维基质的相互作用减弱,水从肌原纤维挤出到位于肌纤维和细胞膜之间的通道,导致肌肉品质劣变。
除上述肌肉品质参数外,虹鳟肌肉的鲜味氨基酸、甜味氨基酸、IMP 含量随着氨氮浓度增加而降低,而苦味氨基酸、HxR 和Hx 含量逐渐增加,表明滋味品质变差。氨暴露通过影响氨基酸代谢中关键酶的活性,导致虹鳟体内游离氨基酸的组成和浓度发生变化。IMP 降低,Hx、HxR 含量增加,这符合ATP 的降解规律,其可能是由于高氨氮应激加速ATP降解所致[5]。
1.5 振动应激对水产动物肌肉品质的影响
与养殖、暂养等环节相比,运输过程的应激源存在不确定性。尤其在活运过程中,公路路况、车辆启停、车辆抖动、风浪引起汽车和船振动也是影响鱼应激的环境因子之一,会引发鱼体产生机械损伤。运输过程中水产动物将储能物质转化为葡萄糖,再通过消耗葡萄糖提供能量来抵抗外界振动应激的影响。因此,在运输过程中鳜(Siniperca chuatsi)的肌糖原和蛋白质含量随着振动频率的增加而降低,乳酸含量增加[18]。真鲷(Pagrus major)肌肉中肌糖原含量也随着振动频率的增加而下降[24]。强烈振动会导致鱼身体无法及时清除自由基,加剧氧化应激反应,破坏抗氧化系统的平衡,从而导致肌肉品质下降,甚至死亡[8]。因此,根据不同公路等级或者洋流情况,规划好运输路径,可在保证品质的同时有效降低运输成本。然而目前关于振动对运输中水产动物肌肉品质影响的研究报道较少。
1.6 空气暴露应激对水产动物肌肉品质的影响
蟹类在无水条件下进行活运,其间会受到空气暴露应激。Lorenzo等[25]研究运输中空气暴露应激(8 ℃、24 h)对堪察加拟石蟹(Paralithodes camtschaticus)肌肉品质的影响,发现肌糖原含量降低,而乳酸含量增加,这与其它水产动物应激后变化一致[12,14]。空气暴露应激后肌肉TVB-N 含量增加20%,pH 值略有下降,前者可能与肌肉自溶产物有关,后者是因为空气暴露会使蟹在血淋巴中积聚碳酸[26]。
1.7 混合应激源应激对水产动物肌肉品质的影响
水产动物在运输过程中往往受到多种应激源影响,且应激反应有时间积累效应。随着运输时间的延长,牡蛎(Ostreidae)[26]、虾夷扇贝(Mizuhopecten yessoensis)[30]、团头鲂[31]、文蛤[13]等水产动物的储能物质如肌糖原、脂肪、蛋白质等逐渐被消耗以对抗应激,因而乳酸逐渐积累,pH值下降。然而,林恒宗等[26]发现牡蛎在无水、低温保活运输期间乳酸呈下降趋势,这归因于牡蛎耗氧呼吸作用占比增加,呼吸及能量代谢减弱,乳酸被分解,使得乳酸积聚减少。同时还发现苹果酸及琥珀酸含量随着运输时间的延长而增加,推测可能是因为牡蛎呼吸代谢在保活运输初期旺盛导致保温箱内产生大量二氧化碳,并在随后的运输过程中抑制机体呼吸,导致牡蛎组织中酸鲜味(苹果酸和琥珀酸)明显增加。
随着有水运输时间延长,在混合应激下团头鲂[31]和鲫[32]的L*和W下降,团头鲂剪切力下降[32],黄颡鱼(Pelteobagrus fulvidraco)硬度、弹性、胶黏性、凝聚性、咀嚼性、恢复性等质构指标均显著降低[33]。在运输过程中以上三种鱼肌肉逐渐变暗主要是由于当鱼体受到应激时下丘脑-垂体-肾间组织轴(HPI)被激活,诱导黑色素细胞刺激素释放增加;质地劣变主要与细胞凋亡有关[6]。然而,张玉平等[32]发现异育银鲫(Carassis auratus gibelio)硬度和弹性随着运输时间的延长而增加,其原因可能与pH变化有关。
在保活运输中牡蛎必需氨基酸EAA 和呈味氨基酸下降明显,其原因可能是其作为底物被用于合成应激胁迫下所必需的化合物[27]。而闫丽新等[30]发现无水运输24 h 后虾夷扇贝的呈味物质(甘氨酸、丙氨酸、精氨酸)含量显著增加。在保活运输过程中,随着时间延长,牡蛎腺苷酸AMP、IMP 呈缓慢积累特征,这归因于长时饥饿及低温胁迫导致牡蛎机体代谢发生显著变化,ATP、二磷酸腺苷ADP 发生降解,最终导致AMP 与IMP 的积聚[27]。然而,在有水运输中团头鲂肌肉中AMP含量逐渐下降,IMP含量先下降后增加,HxR和Hx含量逐渐增加[31]。在运输应激中,随着时间的延长不同水产动物的氨基酸和ATP 衍生产物未呈现出稳定的变化规律,甚至出现研究结果相反的现象,可能存在两方面原因:一方面不同水产动物物种间代谢存在差异;另一方面这些物质是代谢中间物质,在整个运输过程中一直处于动态变化中。在运输过程中,随应激累计时间的延长牡蛎脂质氧化程度加剧,挥发性异味物质(如2-乙基呋喃、2-戊基呋喃、二甲基硫、吲哚、1-戊烯-3-醇等)呈缓慢积聚特征[26]。运输中应激积累还会影响水产动物的微生物组成。闫丽新等[28]发现牡蛎无水运输1 d 后细菌总数显著上升,通过高通量测序确定交替假单胞菌属(Pseudoalteromonas)和希瓦氏菌属(Shewanella)为牡蛎中的优势微生物。
2 减缓应激对运输后水产动物肌肉品质的影响
在运输过程中,通过采取减缓应激的措施(如加盐、暂养、加抗氧化剂、休眠和冷驯化等)可提高水产动物的存活率和增加运输距离。近年来国内外研究发现,减缓应激在提高存活率的同时,还会影响水产的肌肉品质(表2)。

表2 减缓应激对运输后水产动物肌肉品质的影响Table 2 Effects of alleviating stress on muscle quality of aquatic animals after live transportation
2.1 加盐对水产动物肌肉品质的影响
加盐(常用NaCl)是减缓运输过程中应激反应的常见措施,有助于维持水产动物的渗透压平衡,同时刺激鳃上皮黏液的分泌。添加氯化钠盐度在5~25范围内时,大黄鱼(Larimichthys crocea)应激反应减弱,肌糖原含量增加,乳酸含量降低[34]。然而ROSA 等[35]发现添加氯化钠(8、12 g/L)对减缓应激并无效果,还会降低肌肉品质,表现在嗜冷需氧菌增加、a*值和感官评分降低;这可能是因为添加NaCl 浓度过高,不利于鲇(Rhamdia quelen)体内的渗透压平衡,使宰杀后尸僵现象提前,对肉质产生负面影响。
不同水产动物在运输过程中的适宜氯化钠浓度存在差异,应该对添加的氯化钠浓度进行优化,确定最佳氯化钠添加浓度,以保障水产动物存活率和肌肉品质。
2.2 暂养对水产动物肌肉品质的影响
暂养能够使水产动物提前排空肠道,减少运输时的排泄废物,延缓水体氨氮浓度的升高速度,同时还可以去除水产动物体内的致病微生物。在保活运输过程中,与未暂养净化组相比,经暂养的虾夷扇贝肌糖原、蛋白质和脂肪含量较高,菌落总数下降,说明暂养能降低保活运输过程中的应激反应[36]。随着暂养时间的增加,运输后团头鲂肌糖原含量逐渐降低[2]。这两种水产动物肌糖原变化趋势相反,可能与不同物种间的代谢差异有关。团头鲂的代谢速率高,其在暂养过程中因饥饿应激导致肌糖原被大量消耗,在运输过程中因应激消耗的肌糖原减少,且运输后期其总肌糖原依旧呈现降低趋势。团头鲂的肌肉品质在暂养24 和48 h 时有明显改善,但当暂养时间延长到72 h 时肌肉品质有所下降[2],可以看出<48 h 的暂养,其间肌肉质量的提升主要归因于鱼类消化道残渣的排出,这能有效缓解鱼类在活运过程中的氨氮应激,使乳酸含量降低,色度、持水力和剪切力显著增加。然而,长时间的暂养会过度消耗储存的能量物质(如糖原),不利于抵抗运输中的应激,导致肌肉细胞破裂,L*、W、持水力、ATP末端衍生产物(IMP、HxR和Hx)降低。
2.3 加抗氧化剂对水产动物肌肉品质的影响
运输中常添加的抗氧化剂有抗坏血酸、核苷酸和植物提取物[12]。运输过程中水产动物应激后会产生大量ROS,造成氧化应激和细胞损伤,从而降低存活率和肌肉品质。研究发现,添加抗坏血酸和生姜汁等抗氧化剂可缓解运输过程中虹鳟[12]和草鱼(Ctenopharyngodon idella)[37]的氧化应激,从而改善肌肉品质,表现为L*、a*、b*、持水性增加,TVB-N 质量分数、菌落总数、K值和三甲胺浓度下降。
2.4 休眠对水产动物肌肉品质的影响
在休眠状态下,水产动物的代谢速率降低,可以缓解应激反应。冰温、麻醉剂(MS-222、二氧化碳)休眠是常见的休眠方法,可降低牡蛎[38]、大菱鲆(Scophthalmus maximus)[39]、石斑鱼(Epinephelusspp.)[40]的应激反应,使肌糖原、脂肪、蛋白质等能量物质含量增加,乳酸降低;同时色度、质构等品质改善,呈味氨基酸增加,TVB-N、硫代巴比妥酸TBA 值和细菌总数下降。
2.5 冷驯化对水产动物肌肉品质的影响
冷驯化可以使水产动物适应运输中的低温,避免出现冷应激。潘澜澜等[36]对虾夷扇贝的暂养水体进行3 ℃/h 降温,每降温3 ℃后对水体恒温6 h,完成水体梯度降温;冷驯化24 h 后,采用聚乙烯保温箱(3 L)加冰袋(250 mL)进行离水运输实验,发现冷驯化后,虾夷扇贝的应激反应减弱,肌糖原、蛋白质和脂肪含量增加,而菌落总数降低。
3 组学技术在运输应激反应影响肌肉品质变化机制分析中的研究进展
高通量检测技术、数据统计分析软件和数据库是组学技术的三大法宝[41]。基因组学、转录组学、蛋白质组学和代谢物组学是常用的组学技术,它们的检测对象分别为DNA、mRNA、蛋白质、代谢产物。近年来组学技术高速发展,其在水产动物应激机制[42]、食品安全和食品加工[43]中广泛应用,也逐渐用于揭示运输应激对水产动物肌肉品质的影响机制(表3)。

表3 组学技术在分析水产动物活运过程中应激反应对肌肉品质的影响机制Table 3 Appliaction of omics technology in analysis of the impact mechanism of stress response on muscle quality of aquatic animals during live transportation
3.1 转录组学
Wu 等[12]采用转录组学研究运输中热应激对虹鳟肌肉品质的影响机制,差异表达基因(DEG)结果提示,急性热应激对虹鳟体内碳水化合物代谢的影响可能归因于肌糖原减少。热应激后,虹鳟肌肉中谷氨酸、天冬氨酸等风味氨基酸和丝氨酸等甜味氨基酸的生物合成途径受到抑制,可能导致口感和质地劣变;进一步发现抗坏血酸可调节氨基酸的生物合成和糖酵解-糖异生过程以缓解热应激,从而改善虹鳟运输后的肌肉品质。
3.2 代谢组学
Peng 等[6]采用代谢组学技术研究高密度充氧运输过程中团头鲂鱼肉质地劣变的机制,发现运输6、12、24 h与3 h相比,样品间代谢物存在显著差异,其中有机酸及其衍生物上调,而核苷酸代谢物下调;团头鲂在活体运输过程中质地的恶化原因是氧化应激引起氨基酸、嘌呤、组氨酸和胆碱代谢途径的改变,从而导致肌肉细胞和纤维的破坏。
Wu 等[21]采用代谢组学技术探讨短时间运输过程中缺氧对虹鳟肌肉质量的影响及其通过复氧缓解应激反应的作用机制,结果表明,缺氧导致琥珀酸、黄嘌呤、腺嘌呤的含量上调和15-羟基二十碳四烯酸15(S)-HETE 含量下调,使剪切力、a*、饱和脂肪酸含量和呈味氨基酸含量下降;复氧促进甘氨酸和丝氨酸的生物合成,硫胺素和半胱氨酸的代谢,抑制琥珀酸的积累,从而缓解缺氧应激,复氧后肌肉品质能基本恢复到未受缺氧应激的水平;其结果表明,复氧24 h 是缓解缺氧引起的虹鳟鱼片质量劣变的有效措施。
3.3 蛋白质组学
Li等[44]采用定量蛋白质组学研究冷胁迫对克氏原鳌虾(Procambarus clarkii)肌肉蒸煮后硬度的影响机制,发现与未受冷应激对照组相比,在4 ℃冷应激24 h 的虾肉内检测到20 种上调的差异丰富蛋白(DAP),其中肌球蛋白轻链1(MLC1)和骨骼肌肌动蛋白6可能是克氏原鳌虾对冷应激反应中肌肉硬化的潜在标志物。这些DAP 表达的减少可能归因于低温下产生的活性氧对肌球蛋白MP的氧化降解。
3.4 联合组学
Lei 等[45]联合转录组学、蛋白质组学技术,研究模拟运输过程中低温冷应激对克氏原螯虾尾部肌肉质构变化的机制,基因本体论(GO)结果表明,克氏原螯虾应对冷应激的差异表达基因(DEGs)和差异表达蛋白(DEPs)主要参与代谢和糖酵解。京都基因和基因组百科全书(KEGG)代谢结果发现,对冷应激的代谢反应包括缬氨酸和异亮氨酸等氨基酸的变化。在较低温度下,机体通过激活糖酵解和氨基酸代谢途径以及过氧化物酶途径以维持身体平衡,细胞骨架蛋白肌动蛋白β-肌动蛋白和ACT1 等相关基因表达显著增加可能导致应激下肌肉硬度增加。Shi等[7]联合转录组学、代谢组学技术,研究拥挤应激导致克氏原螯虾虾尾肉品质劣变的机制,结果显示差异基因和代谢产物大多分布在氨基酸代谢、脂肪酸降解、三羧酸循环和碳水化合物代谢通路中;运输中拥挤应激会干扰虾尾肉中细胞膜结构的稳定性和渗透性、核糖体蛋白质的翻译和蛋白质折叠功能、脂肪酸代谢和降解、线粒体功能和能量代谢等,从而导致水分含量、持水性降低,硬度增加。
4 结语
运输过程中应激源众多,均会引起水产动物不同程度的应激反应。不同水产动物对相同应激源的耐受性差别较大,导致肌肉品质的变化差异大。在运输过程中,水产动物通过消耗能量来抵御应激,基因表达、代谢、免疫等都可能会发生变化,使得储能化合物、代谢物质和细胞形态等发生改变,并最终导致营养组成、质构、色度、蒸煮损失、风味等肌肉品质的变化。总结国内外研究发现,能量消耗是营养组成、滋味变化的直接原因,细胞凋亡是应激影响肌肉质构的主要原因。但运输后水产动物肌肉中氨基酸、持水性、风味的变化机制尚缺乏理论支撑。采用措施消除应激后,水产动物的部分肌肉品质能够恢复,然而关于消除应激对提升肌肉品质的机理方面研究还非常欠缺。未来应进一步加强对水产动物应激和肌肉品质变化的基础研究,为指导保水产品活运输提供理论依据。
