性别和发育阶段对绒毛白蜡光合特征及叶功能性状的影响
2024-01-25张碧嘉王泽锦樊大勇杨宝祥袁启华张新娜徐程扬
张碧嘉,于 淼,徐 晴,王泽锦,樊大勇,杨宝祥,袁启华,张新娜,*,徐程扬
1 北京林业大学 森林培育与保护教育部重点实验室,干旱半干旱地区森林培育及生态系统研究国家林草局重点实验室,城市林业研究中心,北京 100083 2 北京市西山试验林场管理处,北京 100095 3 永定河休闲森林公园管理处,北京 100010
叶片功能性状能够显著影响生态功能,并反映植被对环境的响应和适应对策。Wright等[1]提出的叶经济型谱中包括了净光合速率(Pn)、比叶面积(SLA)、叶氮含量(LNC)等六类叶片功能性状。研究发现植物叶经济型谱由快速投资-回收到缓慢投资-回收维度变化,反映了植物对环境适应的不同对策[2-4]。叶经济型谱在预测全球变化下生态系统结构和功能的变化[5-6]、恢复生态学[7]、入侵生态学[8-9]等各个生态学研究领域均有重要的研究价值。对于叶经济型谱的进一步研究表明,气候[3,10]、种间及种内关系[11-12]、基因变异[13]、环境胁迫[14-15]均显著影响叶经济型谱,反映在两两性状关系之间的斜率、截距和位移上。
然而,目前很少研究性别分化对叶经济型谱的影响。事实上,雌雄异株植物对维持物种多样性具有重要意义,是陆地生态系统必不可少的组成部分。相关研究表明,雌雄异株植物生殖资源分配策略是否存在性别间差异尚无统一认识,绝大多数研究认为,雌性植株往往在繁殖过程中投入更多的资源,进而导致不同性别个体在叶片大小[16]、光合性能[17]、气孔导度和水分利用效率[18]等叶片功能性状上存在遗传差异。但不同研究结果也不尽相同,以光合能力和气孔导度为例,部分研究表明,雌性植株净光合速率普遍高于雄性,同时表现出更高的气孔导度[17,19-20],但也有研究发现雄性植株具有更高净光合速率和水分利用效率[18-19],这可能与不同的发育阶段相关。有研究表明,植物叶片化学元素在不同的发育阶段存在差异[21-22],养分在不同发育阶段具有的不同分配方式,主要是满足不同生长期对植物养分的需求。
雌雄异株植物绒毛白蜡(FraxinusvelutinaTorr.)具有适应性强、耐盐碱、耐水涝等优良特性,是盐碱地的重要绿化树种[23]。已有研究发现绒毛白蜡树种存在明显的性别分化,主要表现在叶片矿质元素含量等方面,已经成为影响林分健康及其生态功能发挥的关键问题[24]。光合能力、叶功能性状与树体的生长发育密切相关,在不同季节动态,植物对光需求量和资源分配存在性别间差异,探究性二态植物的光合特征及其叶功能性状的关系,对造林树种选择及树木精准管护具有重要意义。本研究以雌雄异株绒毛白蜡为研究对象,在不同繁殖活动时期对不同性别植株的光合特征和叶功能性状进行测定,旨在回答以下问题:(1)不同发育阶段绒毛白蜡的光合能力是否存在差异?(2)不同发育阶段绒毛白蜡叶功能性状是否存在性别间差异?(3)净光合速率与叶功能性状间的关系是否存在性别间差异?
1 材料与方法
1.1 采样方法
试验地点位于北京市海淀区温泉苗圃(40°3′N,116°10′E),在样地中标记9年生不同性别绒毛白蜡31株,其中雄树22株,雌树9株,雄树树高为(7.4±0.6)m、胸径为(13.4±1.1)cm,雌树树高为(7.3±0.4)m、胸径为(13.5±1.1)cm,雌雄样本植株树高、胸径无显著差异(T=27.32,P=0.15),生长状况良好。于2021年5月至10月进行采集,将整样本植株树枝条进一步划分为一级枝、二级枝和三级枝(图1)。每一个样本植株选择南向生长状况一致、健康无病虫害的二级枝3-5个,用标签进行标记。将发育阶段划分为坐果期(5-6月)、果实膨大期(8月)和果实成熟期(10月)3个时期,在每个发育阶段,选择晴朗无风的天气,在上午9:00-11:00期间采集标记好的三级枝的叶片进行生理指标的测定。
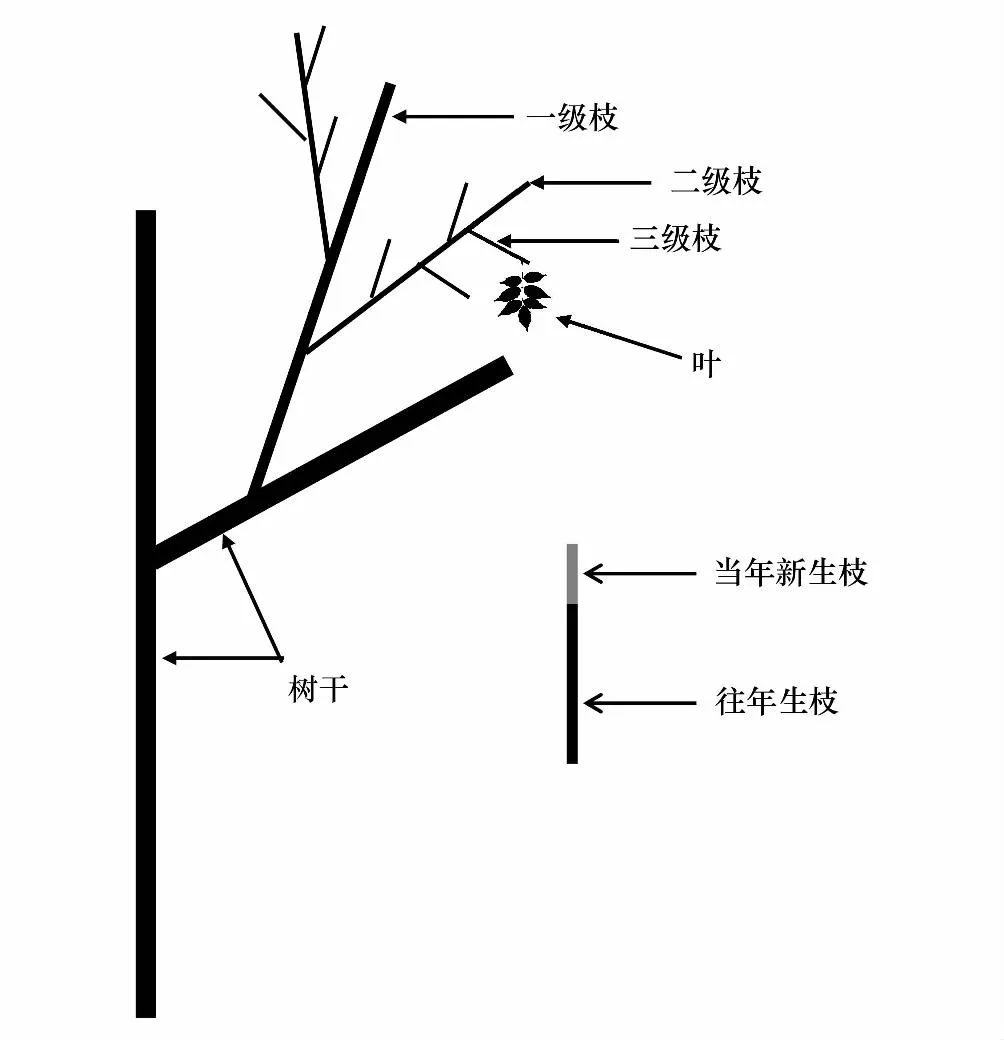
图1 枝条分级示意图Fig.1 Branch classification schematic
1.2 生理指标测定方法
1.2.1光合指标测定
在天气晴朗的上午9:00-11:00点,对每个发育阶段的31株绒毛白蜡,采用高枝剪剪取所标记的三级枝(2-3枝),即刻放入装满清水的水桶中,随后于水下剪去1/2长度枝条以防木质部栓塞导致的光合能力下降。预实验表明,在1200μmol m-2s-1饱和光强下2个小时光合速率、气孔导度没有下降,说明并未出现木质部栓塞现象。采用Li-6400便捷式光合测定仪(Li-Cor,Lincoln,NE,USA),选取苗木上的第5-6片(从顶部向下数)向阳完整功能叶片进行光合作用的测定,测定最新展开叶的净光合速率(Pn),测定时设置光强为1200μmol m-2s-1,相对空气湿度30%-50%,叶室温度为25℃,CO2浓度为(400±5)μmol/mol。当在此条件下达到稳态气体交换速率时,记录净光合速率(Pn)、气孔导度(Gs)、蒸腾(Tr)和细胞间CO2浓度(Ci)。瞬时水分利用效率(WUEi)为净光合速率与蒸腾速率之比。
CO2响应曲线的测定,保持光强和叶室温度条件不变,CO2浓度分别设置为400、300、200、150、100、50、400、400、600、800、1200、2000μmol/mol,每个浓度下稳定4-6min,稳定后记录数据。根据Long和Bernacchi模型[25]计算Rubisco的最大羧化速率(Vcmax)和最大电子传递速率(Jmax)。
1.2.2叶面积、叶干重测定
光合测定结束后,将部分最新完全展开叶取下带回实验室,利用Epson数字化扫描仪以及Win RHIZO图像分析软件测量叶面积(LA,cm2)。将叶片样品置于105℃下杀青30min,后在80℃下烘干至恒重(至少72h),称量叶干重(LB,g)。比叶面积(SLA,cm2/g)为叶面积与叶干重比值,比叶重(LMA,g/cm2)为叶干重与叶面积之比。
1.2.3叶绿素含量测定
将光合测定结束后的叶片,采用乙醇丙酮混合液提取法进行测定[26]。避开主脉将新鲜叶片剪成1-2mm宽的细条后混匀,准确称取0.1g于15mL离心管中并加入10mL乙醇∶丙酮=1∶1的提取液,将装有样品的离心管放置于黑暗环境中直到叶片完全变为白色。将提取液倒入比色皿中,分别在波长663、645nm下使用岛津UV-2600紫外可见分光光度计测定吸光值。每个样品重复测定3次。计算公式如下:
(1)
(2)
叶绿素总量:Chl(mg/g)=Chla+Chlb
(3)
其中,M为称取的叶片质量(g)。
1.2.4叶片全氮含量测定
将完全烘干的叶片样本用研磨仪将叶片粉碎,过100目筛(孔径为0.147mm),称取叶片5mg,采用“动态闪烧-色谱分离(改进杜马斯燃烧法),使用元素分析仪(FLASH2000 CHNS/O)测定样品全氮含量(LNC,mg/g)[27]。
1.3 数据分析
本研究数据采用双因素方差分析研究性别与发育阶段对光合特征、叶功能性状的影响,采用Pearson分析净光合速率与叶功能性状间的两两相关性。并针对净光合速率与叶功能性状间关系采用标准化主轴估计(Standardized major axis regression,SMA)分析。本研究涉及的所有数据均采用Excel 2016进行数据整理,使用R 4.1.2软件进行计算,其中“smatr”程序包用于SMA计算[28],使用Origin 2022软件进行绘图。
2 结果与分析
2.1 不同发育阶段雌雄个体的光合能力
绒毛白蜡的光合能力在不同发育阶段存在显著差异,在坐果期,光合能力偏低,在果实膨大期光合能力增强,而在果实成熟期下降,其总体趋势为先上升后下降。对于不同性别植株而言,在坐果期与果实成熟期,雄树的光合能力高于雌树,而在果实膨大期,雌树的光合能力偏高于雄树(图2)。
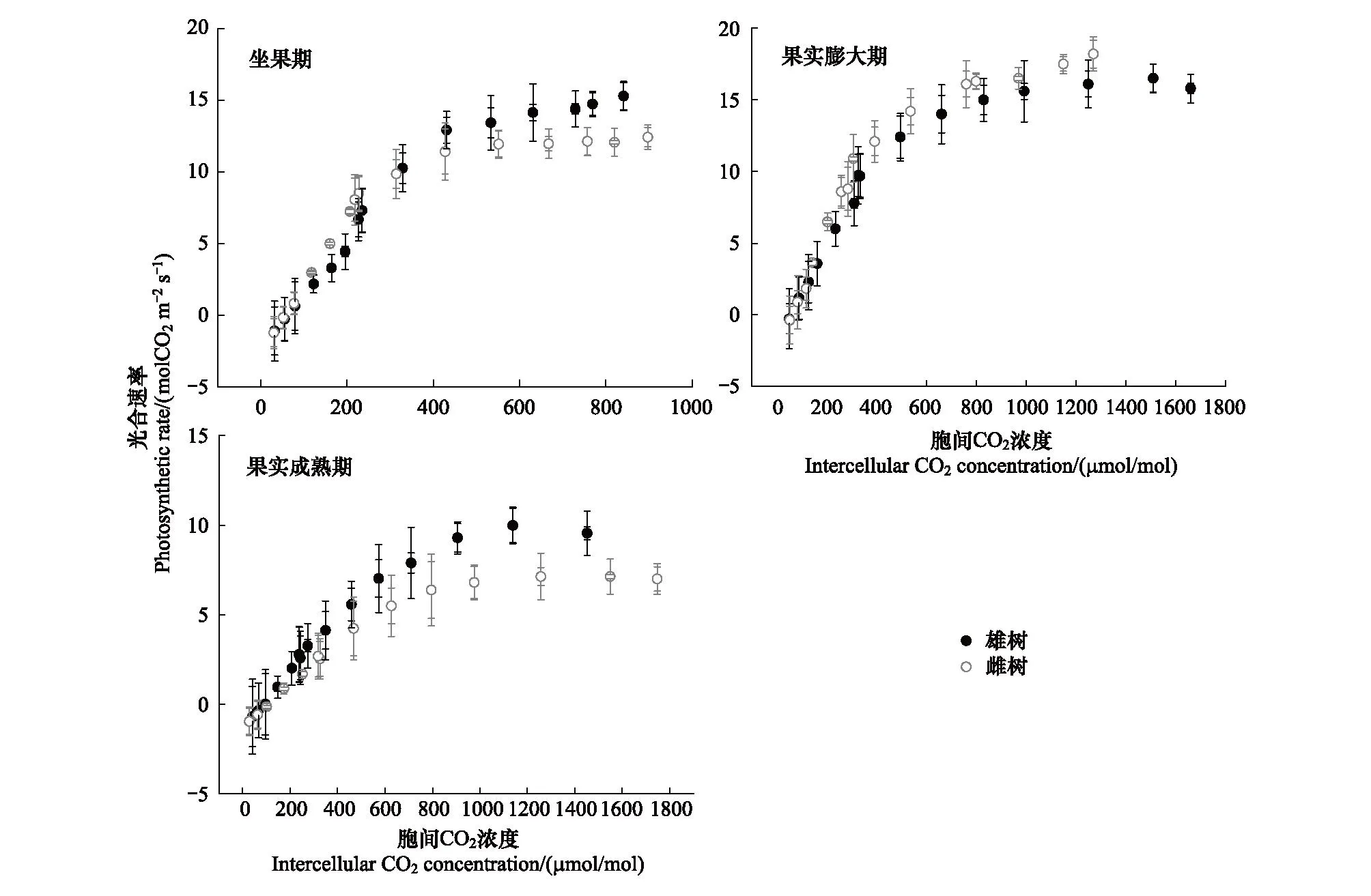
图2 绒毛白蜡雌雄植物在不同发育阶段的CO2响应曲线Fig.2 CO2 response curves of male and female plants of Fraxinus velutina Torr. at different development stages
由表1、表2可知,果实膨大期的净光速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、瞬时水分利用效率(WUEi)显著高于坐果期与果实成熟期。坐果期与果实成熟期,绒毛白蜡雌树的Pn、Gs、Ci、Tr、WUEi、最大羧化速率(Vcmax)和最大电子传递速率(Jmax)显著高于雌树,但在果实膨大期,绒毛白蜡雌树的Pn、Gs、Ci、Tr、WUEi、Vcmax、Jmax显著高于雄树。绒毛白蜡雌雄个体的Pn、Gs、Ci、Tr、WUEi、Vcmax、Jmax在发育阶段间均存在显著差异(P<0.05),但雌雄之间的Pn、Ci、Tr、WUEi、Vcmax、Jmax性别间无显著差异(P>0.05)。
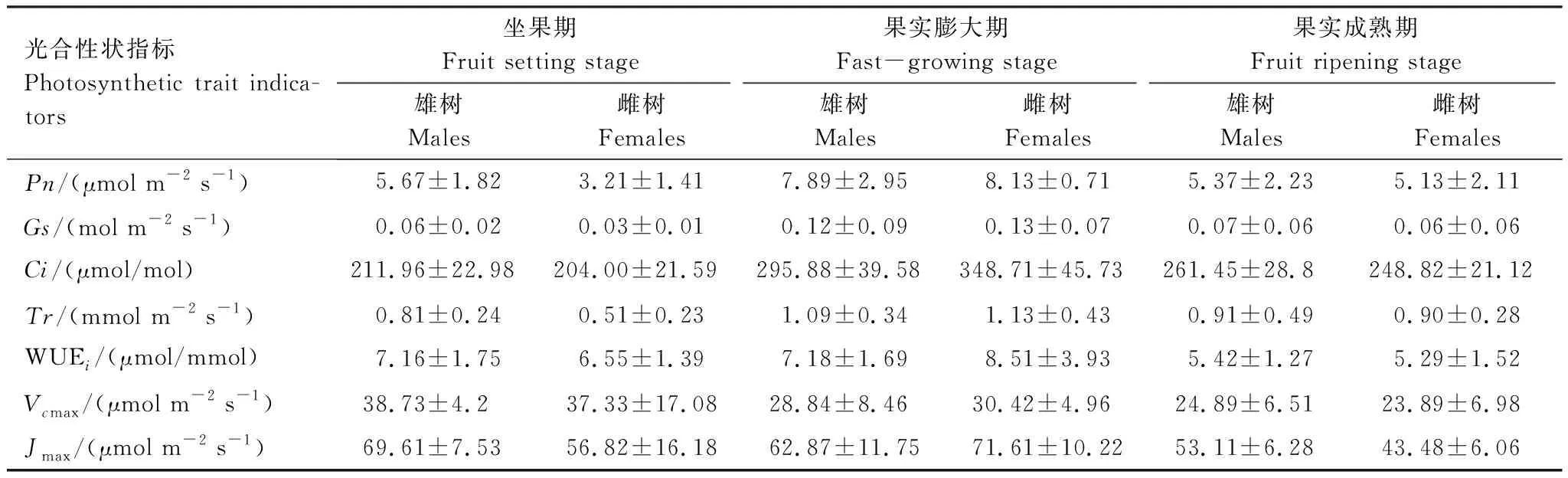
表1 不同发育阶段绒毛白蜡雌雄个体光合特征参数Table 1 Photosynthetic characteristic parameters of male and female plants of F. velutina at different development stages

表2 不同发育阶段绒毛白蜡雌雄个体光合特征的双因素方差分析Table 2 Results of two factor ANOVA on photosynthetic characteristics of male and female individuals of F. velutina at different development stages
2.2 不同发育阶段叶功能性状的性别间差异
如图3所示,雄树的叶干重(LB)和叶面积(LA)均大于雌树,且LB和LA在性别与时期间均显著差异,不同发育阶段叶绿素含量(Chl)和叶片氮含量(LNC)存在显著差异,而比叶面积(SLA)和比叶重(LMA)在性别与时期间差异均不显著。雄树的Chl在坐果期和果实成熟期大于雌树,且时期间差异显著,雄树的LNC在坐果期和果实成熟期高于雌树,且在性别与时期间均显著差异。

图3 不同发育阶段绒毛白蜡叶片生长指标Fig.3 Leaf growth indexes of F. velutina at different development stagesS1:坐果期Fruit setting stage.;S2:果实膨大期Fast-growing stage;S3:果实成熟期Fruit ripening stage;性别与时期间差异显著(P<0.05*,P<0.001***,P>0.05ns)
表3显示,雄树的Pn与LB、LA、Chl、LNC呈显著正相关(P<0.01);LB与LA、Chl呈极显著正相关(P<0.001),与SLA则呈显著负相关(P<0.001);LA与SLA、Chl、LNC呈显著正相关(P<0.05)。雌树的Pn与LB、LA、SLA、Chl、LNC呈显著正相关(P<0.05);LB与LA呈显著正相关(P<0.01),与SLA则呈显著负相关(P<0.01);LA与SLA、Chl、LNC呈显著正相关(P<0.01),Chl与LNC呈显著正相关(P<0.05)。其他各性状之间无显著相关性(P>0.05)。

表3 绒毛白蜡雌雄个体净光合速率与叶功能性状间的Pearson相关性(n=31)Table 3 Pearson correlation between male and female leaf functional traits of F. velutina
2.3 净光合速率与叶功能性状间的性别分析
SMA结果显示(表4),Pn在叶功能性状间的标准化主轴斜率存在性别间差异,雄树的LB斜率显著于雌树且大于1.0(P<0.001),LA、SLA、LNC的斜率大于雌树且小于1.0(P<0.05),雌树的Chl斜率显著于雄树且大于1.0(P<0.05)。

表4 不同性别植株净光合速率与叶功能性状间SMA回归结果Table 4 Summary of standardized major axis (SMA) regression results for the components examined in both genders
图4显示,绒毛白蜡雌雄个体的Pn与LB、LA、SLA、Chl和LNC均呈显著正相关(P<0.05)。结果表明随着净光合速率的增强,LB、LA、SLA、Chl、LNC含量在增大,其中雄树的LB、LA、SLA、LNC斜率大于雌树,说明雄树随着净光合速率增强的LB、LA、SLA、LNC增长速度大于雌树;雌树的Chl斜率大于雄树,雌树随着净光合速率增强的Chl增长速度大于雄树。
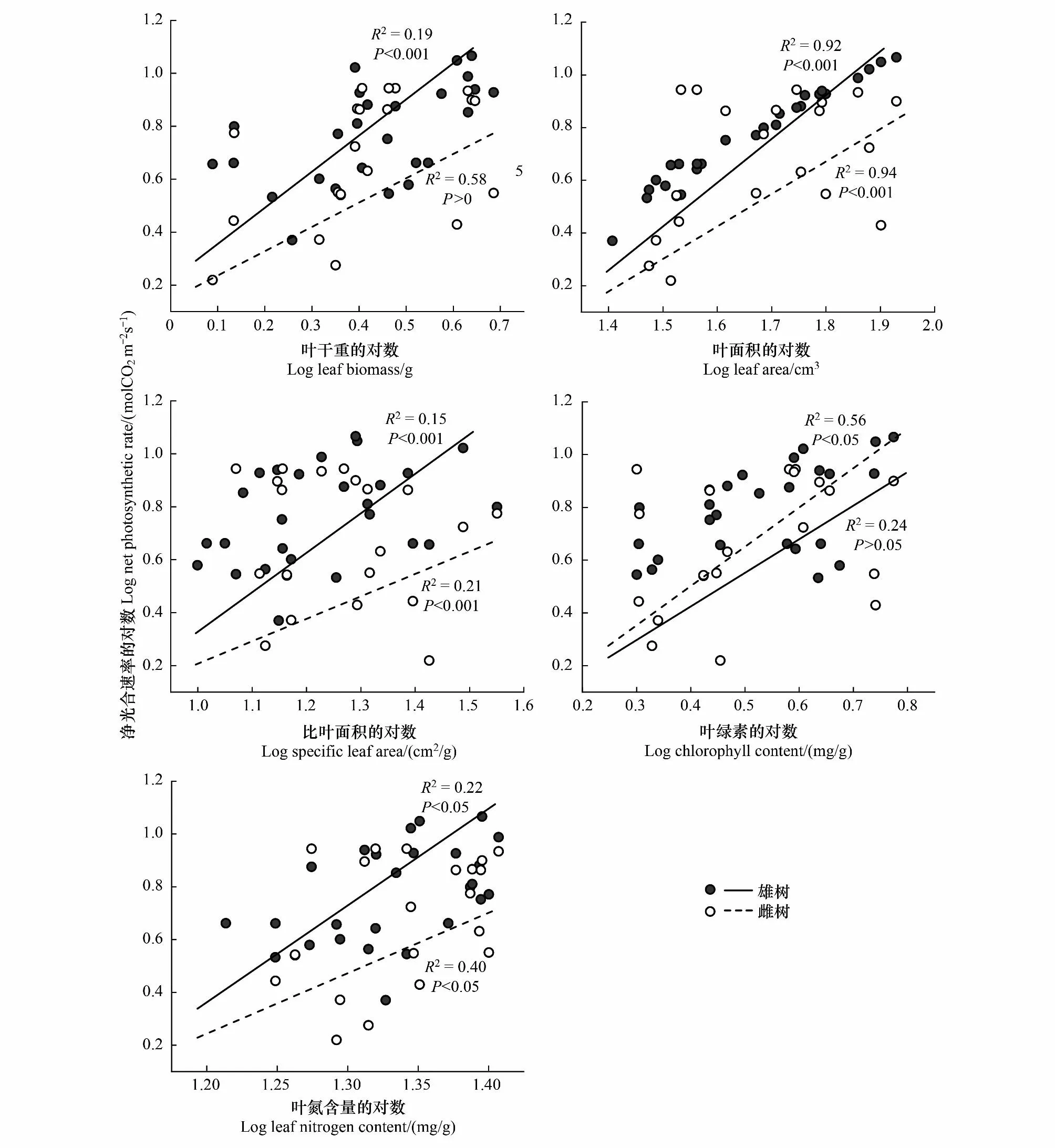
图4 绒毛白蜡叶片净光合速率(Pn)与叶功能性状之间的关系Fig.4 Correlation between net photosynthetic rate (Pn) and leaf functional traits of F. velutina leaves
3 讨论
研究表明不同发育期对雄树和雌树光合和叶片功能性状有显著影响。尽管交互作用对于大多数光合和性状参数影响不显著,但坐果期,雌树的光合能力显著高于雄树。进一步分析其不同发育阶段雌雄株的光合生理和特征后发现,雄树在坐果期和果实成熟期具有更高的Pn、Gs、WUEi、Vcmax、Jmax,这与多数前人研究结果一致[29-31],雌树在果实膨大期的Pn、Gs、WUEi、Vcmax、Jmax均大于雄树,这与马文宝等[32]在连香树的研究结果一致。在坐果期间,雄树Pn、Gs、Tr高于雌树,可能是因为雌树刚进行结实,将大部分资源用于繁殖过程,较少资源用于叶的光合作用过程,进而导致其光合能力低于雄树[33-34]。进入果实膨大期,雌树Pn、Gs、Tr高于雄树,表明雌树在果实成熟过程中,果实充当碳和营养汇的角色,发育中果实的强大碳汇增加了相邻叶片的光合速率,类似的发现在其他树种中也有研究表明[17,20,35-36]。而在果实成熟期,雌雄植株的Pn均下降,但雄树的Pn仍大于雌树,可能是雄树的气孔导度高于雌树而导致的,有研究指出净光合速率的降低是因为气孔导度的下降导致的CO2供应减少[37]。按相对生长理论的碳经济模型[38],相对生长速率是净同化速率(NAR)和叶面积比(LAR)的乘积。而LAR是比叶面积(SLA)和叶生物量比(LMR)的乘积,NAR的主要贡献者是光合速率(Pn)。就整个发育阶段而言,雄树和雌树在Pn和SLA上均差异不显著(表2,图3)。因此宋爱云等人所观察到的雌树长势较弱,抗逆性能较差[24]的原因,可能与LMR等生物量分配模式相关。Pn在性别间差异不显著也体现在Vcmax和Jmax上,两者共同决定了Pn[39-40]。就整个发育阶段而言雄树的Gs大于雌树(表1),暗示雄树对蒸腾水分散失的整体控制能力要低于雌树。两性之间WUEi无显著差异,出现这种矛盾结果的原因可能是瞬时水分利用效率不能反映长期水分利用效率[20,41],因此需要用稳定性碳同位素等研究方法进一步研究。有研究表明生长速率高的树种水分利用效率低[42]。
Cui等人发现[43],柳枝稷(PanicumvirgatumL.)在不同生育期的光合能力因叶片结构和生理调节而不同。雌雄异株植物的雌雄个体的光合能力明显因叶功能性状受到发育期的调节而不同,这表明随着生长发育阶段不同,叶功能性状会发生变化,雌雄植株对环境因子的响应也会发生变化[31,44]。在不同生长发育阶段,绒毛白蜡雌雄个体在叶功能性状上差异显著,雄树在坐果期和果实成熟期的LB、LA、Chl、LNC含量显著高于雌树(图3),雄树表现出更强的光合能力是因为雄树具有更大的叶片进行光资源的捕获[18],此外,LNC在一定程度上能够直接影响植物的光合能力[45];雌树则在果实膨大期的Chl、LNC含量高于雄树(图3),进而导致雌树在该发育阶段具有较强的生长和光合能力,可能是雌树在结果期增加更多的碳进而表现出更大的光合能力[46],可被认为是一种补偿更高生殖投资的机制[33]。可见,随着发育阶段不同,绒毛白蜡植物体内功能性物质会发生变化并存在差异。雌雄异株植物受性选择的影响,为满足生殖的特定需求具有不同的适应策略,造成性二态植物之间生理生态性状出现差异[19]。
叶功能性状与植物生物量及其对资源的获取和利用密切相关[47],且叶功能性状与光合作用之间的关联性是植物长期适应环境的结果。研究SMA结果发现(图4),LB、LA、SLA与Pn呈显著正相关(P<0.05),这与多数研究结果一致[48-49],进一步而言,本研究结果表明在叶功能性状与Pn之间关系上,雄树和雌树之间出现一定差异。有研究认为,SLA反映了物种的截光能力[50](单位资源投资的截光面积),因此SLA vs.Pn的关系表明了其光能利用效率,也就是单位质量的光合速率[51]。雄树在SLA vs.Pn上其斜率高于雌树,说明雄树的光能利用率高于雌树。叶绿素作为植物重要的光合色素,一定浓度范围内直接决定了植物光吸收能力和光合作用能力[52]。研究发现,雌树与雄树的Chl vs.Pn呈显著正相关(图4),但斜率两者之间差异不大,说明在光吸收率上并无性别间差异(P>0.05)。氮是植物生长的重要元素,叶片中氮含量常与叶片的光合速率呈正相关[53-54],两者间的关系反映了光合氮素利用效率[55]。绒毛白蜡雌雄树的LNC vs.Pn呈显著正相关(P<0.05),雄树的斜率大于雌树(图4),斜率越大植物单位叶片含氮量的光合速率越高,这表明雄树比雌树具有更高的光合氮素利用效率,显然,雄树与雌树在生物量和氮素分配上模式并不一致,雄树的光能利用效率和光合氮素利用效率高于雌树,表明雌树将更多的生物量和氮素用于非光合作用过程,这可能与其在生长-防御-繁殖的权衡关系调节相关。
不同环境中植物功能性状的差异,反映了植物个体获取环境资源的方式有所不同,因此植物采取的生活史策略会存在差异[56]。绒毛白蜡雌雄个体在光合特征与叶功能性状上存在差异,在不同发育阶段雌雄个体间差异显著,可能是绒毛白蜡雌雄个体在不同发育阶段的生活史策略存在差异,进而导致在不同发育阶段表现出不同的光合特征与叶功能性状差异。
4 结论
综上所述,绒毛白蜡雌雄个体在不同发育阶段的光合特征和叶功能性状存在差异,是由于两者的生活史策略不一样所导致的。在坐果期和果实成熟期,雄树表现出更高的生长和光合能力,可能是雄树具有较大的叶片,能够获取较多的光合资源用于生长,雌树则在果实膨大期表现出更高的光合能力,这可能有利于补偿雌树的生殖投入,从而维持其生长和生存能力。本文揭示了不同发育阶段下性别对绒毛白蜡光合特征和叶功能性状的影响,为进一步理解性别对性二态植物生长发育的影响机制提供理论依据。
