生态因子对三叶木通和五叶木通居群遗传多样性的影响
2024-01-18汤腾跃余柯达王佳淇范超洋
汤腾跃,余柯达,王佳淇,范超洋
(1.金华市婺城区雅畈镇人民政府,浙江 金华 321000;2.浙江师范大学,浙江 金华 321004;3.上海新纪元武义高级中学,浙江 金华 321017;4.金华先实大农业科技有限公司,浙江 金华 321004)
三叶木通和五叶木通同属木通科(Lardizabalaceae)木通属(Akebia),是营养价值和药用价值极高的藤本植物[1]。五叶木通属落叶或半常绿木质藤本,俗称木通,在我国野生资源丰富,是我国的传统中草药,具有抗癌、抗炎、抗菌、利尿、止痛、抗风湿等功效。三叶木通俗称八月炸、金肾果、野香蕉,果实是中药预知子。在我国分布广泛,其果实味香甘甜,风味独特,营养丰富,富含维生素C(VC)、17种氨基酸、矿物质、脂肪酸、蛋白质、糖和微量元素等,是名副其实的第3代保健型野生水果[2]。然而野生木通资源匮乏,人工培育显得尤为重要。
目前的研究[3-5]表明,温度是影响三叶木通生长的第一要素,光照是次要因素,海拔和坡度是决定三叶木通群落分布的主要因子,土壤性质对三叶木通的生长也存在着一定的影响。有关木通的遗传多样性研究相对较少,其中席在星[6]建立了三叶木通随机扩增多态性DNA(RAPD)最佳反应体系;田宗城等[7]用RAPD技术对木通属4个种进行研究,发现七叶木通与三叶木通种的亲缘关系较远,而白木通与三叶木通的亲缘关系较近;Kitaoka等[8]以三叶木通叶片为材料,建立适合三叶木通种质资源遗传多样性研究的优化AFLP反应体系。目前还没有关于生态因子影响木通遗传多样性的研究。
nrDNA的转录间隔区(internal transcribed spacer,ITS)承受的选择压力较小,具有很高的可变性,且在核基因组中高度重复。ITS区的可变性为物种鉴定提供了丰富的变异位点和信息位点。Zietkiewicz等[9]发现,五叶木通ITS序列不具有独立性。ISSR标记是1994年开始发展起来的一种分子标记技术[10],结合了SSR和RAPD的优点,目前已广泛应用于遗传多样性、进化及分子生态学等研究中。本实验结合ITS序列分析和ISSR标记2种方法探究生态因子对三叶木通和五叶木通遗传多样性的影响,为保护浙江三叶木通和五叶木通种质资源,提供基础资料和科学依据,也为三叶木通和五叶木通的人工培育提供了借鉴。
1 材料与方法
1.1 材料
本研究对浙江省三叶木通和五叶木通主分布区内的天然种群做了采样分析,包括金华北山(Bs)、开化古田山(Gt)、磐安大盘山(Pa)、泰顺乌岩岭(Wy)、杭州西天目山(Xt)、温州雁荡山(Yd)和丽水龙泉市(Zs)7个地区的三叶木通、五叶木通样本,共34个样品,各样地的三叶木通和五叶木通种群之间呈相对隔离状态。采样时,采摘三叶木通和五叶木通的嫩叶放于保鲜袋中,封口,置于样品贮藏箱(用冰块保持冷藏条件)带回实验室,-80 ℃低温冰箱保存。同时采集各种群距地表2~10 cm深的土壤,供土壤分析用。样品采集地点、样品编号等具体信息见表1。
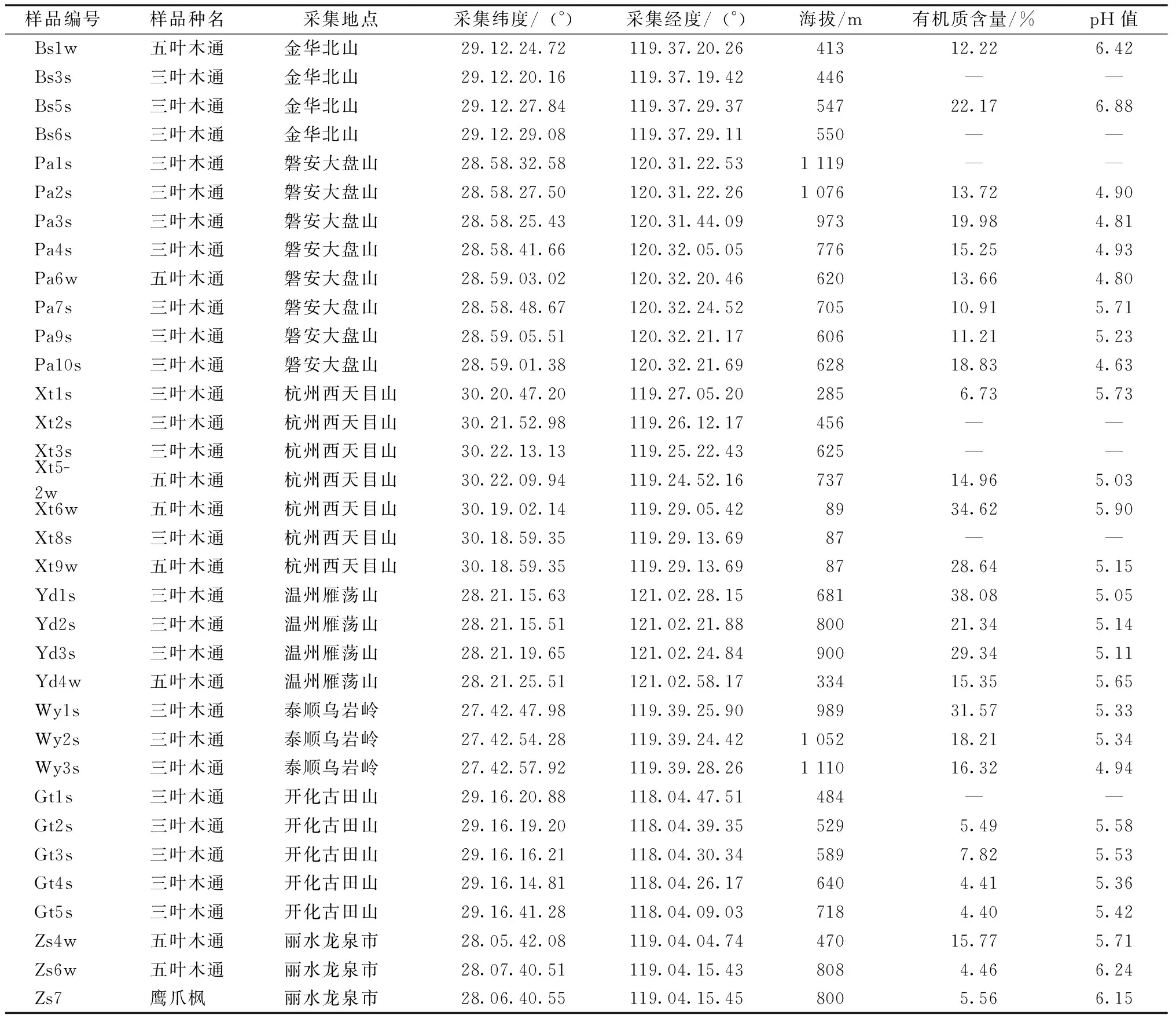
表1 样品采集具体信息Table 1 Sample collection information
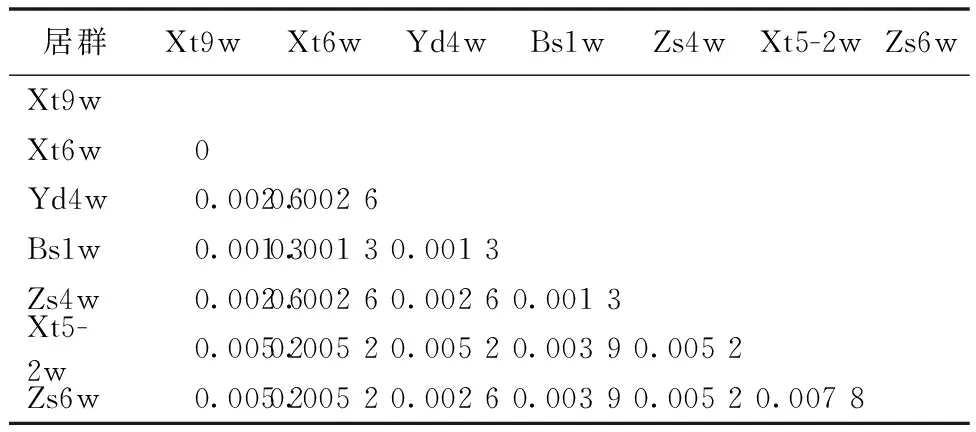
表2 五叶木通不同居群ITS序列的遗传距离Table 2 Genetic distance of ITS sequences in different populations of Akebia quinata
1.2 方法
1.2.1 基因组DNA提取
参考张铮等[10]改良CTAB法提取基因组DNA,用1%的琼脂糖凝胶电泳法检测提取的样本基因组DNA质量,并置于-20 ℃保存备用。
1.2.2 ITS序列扩增和测序
采用引物ITF1(5′-GCTCCTACCGATTGAATGGT-3′)和ITR1(5′-GTAAGTTTCTTCTCCTCCGC-3′),扩增木通样本的ITS序列。PCR扩增程序为:94 ℃预变性3 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,共30个循环。扩增反应体系为25 μL,含2.5 μL 10×ExTaqPCR Buffer、70 ng模板DNA、0.2 mmol·L-1dNTP、0.4 μmol·L-1ITF1、0.4 μmol·L-1ITR1、0.75 U ExTaqDNA聚合酶(宝生物工程大连有限公司)。PCR产物采用上海生工DNA柱式通用回收试剂盒进行回收,10 μL回收产物送至生工生物(上海)公司进行测序,对所有序列均进行双向测序。
1.2.3 ISSR扩增及电泳分析
根据加拿大哥伦比亚大学公布的ISSR引物序列设计,由上海生工生物工程技术服务有限公司合成。对木通属ISSR-PCR反应体系进行了优化,确定了最佳的反应条件为:反应体系为25 μL,其中2.5 μL 10×PCR Buffer(Mg2+Plus)、70 ng模板DNA、0.15 mmol·L-1dNTP、引物浓度0.4 μmol·L-1、0.5 mmol·L-1Mg2+、0.75 U rTaqDNA聚合酶(宝生物工程大连有限公司);PCR扩增程序为:94 ℃预变性4 min,94 ℃变性30 s,退火45 s(退火温度随引物而定),72 ℃延伸90 s,共35个循环。PCR产物用2%琼脂糖凝胶进行电泳,用凝胶成像分析系统拍照保存。
1.3 数据分析
1.3.1 ITS数据分析
对测序公司返回的ITS序列,用ClustalW 2软件进行序列比对。序列使用MEGA 5.0分子进化遗传分析软件构建发育树,发育树各分支的置信度用自举检验法检验,共进行500次循环,以评价各分支的系统学意义与可靠性。
1.3.2 ISSR数据分析
在电泳图谱上的每一个条带均视为一个分子标记,代表1个引物的结合位点。按凝胶同一个位置上DNA带的有无进行统计,有带的记为1,无带的记为0,生成0/1 Excel统计表。采用POPGEN 32软件计算多态位点百分数(P)、Shannon′s信息指数(I)和Nei′s基因多样性指数(h)来估算基因多样性[11]。利用SPSS 16.0软件导入0/1 Excel统计表计算样品遗传相似系数和遗传距离,并按UPGMA法(非加权配对算术平均法)对所有样品进行聚类分析,构建亲缘关系图。
2 结果与分析
2.1 ITS序列分析
2.1.1 ITS聚类分析
用最大似然法(ML法)构建ML系统发育树(图1),用bootstrap自举法检验发育树,重复500次,判断各分支处的可信度。
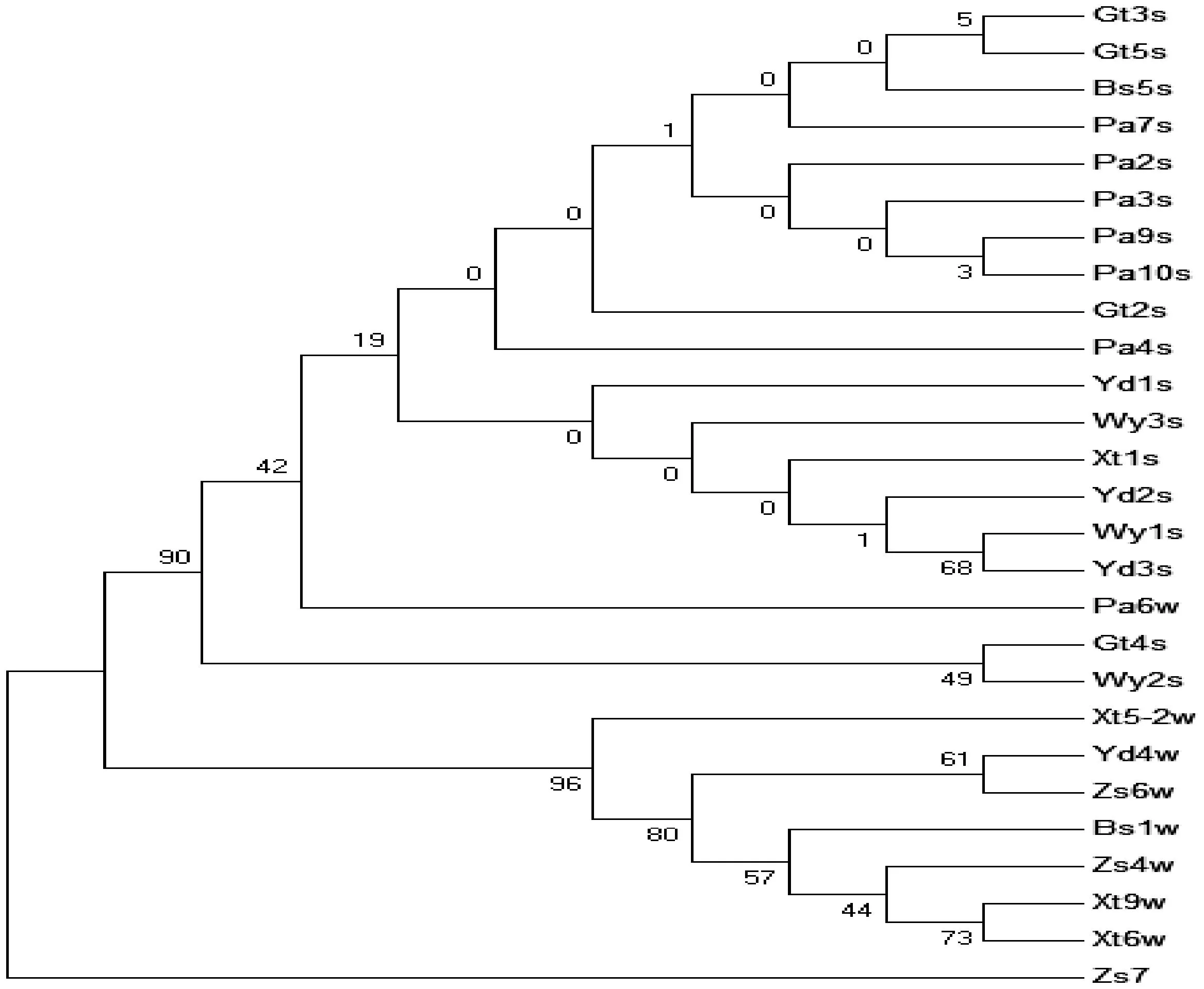
图1 基于ITS序列构建的27个样本ML系统发育树Fig.1 A phylogenetic tree of 27 samples of ML based on ITS sequences
在聚类图中,三叶木通和五叶木通大致各自聚为一支,而Pa6w样本与三叶木通聚为一类,在三叶木通一类中与其他样本遗传距离较远。由叶形观察知,Pa6w样本为五小叶复叶,而叶片大小、叶缘和长宽比与三叶木通相近,因此推断,Pa6w样本可能是三叶木通和五叶木通的过渡类型。从ML聚类图看出,三叶木通样本间的聚类结果支持率十分低,其聚类结果可信度不高,说明不同地区三叶木通的ITS序列间变异不大,遗传距离过小,不适合采用ITS序列来分析它们之间的亲缘关系。相反,五叶木通的聚类结果支持率较高,可以采用ITS序列来分析样品之间的亲缘关系。
2.1.2 海拔与五叶木通遗传多样性具有明显相关性
五叶木通某些样品无法采集土样,未能测定其pH值和有机质含量。利用MEGA 5.0软件计算遗传距离(表2)可知,五叶木通不同居群间ITS碱基序列的遗传距离在0~0.007 8,与其他物种相比,种内ITS序列的变异范围较小。在ML系统发育树中,Xt6w和Xt9w的海拔高度差距最小,两者之间的遗传距离也最短,而Xt5-2w虽然也采自西天目山地区,但并没有与Xt6w和Xt9w聚为一支,可能是由于其海拔较高。计算海拔与样本遗传距离间的相关系数,得出r海拔=0.96,远大于0.8,相关性显著。总体而言,随着居群之间海拔高度差距的增加,其遗传距离也相应增大,说明海拔对五叶木通的遗传多样性产生一定的影响。
2.2 ISSR分析
2.2.1 ISSR引物筛选结果
通过ISSR技术对三叶木通进行遗传多样性分析,从75对引物中筛选出15对扩增条带多、清晰且重复性好的引物用于ISSR-PCR反应(表3)。经统计,这15对引物扩增共得到265条带,其中多态性条带为261条,占比98.49%。扩增的DNA片段长度介于200~2 000 bp。
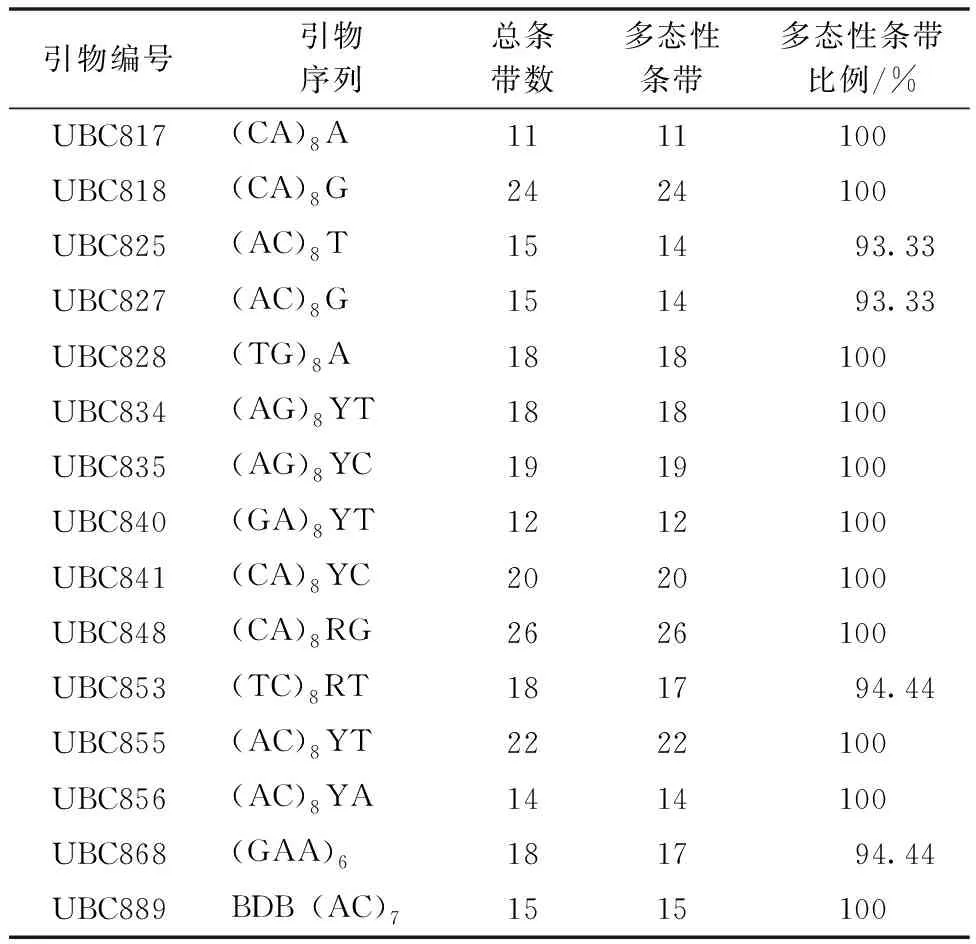
表3 筛选出的15条ISSR引物Table 3 Fifteen screened ISSR primers
2.2.2 三叶木通遗传多样性
各地区样本的多态位点百分率存在着差异,古田山地区最高,北山地区最低,变化范围在49.26%~70.32%,平均为61.42%。各地区三叶木通的Shannon′s信息指数(I)变化范围在0.212 2~0.605 9之间,平均为0.394 0。Nei′s基因多样性指数(h)的大小为0.250 0~0.367 3(表4)。P、h和I都显示出三叶木通样本具有较高的遗传多样性(图2)。
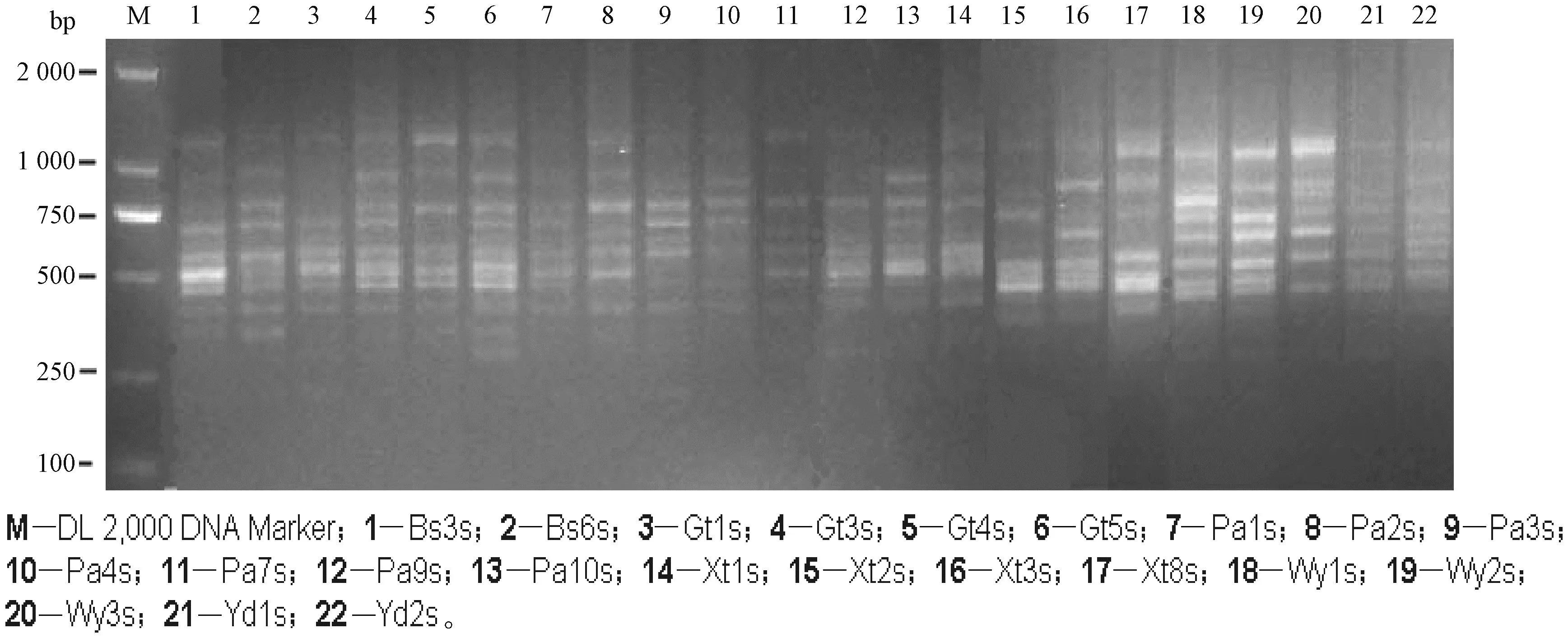
图2 UBC818 ISSR-PCR扩增电泳图Fig.2 ISSR-PCR amplification electropherogram of UBC818
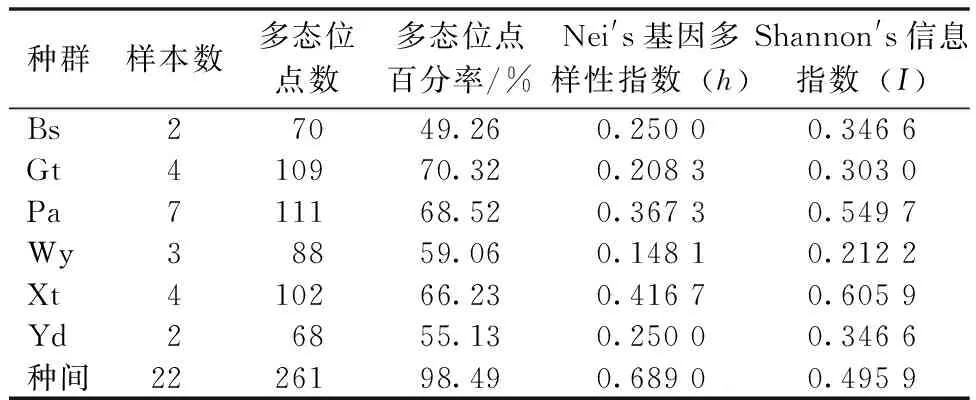
表4 不同地区三叶木通的遗传多样性Table 4 Genetic diversity of Akebia trifoliata in different regions
2.2.3 ISSR聚类分析
运用SPSS 16.0对6个地区的23个三叶木通样本进行UPGMA聚类分析,得到树状聚类分析图(图3)。从图3可以得出,当遗传距离为13时,每个地区的三叶木通样本各自聚为一类;当遗传距离为18时,乌岩岭、西天目山、雁荡山的样品和北山、古田山、大盘山的样品区分开来;当遗传距离为25时,三叶木通和参照样本(鹰爪枫)区分开来。这表明同一个地区三叶木通其遗传差异小于不同地区间三叶木通的差异。
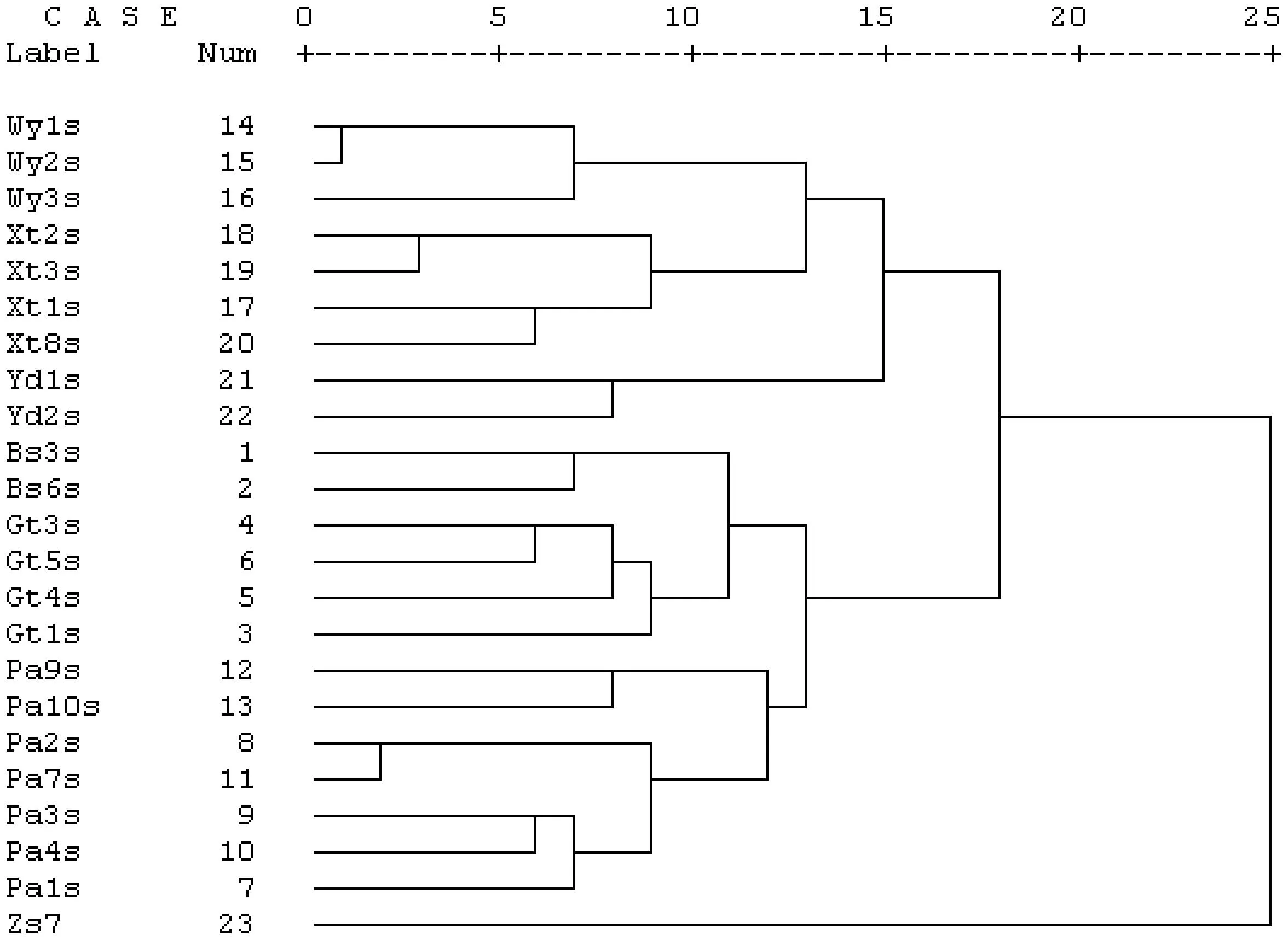
图3 23个三叶木通样本UPGMA聚类树状图Fig.3 UPGMA cluster dendrogram of 23 Akebia trifoliata samples
2.2.4 海拔、土壤有机质含量和pH值与三叶木通遗传多样性的相关性不显著
从三叶木通的聚类分析结果可知,三叶木通的聚类与区域有关,每个地区的三叶木通各自聚为一类。三叶木通样本取自浙江省6个不同的地区,所生长的生境有着一定的差异,结合生境中的土壤pH值、有机质含量和海拔这3个生态因子,计算三者与遗传距离的相关系数,得出r海拔=0.03,rpH值=0.22,r有机质含量=0.02,相关系数r远小于0.8,这表明海拔、土壤的酸碱度和有机质含量对三叶木通的遗传多样性影响不大。
3 讨论
3.1 海拔与五叶木通遗传多样性相关性分析
研究结果表明,海拔与五叶木通的遗传多样性有一定的相关性。五叶木通性喜温暖湿润气候,不耐严寒,多生长在海拔50~1 200 m的山地灌丛、林缘和沟谷中,其中又以海拔200~900 m分布较为丰富[12],推测该海拔高度范围较适宜五叶木通生长,受到的生境选择压力较小,从而影响其遗传多样性。
海拔因素对五叶木通遗传多样性的影响,初步说明了海拔对基因交流存在一定的阻碍作用;而海拔相近的五叶木通居群,如Xt6w和Xt9w遗传距离较小,可能是小范围内生境类型大致相同造成的。由于海拔差距的存在,导致物种不同居群间亲缘关系发生变化的现象,在皮桦[13]和水稻[14]中也有报道。海拔的变化常常是决定物种生境差异的主导因子,可能还伴随着温度、降水、光照和土壤等生态因子的改变,进而影响植物的分布和遗传多样性[15]。
3.2 地区差异与三叶木通遗传多样性相关性分析
利用ISSR技术对浙江省6个不同地域的三叶木通种群遗传多样性进行分析,其物种水平的多态位点百分率(P)高达98.49%,Nei′s基因多样性指数(h)为0.503 3,Shannon′s信息指数(I)为0.689 0,反映出三叶木通的遗传多样性较高。影响种群遗传分化的因素很多,如繁育系统、分布范围、种子传播机制和演替阶段等[16-19]。在UPGMA方法构建的系统树状图中,各三叶木通间是按照浙江省内的金华北山、开化古田山、磐安大盘山、泰顺乌岩岭、临安西天目山和温州雁荡山6个地区进行聚类的,说明三叶木通间的遗传变异程度与地域分布有一定相关性,其原因可能是地理阻隔导致遗传组成发生了变异,地理环境是造成该植物遗传多样性的重要因素[20]。
