接种土著和外源AM 真菌对杜梨抗旱性的影响及其适应机制
2024-01-17栗晗江尚焘彭海英李培根顾长宜张金莲陈廷速徐阳春沈其荣董彩霞
栗晗,江尚焘,彭海英,李培根,顾长宜,张金莲,陈廷速,徐阳春,沈其荣,董彩霞✉
1 南京农业大学资源与环境科学学院,南京 210095;2 广西农业科学院微生物研究所,南宁 530007
0 引言
【研究意义】梨是我国第三大栽培果树,主要采用嫁接繁殖,其中杜梨是应用较广泛的梨树栽培的主要砧木类型,在北方分布广泛[1]。近年来全球极端天气频发,高温热浪及干旱并发,严重影响农业生产。梨树栽培中普遍依靠自然降水,但季节性干旱现象严重,影响树体健康和果品质量。AM(Arbuscular mycorrhizal)真菌是一种广泛存在于自然界中,能与大多数植物建立共生关系的真菌,其发达的菌丝结构扩大了根系吸收面积,促进植物对水分的吸收,提高植物抗旱能力[2-3]。有研究指出,人为引入新的菌根真菌易造成菌根真菌在植物根系内的竞争加剧,导致植物减产[4]。从梨根际土壤中分离土著AM 真菌,比较土著与商业化AM 真菌对梨苗的促生效果,探究其促生、抗旱机制,可在丰富我国果树AM 真菌资源库的同时,为梨菌根化育苗提供理论依据和有效途径,对开发菌根肥料和实现梨果绿色高效生产具有重要意义。【前人研究进展】AM 真菌可缓解干旱胁迫对植物的损伤,一方面,扩大根系水分吸收面积并提高植物水分利用率;另一方面,通过增强生理生化反应改善植物生长及耐旱性[5],如促进养分吸收、改善光合作用、增强抗氧化酶活、增加渗透调节物质含量等,有研究表明干旱可加剧菌根在磷吸收中的作用[6]。AM 真菌还可通过间接影响根际微生物功能增强植物抗旱能力,HOANG 等[7]研究表明干旱胁迫下接种AM 真菌增强了大豆根际葡萄糖的渗出,使根际微生物在缺水但仍活跃的根际热点区域中增加了磷的利用率。AM 真菌资源广泛,但果树AM 真菌研究较少,关于梨根际土壤AM 真菌的分离也少有报道。目前大多数研究多集中在商业菌种,例如,根内根孢囊霉Rhizophagus intraradices、摩西斗管囊霉Funneliformismosseae和异形根孢囊霉Rhizophagusirregularis等,这些属于广谱性外源AM 真菌。AM 真菌具有不同功能性,Funneliformis mosseae[8]、Rhizophagusirregularis[9-10]等菌剂已被验证可帮助宿主植物抵御干旱胁迫,具有抗旱能力。许多研究表明[11-12],土著AM 真菌比外源商业菌剂效果更好,且在一些园艺作物及育苗生产上,土著AM 菌剂效果优于商业菌剂[13-14]。【本研究切入点】目前虽然已有关于干旱胁迫下AM 真菌群落的研究以及抗干旱功能AM 真菌菌剂的报道,但外源AM 真菌与同源土著AM 真菌在抗旱中的作用以及宿主适应胁迫环境是否与特定AM 菌株丰度的变化有关亟待研究。【拟解决的关键问题】验证梨土著AM 真菌较外源AM 真菌是否对梨苗有更显著的促生和抗旱效果;在比较不同AM 真菌菌株抗旱能力的基础上,进一步探究抗旱能力强的AM 菌株在植株遭受水分胁迫时是否具有更高的丰度,筛选出对梨苗促生和抗旱效果较好的菌株并验证其功能的特异性,揭示AM 真菌应对干旱胁迫的适应机制,也为梨菌根化育苗的实际应用奠定基础。
1 材料与方法
试验于2022 年4—7 月在南京农业大学牌楼温室进行。
1.1 试验材料
供试AM 真菌菌种:摩西斗管囊霉(Funneliformis mosseae,BGC HUN03B,Fm)、根内根孢囊霉(Rhizophagus intraradices,BGC BJ09,Ri)、层状近明球囊霉CXL-18(Claroideoglomuslamellosum,OQ154984,Cl)和蜜色无梗囊霉YT-6(Acaulosporamellea,OQ154986,Am)。其中,Ri 和Fm 菌剂均购于北京农林科学院植物营养与资源环境研究所,为商业外源菌剂;Cl 和Am 菌剂分别为梨和桃根际土壤中分离并扩繁得到的土著AM 真菌。该两种菌种分离自南京市溧水区相邻果园,土壤类型一致,根据宿主来源的不同,将Cl定义为土著同源菌剂,Am 定义为土著外源菌剂。形态和分子鉴定结果见图1 和图2。接种物均为孢子、菌丝、根段和基质等混合物。
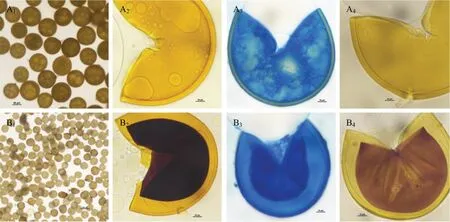
图1 梨和桃根际AM 真菌孢子形态Fig.1 Morphology of AMF spores in pear and peach rhizosphere
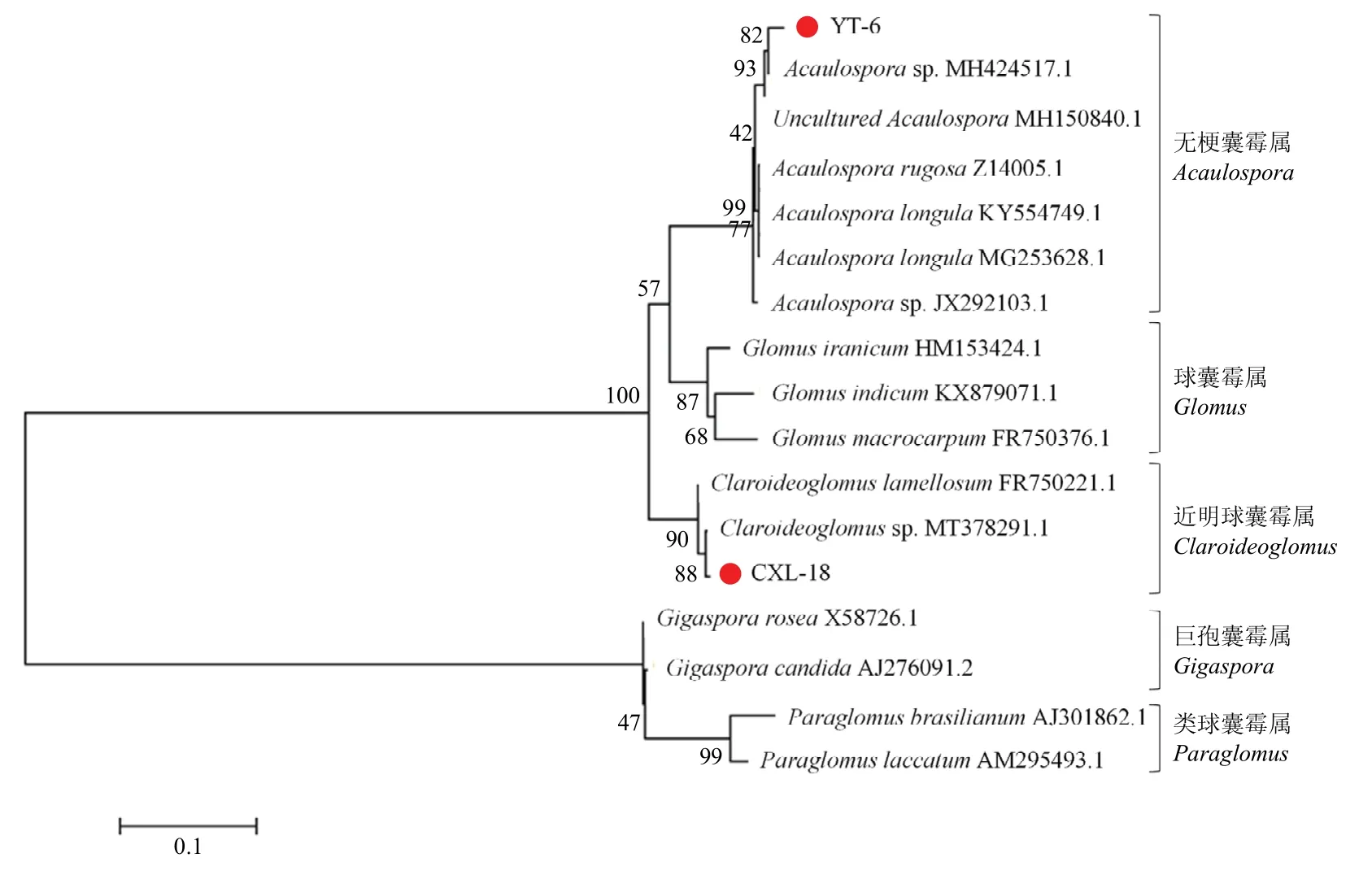
图2 基于18S rRNA 序列构建的系统发育树Fig.2 Phylogenetic tree based on 18S rRNA sequences
培养基质:土∶沙=1∶1(v/v),添加2%有机肥,充分混合后经121 ℃灭菌2 h 备用。供试土壤采自江苏省徐州市铜山梨园表层20 cm,其基本理化性质为:pH 8.66、有机质5.05 g·kg-1、铵态氮4.03 mg·kg-1、硝态氮12.55 mg·kg-1、速效磷40.32 mg·kg-1、速效钾164.63 mg·kg-1、有效钙3 376.24 mg·kg-1、有效镁273.49 mg·kg-1。有机肥生产商为淮安柴米河农业科技股份有限公司,N+P2O5+K2O≥5.0%,有机质含量≥45%。
以穴盘育苗2 周苗龄的杜梨实生苗为试验材料,选取生长一致的幼苗移栽于上口径11 cm、高13 cm的塑料花盆中,每盆1 株。
1.2 试验设计
设置6 种接种处理,分别为不接种处理(CK)、单接种处理(Ri、Fm、Am、Cl)及混合处理(4 种等孢子量混合,Mix);设2 个水分梯度,分别为正常与干旱胁迫。试验共计12 个处理,每个处理设置8 个重复。Mix 处理设置16 个重复,以供在不同时期破坏性取样。于2022 年4 月,选取生长一致的杜梨苗移栽至装有600 g 灭菌基质的花盆中。每盆添加菌剂于根部及根部下层位置,约1 200 个孢子/盆,不接种处理添加AM 真菌菌剂过滤液以平衡微生物的影响。所有幼苗在土壤最大持水量为75%条件下正常生长9 周后,进行正常与干旱两种水分处理,干旱处理土壤含水量设置为土壤最大持水量的35%左右,6 周后收获。处理幼苗置于南京农业大学人工气候室中培养,温度为15—30 ℃,相对空气湿度为30%—60%。
1.3 样品采集与分析
1.3.1 植物形态参数测定 收获时分别用卷尺和游标卡尺直接测定8 株杜梨幼苗的株高、茎粗,使用叶面积测定仪SYE-YM02 测定叶面积。将植株清洗干净,地上(茎、叶)、地下部分(根)分开,蒸馏水冲洗表面,吸干表面水分,置于烘箱中105 ℃杀青30 min 后于75 ℃烘干至恒重,称量地上部和地下部的干重。每个处理取3 份新鲜叶片参考AGANCHICH 等[15]的方法测定叶片相对含水量,采用乙醇提取-比色法测定叶绿素含量。菌根生长效应通过植株地上部生物量计算[16]。计算公式如下:
菌根生长效应=ln(XAMF/Xnon-AMF)
式中,XAMF:接种AMF 后植株地上部生物量(g);Xnon-AMF:不接种AMF 的植株地上部平均生物量(g)。菌根氮、磷、钾、钙、镁吸收效应计算方法同菌根生长效应。
1.3.2 植株元素含量测定 将烘干后的样品磨碎,采用H2SO4-H2O2消煮,流动分析仪测定植株氮(N)含量;采用混酸(浓硝酸∶高氯酸=4∶1)消煮,ICP-OES法测定植株磷(P)、钾(K)、钙(Ca)、镁(Mg)含量。每个处理3 次重复。
1.3.3 菌根侵染测定 将每个处理根系挖出,收集根际土并将根段洗净,选取新鲜根样,剪成1—2 cm,放入50%酒精溶液中固定,采用曲利苯蓝染色法进行侵染率测定。具体方法如下:加10%氢氧化钾完全浸泡根系,90 ℃水浴40 min 后用蒸馏水冲洗3—5 次;用10%双氧水常温脱色10 min,蒸馏水冲洗3—5 次;用2%盐酸常温酸化30 min,蒸馏水冲洗3—5 次;用0.05%曲利苯蓝(用乳酸甘油溶液溶解)90℃水浴染色20 min,蒸馏水冲洗3—5 次;置于乳酸甘油(乳酸∶甘油∶水=1∶1∶1)中常温放置24 h,水洗后进行制片。后续进行显微镜观察和AMF 侵染率的统计,采用侵染率加权法计算侵染率。菌丝密度采用真空泵微孔滤膜抽滤法,孢子密度采用湿筛倾析-蔗糖密度梯度离心法[17]。
1.3.4 抗氧化系统指标测定 取每个处理新鲜叶片,液氮研磨成粉,置于-80 ℃保存。使用试剂盒测定各处理间叶片中过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、过氧化物酶(POD)活性及丙二醛(MDA)、脯氨酸(Proline,Pro)、可溶性蛋白和可溶性糖的含量。MDA 含量用硫代巴比妥酸法测定,SOD 活性用WST-1 法测定,POD 活性用愈创木酚法测定,CAT活性用钼酸铵法测定,游离脯氨酸含量用酸性茚三酮法测定,可溶性糖含量用蒽酮比色法测定,可溶性蛋白含量用考马斯亮蓝法测定。所用试剂盒均购买于南京建成生物工程研究所有限公司,具体操作参考相应试剂盒说明书。每个处理3 次重复。
1.3.5 DNA 样品的采集与提取 干旱处理0、3 和6周时,分别取Mix处理新鲜根际土和根系样置于-80 ℃保存。分别使用Fast DNATMSPIN Kit 试剂盒和Power Soil® DNA Isolation Kit 试剂盒根据说明书从根系和根际土样品中提取总DNA。采用NanoDrop2000 核酸检测仪(NanoDrop Technologies,Wilmington,德国)对DNA 进行纯度检测,并通过1%琼脂糖凝胶电泳检测DNA 完整性。
1.3.6 DNA 样品的扩增与测序 采用高通量测序技术分析3 个不同时期正常与干旱条件下Mix 处理根际土与根系DNA 样品的AMF 基因。对于Illumina MiSeq测序,分别使用引物 GeoA2-AML2[18-19]和 NS31-AMDGR[20-21]通过两步PCR 获得18SrDNA 扩增子,使用AxyPrepDNA 凝胶回收试剂盒切胶回收PCR产物,Tris-HCl 洗脱;2%琼脂糖电泳检测。参照电泳初步定量结果,将PCR 产物用QuantiFluor™-ST 蓝色荧光定量系统进行检测定量,然后按照每个样本的测序量要求,进行相应比例的混合。后续进行Illumina文库构建和测序。具体测序方法参考BABALOLA等[22]。
Illumina 测序得到的PE reads 进行样本拆分后,首先根据测序质量对双端reads 进行质控和过滤,同时根据双端reads 之间的overlap 关系进行拼接,获得质控拼接之后的优化数据。然后使用序列降噪方法(DADA2)处理优化数据,获得 ASV(amplicon sequence variant)代表序列和丰度信息。使用AM 真菌标准序列及分子鉴定所得序列进行物种注释。测序过程交由上海美吉生物医药科技有限公司完成。
1.3.7 AMF gDNA 丰度的测定 AMF gDNA 丰度测定采用实时定量PCR 法(探针法)测定。考虑到实验室和菌种库条件,本试验仅对Fm 和Ri 两种AMF 的gDNA 丰度进行测定,具体方法参考李侠等[16]。根际土丰度测定方法与根系相同。
1.4 统计分析
采用SPSS 22 进行方差分析,采用独立样本t检验比较梨苗根系及根际土AMF gDNA 丰度在2 种菌种间及不同水分条件下的差异。采用GraphPad Prism 9进行简单线性回归及作图;采用RStudio 进行AM 真菌群落β多样性作图。
2 结果
2.1 不同水分条件下不同接种处理对杜梨幼苗长势及养分吸收的影响
2.1.1 对幼苗生长的影响 接种AM 真菌显著促进杜梨幼苗的生长。正常及干旱条件下单接Ri 及Mix接种均显著提高杜梨幼苗株高、茎粗、叶面积,促生效果最好,单接Cl 效果次之。单接Am 效果略优于CK,而单接Fm 与CK 效果相似。与正常条件相比,干旱条件下单接Ri、Am 及Mix 处理叶绿素含量分别增加26.61%、19.12%、26.56%,均高于CK处理(14.69%)。正常条件下,单接Ri、Cl 及Mix处理显著提高杜梨幼苗干重,分别提高41.76%、49.45%、52.20%,单接Fm 效果最差,表现为负效应。干旱条件下,单接Cl 及Mix 处理显著提高杜梨幼苗干重,分别提高35.26%、35.90%。正常及干旱条件下,不同接种处理均显著提高叶片相对含水量(表1)。
2.1.2 对幼苗养分吸收的影响 AM 真菌对植物生长表现有正效应、负效应、无效应。通过计算不同接种处理杜梨幼苗地上部菌根生长效应(图3-A)发现,单接Ri、Cl、Am 及Mix 处理均显著促进杜梨幼苗生长,但效果不一。而单接种Fm 在正常条件下表现为抑制杜梨幼苗生长,干旱条件下这种负效应消失。正常与干旱条件下,Mix 处理效果最好,单接Ri、Cl效果次之。
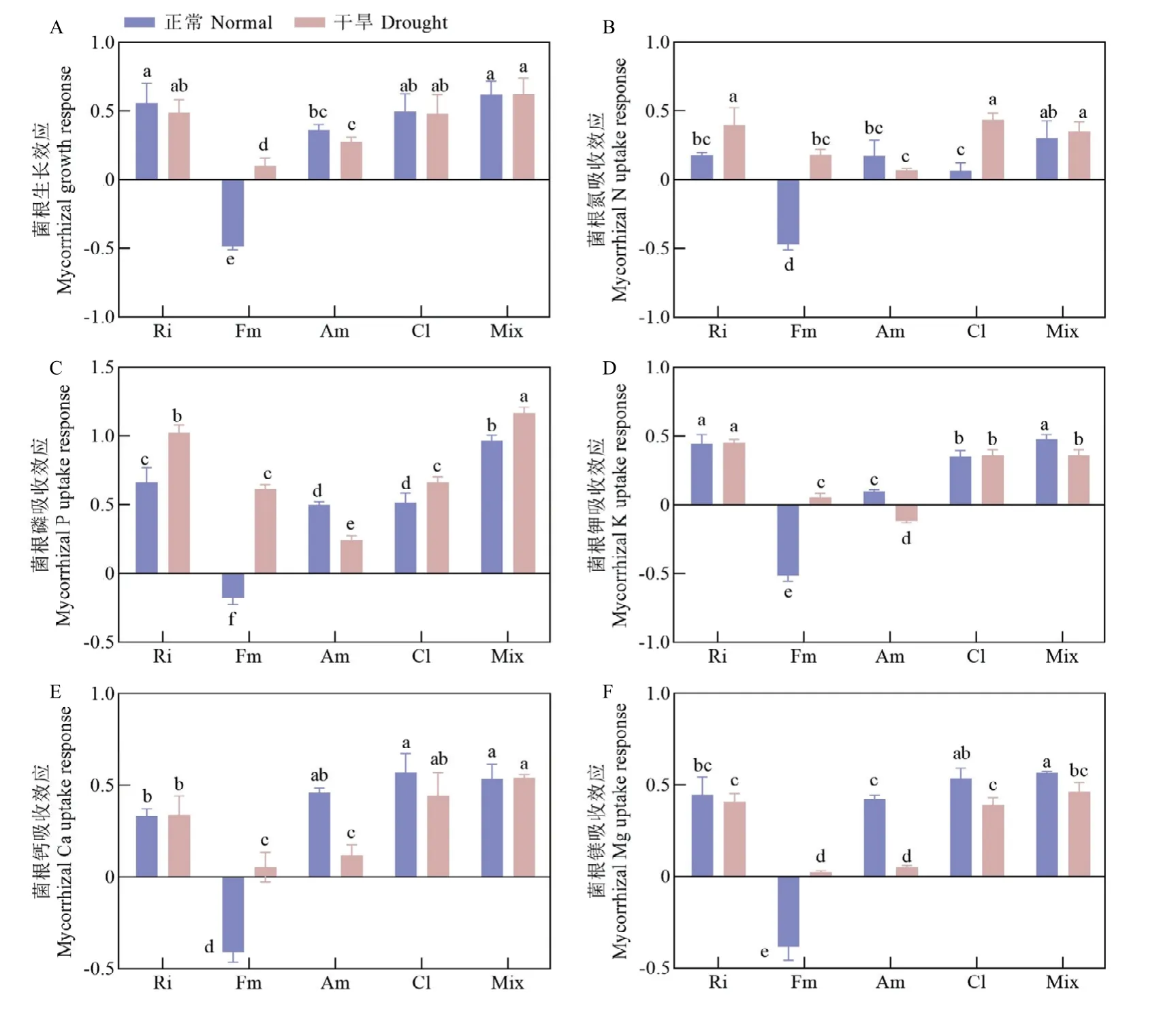
图3 正常及干旱条件下不同接种处理杜梨幼苗菌根生长效应(A)及氮(B)、磷(C)、钾(D)、钙(E)、镁(F)吸收效应Fig.3 Mycorrhizal growth response (A) and mycorrhizal nitrogen (B), phosphorus (C), potassium (D), calcium (E) and magnesium(F) uptake response of P.betulaefolia shoot inoculated with different AMF species under normal and drought conditions
通过计算不同接种处理杜梨幼苗地上部菌根元素吸收效应发现,与其他接种处理相比,正常和干旱条件下单接Ri、Cl 及Mix 处理显著促进杜梨幼苗地上部磷、钾、钙、镁等元素的吸收,尤其是磷的吸收,吸收效应最高达1.0 以上,而其菌根氮吸收效应仅在干旱条件下最显著。正常条件下单接种Fm 的菌根元素吸收效应均表现为负效应,而在干旱条件下负效应消失,甚至促进氮、磷元素的吸收。正常条件下,单接种Am 显著促进磷、钙、镁的吸收,干旱下表现为抑制植物钾元素的吸收,氮、磷、钙、镁元素吸收在接种处理间最低(图3-B—F)。
2.2 不同水分条件下不同接种处理对杜梨幼苗菌根侵染的影响
单接种Ri、Fm、Am 和Cl 及Mix 接种均对杜梨幼苗根系有较好的侵染。单接种Ri 和Mix 处理侵染强度及丛枝丰度在正常及干旱条件下均较高。干旱条件下,单接种Ri、Cl 侵染强度显著上升,其他接种处理无显著变化。与其他接种处理相比,干旱条件下单接种Cl 丛枝丰度显著上升,其他接种处理均有不同程度下降。不同接种处理在干旱条件下的菌丝密度均高于其在正常条件下的菌丝密度,且除单接种Am 处理外,其他均达到显著水平,Mix 处理菌丝密度最高,Cl、Ri 次之。但单接Fm 的菌丝密度增加幅度最高,为118%。除单接种Fm 外,其他接种处理的孢子密度在干旱条件下均显著下降(表2)。

表2 正常及干旱条件下不同接种处理杜梨幼苗菌根侵染情况、根外菌丝密度和孢子密度Table 2 Mycorrhizal infection, hyphal density and spore density of P.betulaefolia seedlings inoculated with different treatments under normal and drought conditions
2.3 不同水分条件下AM 真菌生长与功能的关系
AM 真菌侵染强度作为自变量,菌根生长效应和氮、磷、钾、钙、镁吸收效应分别作为因变量进行简单线性回归分析(图4)。正常条件下,回归方程分别是:菌根生长效应=侵染强度×0.020-0.325;菌根氮吸收效应=侵染强度×0.015-0.435;菌根磷吸收效应=侵染强度×0.023-0.214;菌根钾吸收效应=侵染强度×0.019-0.437;菌根钙吸收效应=侵染强度×0.013-0.100;菌根镁吸收效应=侵染强度×0.015-0.152。干旱条件下,回归方程分别是:菌根生长效应=侵染强度×0.013-0.076;菌根氮吸收效应=侵染强度×0.007+0.056;菌根磷吸收效应=侵染强度×0.020-0.009;菌根钾吸收效应=侵染强度×0.014-0.308;菌根钙吸收效应=侵染强度×0.012-0.127;菌根镁吸收效应=侵染强度×0.012-0.174。AM 真菌侵染强度与梨苗生长及元素吸收均具有较强正相关性,干旱条件下更为显著。
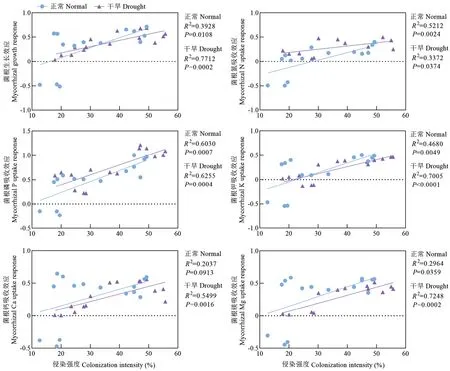
图4 正常及干旱条件下AM 真菌侵染强度与菌根生长和养分吸收效应的关系Fig.4 Relationship between AM fungal infection intensity and mycorrhizal growth and nutrition uptake response under normal and drought conditions
2.4 干旱胁迫下不同接种处理对杜梨幼苗抗氧化系统的影响
干旱胁迫使杜梨幼苗MDA 含量增加,但接种AM真菌处理的MDA 含量均显著低于CK 处理(图5)。干旱胁迫下,单接种Am 及Mix 处理均显著提高CAT活性,其他接种处理无显著影响;与CK 相比,单接种Ri、Fm 和Mix 处理显著提高POD 活性,单接种Am 显著下降,单接种Cl 无显著影响;单接种Ri 显著提高SOD 活力,其他接种处理无显著影响;与CK相比,单接种Ri、Cl 及Mix 处理显著提高杜梨幼苗叶片脯氨酸含量,可溶性糖含量无显著增加,单接种Fm、Am 处理对杜梨幼苗叶片脯氨酸含量无显著影响,可溶性糖含量显著下降。与CK 处理相比,单接种Fm、Am 及Mix 处理可溶性蛋白含量显著下降,单接种Ri、Cl 无显著影响。
2.5 不同干旱时期混合处理杜梨幼苗根系及根际土AM 真菌群落变化
通过基于根系与根际土AM 真菌组成群落水平的主坐标分析(CPCoA)发现,干旱处理与正常处理间的AM 真菌群落组成明显不同,根际土中更为显著,而3 周与6 周间AM 真菌群落组成未明显分开(图6)。

图6 正常及干旱处理根系(左)和根际土(右)AM 真菌β 多样性主坐标分析图Fig.6 Principal coordinate analysis of AM fungal β diversity in root (left) and root soil (right) during normal and drought treatments
不同时期杜梨幼苗根系和根际土AM 真菌群落组成不同(图7),但Ri 丰度占主导。如表3 所示,根系Ri、Cl 丰度变化显著受水分处理的影响(P=0.012、P=0.025),但不受处理时间影响(P=0.061、P=0.052)。在干旱处理6 周时根系Ri 丰度显著升高(P=0.040)。根际土Ri、Cl 丰度变化显著受水分处理(P=0.000、P=0.002)影响,Ri、Am 丰度显著受处理时间影响(P=0.032、P=0.030)。根际土Ri 丰度在干旱3、6周时均显著上升(P=0.002、P=0.025),Cl 丰度在干旱3 周时显著下降(P=0.009)。
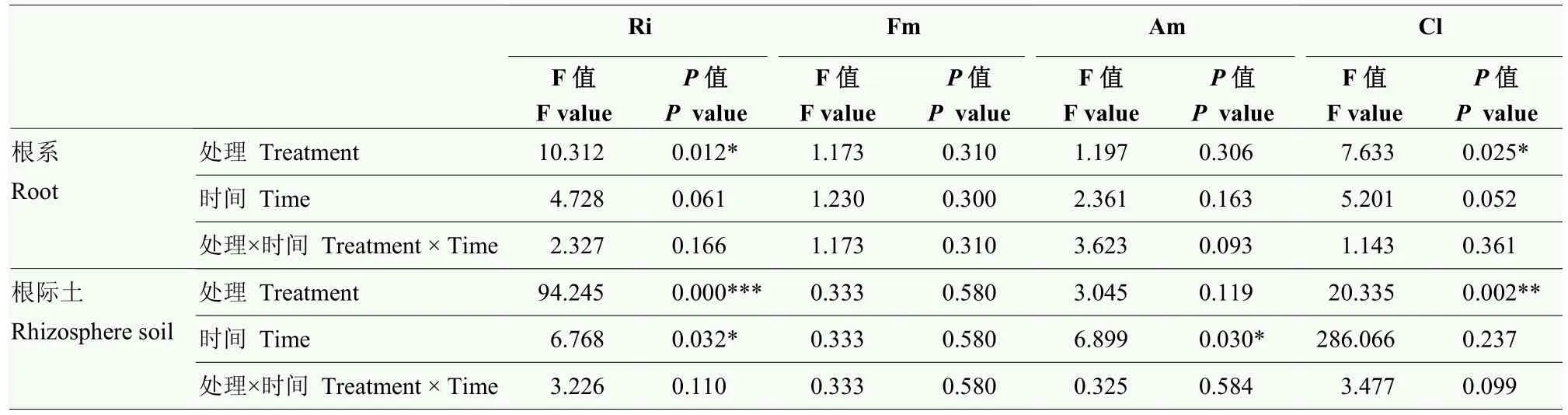
表3 干旱和时间对杜梨幼苗根系及根际土AM 真菌丰度的影响Table 3 Effects of drought and time on AM fungal abundance in roots and rhizosphere soil of P.betulaefolia seedlings

图7 正常及干旱0、3、6 周混合处理根系(A)和根际土(B)AM 真菌组成群落图Fig.7 Composition of AM fungi in root (A) and root soil (B) after 0, 3 and 6 weeks of normal and drought mixed treatment
通过对干旱6 周根系和根际土Ri、Fm 的绝对定量验证(图8)发现,Ri 丰度显著高于Fm 丰度,与高通量测序结果一致。干旱胁迫下,根系与根际土中Ri 丰度显著升高,而Fm 丰度无显著差异。

图8 正常及干旱处理6 周根系(左)和根际土(右)AM 真菌gDNA 丰度Fig.8 AM fungi gDNA abundance in root (left) and root soil (right) after 6 weeks of normal and drought treatment
3 讨论
3.1 干旱条件下接种土著AM 真菌影响杜梨幼苗生长及养分吸收
AM 真菌是分布最为广泛的一类真菌,在果园中也普遍存在。AM 真菌具有功能多样性,一方面可通过扩大菌丝面积促进宿主水分及养分吸收;另一方面调节宿主植物渗透代谢及相关酶活或基因表达等增强植物对逆境的耐性。本研究发现,除单接种外源菌剂Fm 处理外,正常及干旱胁迫下单接种AM 真菌及Mix接种处理均显著促进了杜梨幼苗生长,其中单接种外源菌株Ri、梨土著菌株Cl 及Mix 接种处理效果较好,外源菌株Am 效果其次,促生效果因AM 真菌种类而不同。王紫瑄等[23]发现对蒙古沙冬青幼苗单接与混接根内根孢囊霉(R.intraradice)和摩西斗管囊霉(F.mosseae)均显著增加植物株高、生物量。骆礼华[24]研究发现接种外源菌株F.mosseae对金佛山方竹的促生效果优于接种土著菌株A.mellea,ZOU 等[25]在甘草上也发现相似结果。C.lamellosum已被报道可显著提高柑橘砧木资阳香橙的株高、茎粗、叶片数[26]。前人研究结果表明菌株Ri、Fm、Am 和Cl 均具有促生能力,但促生能力不一,与本研究结果相似。植物-土壤反馈同时受到寄主物种和AM 真菌接种物的影响,接种同源AM 真菌的植株生长往往优于接种外源AM 真菌[27]。本研究中外源菌株Am 对杜梨幼苗的促生效果不如梨土著菌株Cl 及广谱性外源菌株Ri,外源菌株Fm 在正常条件下抑制杜梨幼苗生长,造成这种差异的原因可能是菌株对杜梨亲和性不同。另一方面,干旱胁迫下,接种处理对杜梨幼苗叶片相对含水量、叶绿素含量及相关抗氧化酶活性也有不同程度的提高,活性氧清除降低氧化损伤,渗透平衡及光合作用得到了改善,有利于梨苗抵御胁迫。AM 真菌还可改善植物营养。植物与AM 真菌间碳水化合物和矿质养分的物质交换是自然界物质循环的重要内容。已有研究表明,AM 真菌显著促进植物吸收N、P、K、Fe、Mn和Cu,以抵御干旱胁迫[28]。本研究中干旱下接种处理均不同程度地显著提高杜梨幼苗对元素的吸收,对磷吸收效果最显著,且菌根生长效应及元素吸收效应均与侵染强度呈正相关,与正常条件下相比,其相关性在干旱胁迫下均增强,表明在胁迫条件下AM 真菌的侵染有利于植物对养分的吸收。
AM 真菌与植物间的联系是相互的,也受环境影响。本研究中AM 真菌均对杜梨幼苗有较好的侵染,但侵染强度有所不同,外源菌株Ri 及Mix 处理侵染强度较强。已有研究表明在人为合成AM 真菌群落时,Rhizophagus属通常是最成功的植物根定植体[29]。与其他接种处理相比,干旱胁迫下单接种外源菌株Ri 和土著菌株Cl 的侵染强度显著上升,且土著菌株Cl 的丛枝丰度也显著上升,而AM 真菌菌丝一般是在土壤肥力良好的环境中形成较多的丛枝吸收养分,当肥力较低时菌丝以菌丝圈形式吸收和储存能量[30]。3 种处理在干旱下侵染情况不一,可能是不同AM 真菌种类与环境相互作用的结果,丛枝在细胞中周转也比较快。干旱胁迫下,接种处理根际菌丝密度均有显著上升,尤其是单接种Fm 处理。有研究表明,Funneliformis属可以在土壤中形成非常广泛的菌丝网络,可能桥接在干燥的土壤块间,促进矿物质营养的吸收及将水从远处土壤引向植物[31]。这也表明AM 真菌通过扩大菌丝面积帮助植物增加水分吸收,进而抵御干旱胁迫。干旱胁迫不利于AM 真菌孢子的繁殖。刘海跃等[32]发现沿荒漠植物花棒天然分布带由东到西随干旱程度增加,AM 真菌孢子密度逐渐降低,这与本研究中接种处理的孢子密度在干旱胁迫下均显著下降相一致。另外,不同AM 真菌孢子密度不同也侧面说明不同菌株存在不同步的生长周期。
AM 真菌具有特异性,它对植物的影响可从共生跨越到寄生。本研究中,单接种外源菌株Fm 对杜梨幼苗生长表现为负效应,但干旱胁迫下负效应消失。KAUR 等[33]用两种AM 真菌(Rhizophagusirregularis和Gigasporagigantea)分别接种高粱,也观察到了正面和负面效应。植物对AM 真菌接种的反应取决于环境。AM 真菌对植物生长的影响也因植物-AM 真菌特异性而异[27]。已有研究表明,F.mosseae的接种可显著促进干旱胁迫下小马鞍羊蹄甲[34]、胡杨[35]、青冈栎[36]等的生长,表现出其强大的抗干旱能力。本研究中Fm 菌株在正常条件下不利于杜梨幼苗生长,表现为寄生,而它在干旱胁迫下与植物转变为共生关系,能促进氮、磷的吸收,表明Fm 利于干旱胁迫下梨苗生长,只是当前土壤条件不利于其生长与繁殖。有研究指出植物和土著菌株组合的生长表现或者抗逆性显著优于植物和外源菌株组合[12],说明利用土著菌根真菌以促进植物生长比引入外来菌根真菌的效应更加稳定,也不易造成生态失衡。本研究中单接外源商业菌株Ri 的效果显著优于外源商业菌株Fm,梨土著菌株Cl 效果显著优于外源菌株Am,表明并非所有商业菌剂均适用于梨,土著AM 菌株的效果也不一定优于外源AM 菌剂,但同源土著AM 菌剂效果优于外源土著AM 菌剂效果。
3.2 梨苗AM 真菌群落对干旱的响应
亲缘关系较远的AM 真菌在功能上往往也具有较大的差异,因此与单一AM 真菌相比,不同AM真菌在植物根系定殖具有功能互补效应,帮助宿主植物应对不同环境的变化[37]。植物对不同AM 真菌的敏感性不同,宿主倾向更具合作性的AM 真菌提供资源[38]。本研究中,干旱胁迫下杜梨幼苗根系及根际土AM 真菌群落结构显著不同于正常条件,但外源菌株Ri 均占主导。SYMANCZIK 等[39]从沙漠中分离出4 种土著AM 真菌,以高粱为宿主,并引入外源菌株Rhizophagusirregularis,探究不同水分处理对AM真菌丰度的影响,研究发现在干旱条件下,引入Rhizophagusirregularis会降低根中所有土著AM 真菌物种的总丰度。EMERY 等[40]研究发现干旱胁迫显著改变了柳枝稷的 AM 真菌群落组成,主要是Rhizophagus类群丰度的增加。AM 群落功能的差异与某些AM 真菌物种的丰度有关[41]。菌株Ri 丰度较高一方面是其菌种特性决定的,另一方面因为它本身具有抗干旱能力,有利于梨苗干旱下的生长。本研究中,根系及根际土AM 真菌群落丰度均受干旱胁迫影响,干旱胁迫下外源菌株Ri 丰度显著上升,梨土著菌株Cl 丰度显著下降,而外源菌株Fm 所占丰度几乎没有,外源菌株Am 丰度无显著变化,通过定量发现根系及根际土中外源菌株Ri 丰度显著上升,Fm 丰度无显著变化。Fm 只在干旱处理前能被检测到,且在根系及根际土中丰度均较低,根据其在杜梨幼苗上单一接种结果分析,它在群落中竞争能力较弱,进一步印证该试验条件不利于其生长。梨土著菌株Cl 在单一接种时表现为明显的促生及抗旱效果,而在Mix 处理时效果不显著,可能是受到外源菌株Ri 的影响,干旱胁迫下Cl 对植物的效益不如Ri,植物对AM 真菌的选择更倾向于合作利益最大化的一方。NACOON 等[42]对洋姜分别接种分离到的4 种土著AM 真菌及从桉树根际土壤分离出的菌株A.mallea,研究发现A.mallea在正常和干旱条件下均未显著促进洋姜的生长和产量。Am自身对植物生长的影响较小,在杜梨幼苗上的促生能力也较弱,另一方面也因为它是外源土著菌株,对梨亲和性不强。根际土中AM 真菌群落结构还受到时间影响,随着时间的变化,外源菌株Ri 丰度逐渐下降,外源菌株Am 显著上升,这可能与无梗囊霉属AM 真菌在暖季产孢的物候特性有关[43]。综上所述,干旱胁迫下Mix 接种处理中AM 真菌群落发生了改变,群落中外源菌株Ri 更有利于植物抵御干旱胁迫,并且丰度占据主导位置。
4 结论
本研究通过梨苗促生盆栽试验,发现正常及干旱胁迫下,梨土著菌株Cl 和外源菌株Ri 及Mix 处理均显著促进杜梨幼苗生长,筛选出了对梨苗促生耐旱能力较好的菌株,而外源菌株Fm 与Am 对杜梨幼苗促生效果较弱,表现出AM 真菌促生及抗旱能力的差异。研究结果也表明并非所有商业菌剂均适用于杜梨,土著AM 菌株的效果也不一定优于外源AM 菌剂,但同源土著AM 菌剂效果优于外源土著AM 菌剂效果。高通量测序及绝对定量结果表明干旱胁迫下Mix 接种处理有利于杜梨幼苗生长,这与AM 真菌群落中利于抵御干旱胁迫的菌株Ri 丰度上升有关。
