生物强化促进餐厨垃圾高温厌氧消化产甲烷性能的研究*
2024-01-15杜学勋史晶晶张斯颖
杜学勋,史晶晶,张斯颖
(1.上海老港固废综合开发有限公司,上海 200237;2.中国科学院上海高等研究院,上海 201210)
0 引言
餐厨垃圾是居民在日常饭后所剩余的各类残渣的总称,也是城市生活垃圾的重要组成部分[1]。随着生活水平的提升,餐厨垃圾的产生量逐年上涨[2],预计到2025 年我国餐厨垃圾产生量将超过1.7×108t[3]。餐厨垃圾具有高含水量、高有机物含量的特点,如果不及时处理,极易腐烂变质、产生恶臭气味并滋生细菌和蚊虫,不仅会造成严重的环境污染,还会危害人类健康[4]。但同时餐厨垃圾也具有高有机质的能源属性,能够产生生物质能源[5]。因此近年来,如何高效处理餐厨垃圾,将餐厨垃圾的资源与环境属性相统一,在2060 年基本完成低碳能源转型目标成为研究的重点。
厌氧消化技术是目前处理餐厨垃圾的主流技术,主要由厌氧微生物经过水解、酸化、乙酸化、产甲烷这4 个阶段降解有机物,将其转化为清洁能源,实现废弃物再利用,是一种高效可行的技术[6]。然而餐厨垃圾的厌氧消化过程容易受到抑制,这是由于餐厨垃圾低C/N、高油高盐的特性导致微生物活性低,最终造成了厌氧消化效率低下甚至系统崩溃。而生物强化技术可以通过在厌氧消化系统中加入经过筛选、优化的优势功能菌种纯培养物,提高有机物的生物转化效率[7]。相关研究发现,加入具有一定功能的微生物能够在很大程度上提高厌氧消化的效率。在马缨丹(Lantana camara)厌氧消化体系中接种2 种纤维素降解菌Microbacteriumsp.DSB1、Arthobactersp.DSB12进行生物强化,甲烷产量分别提高了57%和60%[8]。有研究利用在生物炭上生长的嗜热甲烷单胞菌(Methanosarcina thermophile)对餐厨垃圾进行生物强化,甲烷产率最高增加了37%[9]。另外有研究发现将富集得到的耐丁酸微生物菌群(属水平主要包括Sporanaerobacter、Proteiniphilum、Syntrophomonas)加入到以餐厨垃圾为原料的酸化发酵罐中进行生物强化后,酸化系统得到明显恢复,产甲烷量(以VS 计)提高了52.2 mL/g[10]。因此添加功能微生物是提高厌氧消化效率切实可行的方式。
此前,实验室从餐厨垃圾的高温厌氧消化反应器中分离出1 株水原脲芽孢杆菌(Ureibacillus suwonensisE11),目前Ureibacillus suwonensis仅被报道在棉花废弃物堆肥的堆体中分离得到,其功能还未被研究[11],也没有将其应用在厌氧消化过程中。因此有必要探究从高温厌氧消化系统中分离得到的E11 对餐厨垃圾高温厌氧消化系统的生物强化作用。前期已经通过甲烷潜力测定实验证实E11 的添加可以有效提高餐厨垃圾的高温厌氧消化系统的甲烷产量,然而功能微生物的添加量是影响生物强化效果及成本的重要因素,故本研究考察功能微生物E11 对餐厨垃圾高温厌氧消化生物强化效果的影响,确定最佳剂量,并揭示生物强化的微生物机理,为生物强化提高餐厨垃圾的高温厌氧消化性能提供理论基础和指导。
1 材料与方法
1.1 底物与接种物
底物餐厨垃圾取自某单位食堂,主要包括餐后剩余的饭菜、废弃的菜叶、果皮和生肉等。所有原料经粉碎后于4 ℃储存。原始发酵污泥取自上海某湿垃圾处置单位稳定运行的餐厨垃圾中温(37 ℃)湿式厌氧发酵罐。采用一步式启动法驯化高温接种物,将厌氧连续搅拌反应器(CSTR)的运行温度设定至55 ℃,每天投加餐厨垃圾,其OLR(以VS 计)为1 g/(L·d),富集嗜热厌氧微生物。接种物取自实验室稳定运行3 个月以上的CSTR。实验开始前,对接种物进行饥饿处理,完全消耗接种物中的有机物直至不产气。底物和接种物的基本特征如表1 所示。
1.2 微生物培养
本实验生物强化使用的微生物取自实验室前期从餐厨垃圾高温厌氧消化反应器中分离出的土著微生物E11,鉴定为Ureibacillus suwonensis。在实验开始之前,将保菌管内的菌体稀释涂布至固体培养基上活化,放置55 ℃厌氧培养箱培养24 h。随后将单菌落接至试管中培养24 h 作为一级种子培养液,再按3% 的比例接至125 mL 的液体培养基中扩大培养24 h 得到菌液。
固体培养基:葡萄糖5 g/L,蛋白胨3 g/L,牛肉粉2 g/L,酵母粉3 g/L,NaCl 4 g/L,(NH4)2SO43 g/L,KH2PO40.75 g/L,K2HPO40.75 g/L,MgSO4·7H2O 0.3 g/L,FeSO4·7H2O 0.05 g/L,半胱氨酸0.25 g/L,琼脂粉15 g/L,自然pH。
液体培养基:葡萄糖5 g/L,蛋白胨3 g/L,牛肉粉2 g/L,酵母粉3 g/L,NaCl 4 g/L,(NH4)2SO43 g/L,KH2PO40.75 g/L,K2HPO40.75 g/L,MgSO4·7H2O 0.3 g/L,FeSO4·7H2O 0.05 g/L,半胱氨酸0.25 g/L,自然pH。
1.3 菌悬液制备方法
本实验设置实验组添加菌液的体积为CSTR 工作体积的5%、10%、15%和20%。为了排除菌液中培养基对厌氧消化的影响,本实验将不同体积的菌液在冷冻离心机中以6 000 r/min、15 min 的条件离心得到菌体沉淀,将不同体积菌液离心得到的沉淀分别重悬至100 mL 的无菌水中,得到不同浓度的菌悬液备用。
1.4 实验方法
在工作容积为5 L 的CSTR 中进行高温厌氧消化的甲烷潜力测定实验。所有反应器均在55 ℃下稳定运行,以60 r/min 的速度搅拌。厌氧消化的接种物和底物比例(ISR),基于VS,设定为2.0。为研究不同添加量条件下餐厨垃圾厌氧消化的产甲烷性能,本实验设置1 个对照组(CK)和4 个实验组(R1、R2、R3、R4),实验组对应的加菌浓度分别为5%、10%、15%、20%。CK 组加入100 mL 无菌水,实验组加入100 mL 菌悬液。在实验开始前,用氮气冲洗顶空15 min 以确保厌氧条件。在厌氧发酵过程中,每天记录发酵系统的甲烷产量、甲烷含量、pH。定期采集发酵液样品用来检测氨氮和VFAs。所有实验均设置3 组平行样。
1.5 分析方法
根据产气情况分别使用规格为10、5、1 L 的沼气采样袋(E-switch,上海)收集沼气,并通过沼气流量计(VIPPEN HR03,碧臣仪器,中国)测量甲烷体积。所有甲烷产量数据都转换为标准温度(273 K)和1 个标准大气压(101 325 Pa)下的体积。pH 通过pH 计(赛多利斯PB-10,德国)测定。气体成分(甲烷、二氧化碳)含量采用日本岛津气相色谱仪GC-2010 Plus 测定,热导池检测器(TCD);色谱柱为TDX-01(2 m×2 mm)填充柱,载气为高纯氩气,流量40 mL/min;柱温120 ℃,进样口和检测器温度150 ℃,电流20 mA,进样量1 mL。挥发性脂肪酸(Volatile Fatty Acid,VFA)采用日本岛津气相色谱仪GC-2010 Plus 测定,氢火焰离子化检测器(FID),色谱柱为Rtxwax(30 m×0.25 mm×0.25µm)弹性石英毛细管柱,载气为高纯氮气;检测器温度为230 ℃;进样器温度为220 ℃;程序升温80 ℃(保持2 min),升温速率20 ℃/min,最终温度200 ℃(保持8 min);进样量为0.3 µL。利用Nessler 试剂分光光度法分析总氨氮(Total Ammonia Nitrogen,TAN)。根据式(1) 计算游离氨氮(Free Ammonia Nitrogen,FAN)。
式中:T为温度,℃。
1.6 宏基因组测序分析方法
使用100 mL 的无菌注射器从反应器中收集后续用于基因组DNA 提取的样品,置于4 ℃,6 000 r/min、15 min 离心后分离沉淀和上清液,沉淀用液氮速冻后存放于-80 ℃冰箱,用于后续宏基因组分析。
全基因组DNA 提取利用E.Z.N.A.®Soil DNA Kit(Omega Bio-tek,美国)试剂盒进行样品DNA抽提。之前的研究中详细报道了双端文库构建、宏基因组测序生成、拼接组装和非冗余基因集构建的方法[12]。使用Diamond 软件(Version 0.8.35)将非冗余基因集的氨基酸序列与NR 数据库进行比对(BLASTP 比对参数设置期望值e-value 为1×e-5),并通过NR 库对应的分类学信息数据库获得物种注释,然后使用物种对应的基因丰度总和计算该物种的丰度。
2 结果与分析
2.1 功能微生物添加量对高温厌氧消化产甲烷性能的影响
通过添加不同比例的功能微生物对餐厨垃圾的高温厌氧消化进行生物强化,探究最佳的功能微生物添加量。产甲烷量是评价厌氧消化性能最重要的指标,不同处理组下餐厨垃圾厌氧消化的产气性能如图1 所示。接种后,R3 组的日产甲烷量在第1 天就达到峰值,其他处理组日产甲烷量均在第2 天达到最大值,随后呈整体波动下降的趋势(图1)。各组累积产气量持续上升,在第17天达到最大值,随后保持稳定。由图1 中可以看出,生物强化组的累积产甲烷量均高于CK 组。之前有研究指出,嗜热尿芽孢杆菌(Ureibacillus thermophilus)能够产生降解复杂有机物的裂解酶,进而有效地降解污泥[13-15]。根据推测,经E11 生物强化后的厌氧消化甲烷产量较高,这可能是因为E11 的添加能够产生降解有机物的活性酶,或者通过微生物之间的相互作用使其他功能微生物富集,最终表现在有机物水解更加彻底、甲烷产量更高。在生物强化组中累积产甲烷量随着E11 添加量的增加而增加,R4 组的产甲烷量最高,在第18 天的累积产气量(以VS 计)达到576.03 mL/g,相比于CK 组(452.86 mL/g)提高27.20%。当菌种添加量为15%时(R3),累积产甲烷量(以VS 计)略低于 R4, 为 575.14 mL/g, 相 比 CK 组 提 高 了27.00%。虽然R4 中E11 的添加量更高但是累计产甲烷量和R3 组几乎相同,这一现象与张新杰等[16]的研究结果相似,推测可能是由于功能微生物与本土菌株之间的竞争以及营养缺乏所致。结合产甲烷量的结果和经济性考虑,利用E11 进行生物强化的最佳添加剂量为15%。
2.2 功能微生物添加量对高温厌氧消化甲烷含量的影响
在厌氧消化的过程中甲烷含量是反映甲烷菌活性和消化性能的重要指标。在本实验中,不同功能微生物添加量对甲烷含量的影响如图2 所示。在初始阶段,各组甲烷含量先下降,随后增加,并在第5 天达到最高。在系统稳定运行的情况下,各组的甲烷含量差异不大,波动范围为50%~70%。相比于文献中报道的甲烷含量[17-19],本实验的甲烷含量均处于较高水平,可能是由于本实验的运行条件是55 ℃,在高温条件下水解产酸阶段进程更快、效率更高,使产甲烷菌有更多可利用的底物,从而提高甲烷浓度。各组的甲烷含量差异随着消化进程逐渐增大,CK 组整体的甲烷含量水平低于添加E11 的实验组,甲烷含量最大值出现在R3 中,达到73.53%。说明E11 的添加可有效提高甲烷产率,其中添加量为15% 时甲烷产生的效果最佳。
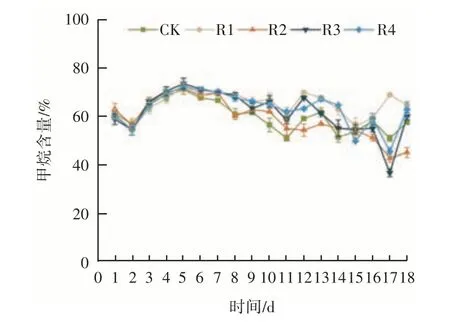
图2 功能微生物添加量对甲烷含量的影响Figure 2 Effect of bioaugmentation dosage on methane content
2.3 功能微生物添加量对高温厌氧消化pH 和乙酸浓度的影响
对于厌氧消化系统中的产甲烷菌来说,合适的pH 是非常重要的,pH 过高或过低都会直接影响微生物的活性,降低底物的转化效率,导致厌氧消化失败[12,20]。而VFAs 是厌氧消化过程中一种非常重要的中间产物,其中乙酸占比最高。乙酸是可以被产甲烷菌直接利用的一种物质,乙酸浓度过高会导致pH 的迅速降低,从而降低厌氧消化效率,但是浓度过低又会造成产甲烷菌可以利用的底物不足,导致产甲烷效率低。因此,pH和乙酸浓度是反映厌氧消化系统稳定性的重要指标。
厌氧消化期间pH 变化如图3 所示,pH 在所有组之间的变化趋势相似,每组的pH 均在实验开始的第1 天迅速下降,随着消化过程的进行pH 持续升高,第6 天后逐渐稳定,最终稳定在8.5 左右,这与He 等[21]的研究中由于VFAs 的转化和氨的积累,导致高温消化系统中的高pH 现象一致。第1 天pH 的迅速降低可能是由于底物的快速水解,产生大量的乙酸,产甲烷菌未将其及时转化。这一现象在Zhang 等[22]的文章中得到证实。乙酸在消化过程中经过产甲烷菌的代谢生成甲烷和其他产物,导致系统中的pH 升高。

图3 功能微生物添加量对高温厌氧消化系统中pH 和乙酸浓度的影响Figure 3 Effects of bioaugmentation dosage on pH and acetic acid concentration in thermophilic anaerobic digestion system
在厌氧消化过程中仅有乙酸和极少量丙酸被检出,由图3 中可以观察到乙酸浓度在不同组中的变化规律。所有组的乙酸浓度变化趋势相似,均在第1 天达到了最大值,随着消化的进行乙酸被快速代谢,浓度迅速下降,从第6 天开始趋于稳定。R1、R2、R3 组第1 天的乙酸浓度都比CK组高,但最后稳定阶段CK 组的乙酸浓度最高,并且R3 组的乙酸浓度在第9 天降为0,说明R3 组中的乙酸被完全利用,这可能是R3 组累积产甲烷量较高的原因之一。在其他生物强化组中甲烷产量的高低和稳定阶段的乙酸浓度成反比,说明添加E11 的生物强化措施可以有效促进乙酸被产甲烷菌利用,提高沼气产量。
综上,生物强化可以在一定程度上提高乙酸的利用率。当E11 的添加量为15% 时,乙酸被利用得最彻底。
2.4 功能微生物添加量对高温厌氧消化TAN 和FAN 浓度的影响
在餐厨垃圾的厌氧消化过程中,由于有机氮化合物的转化,氨氮含量和pH 会逐渐增加,当TAN 超过阈值浓度时会产生抑制作用。TAN 是FAN 和NH4+测量值的总和,从现有研究来看,FAN 似乎是导致氨氮抑制更为重要的因素,FAN可能直接抑制特定酶的活性或通过被动扩散进入微生物细胞,改变细胞外基质,随后改变钠钾平衡。在本实验中通过对TAN 和FAN 浓度的监测来评判高温厌氧消化体系的稳定性。不同生物强化剂量下厌氧消化系统TAN 和FAN 浓度的变化如图4 所示。图4 显示了每组TAN 浓度的变化,5 个反应器的TAN 浓度范围为880~3 660 mg/L。有文献报道称当TAN 浓度高于8 000 mg/L 时,通常会发生完全氨抑制[23],根据本实验反应体系中的TAN浓度推测该反应体系相对较为稳定,并没有明显的氨抑制现象发生。CK、R1、R2 组的TAN 浓度均在第1 天达到最低值,R3 组和R4 组在第2 天达到最低,随后各组TAN 浓度开始增加,从第6天开始基本稳定。此外,由图4 可看出,各组FAN 浓度波动范围为71~645 mg/L,所有组的FAN浓度变化趋势与TAN 的变化趋势基本一致。有研究指出高温厌氧消化系统中FAN 的浓度会高于中温厌氧消化,一般波动范围为400~800 mg/L[24],本实验系统中的FAN 浓度均处于正常范围,消化系统可以稳定运行。

图4 功能微生物添加量对TAN 和FAN 的影响Figure 4 Effects of bioaugmentation dosage on TAN and FAN
2.5 生物强化对高温厌氧消化微生物群落结构的影响
根据累积甲烷产量的数据并考虑经济性,E11 的最佳添加量为15%。为探究生物强化高温厌氧消化产沼气的微生物作用机制,本实验选择CK 组和R3 组消化达到稳定时(第7 天)的样品进行宏基因组测序,将得到的序列与NR 数据库进行比对,其微生物群落差异分析如图5~图7所示。

图5 功能微生物添加量对门水平微生物群落的影响Figure 5 Effects of bioaugmentation dosage on microorganism at phylum level
CK 组和R3 组在门水平上的微生物结构的差异如图5 所示,添加E11 后虽然E11 所在的厚壁菌门(Firmicutes)的相对丰度并没有增加,但是反应器中的热袍菌门(Thermotogae)、广古菌门(Euryarchaeota)和增效菌门(Synergistetes)微生物的相对丰度明显增加。热袍菌门在之前的研究中被报道是嗜热厌氧消化中的优势门,可以在高温下稳定存活并且降解糖产生CO2和乙酸,为产甲烷菌提供可以利用的底物。热袍菌门微生物的增加说明在添加E11 后糖类物质的降解更加彻底,从而提高甲烷的产率。广古菌门是厌氧消化系统中非常重要的一类微生物,包含能够产生甲烷的产甲烷菌,这可能是添加E11 后甲烷产率高的原因之一。在厌氧消化过程中蛋白质被水解为肽和氨基酸,这些肽和氨基酸随后被发酵为VFAs,最后被混合微生物种群发酵为甲烷和CO2,增效菌门已经被证明可以降解氨基酸[25],属于该门的微生物很可能在消化系统中的产酸阶段发挥重要作用。添加E11 后增效菌门相对丰度的提升说明厌氧系统的水解效率在一定程度上可能被提高,这与生物强化组的乙酸浓度较高的现象基本一致。VFAs在乙酸生成过程中的氧化通常发生在与产甲烷菌的协同共生关系中,产乙酸菌通常将醇类和VFAs分解成乙酸盐、甲酸盐、H2和CO2,这些物质随后被产甲烷菌利用[26]。因此,这类细菌的丰度增加可以提高甲烷产量。
从属水平(图6~图7)上看,Defluviitoga的相对丰度在R3 组中显著提高(p<0.05),Defluviitoga是厌氧消化过程中相对丰度最高的水解细菌,可以产生不同的碳水化合物活性酶,并利用多糖(如纤维素、几丁质和木聚糖)产生乙酸、H2和CO2,随后被产甲烷古菌利用产生甲烷[27]。Defluviitoga丰度的提升可以有效促进厌氧消化过程中底物的水解。Hydrogenispora同样是一种在厌氧消化系统中丰度较高的水解菌,这类微生物可将葡萄糖、麦芽糖和果糖等碳水化合物发酵成乙酸、乙醇和H2。这2 类关键水解菌丰度的提高表明生物强化系统中微生物对底物的水解能力加强,底物可以更加充分地被利用。醋微菌属Acetomicrobium是一类严格厌氧的耐高温细菌,可以将底物中的糖转化为乙酸盐、乙醇、CO2和H2,该菌属的增加有助于厌氧消化过程中有机质的分解和沼气的产生[28]。Syntrophaceticus是一类共养型乙酸氧化菌,可以将乙酸盐氧化为CO2和H2,该菌属是以互养乙酸盐氧化为主要产甲烷途径的产甲烷过程中的关键微生物,其相对丰度的增加对厌氧消化有促进作用。脲芽孢杆菌属Ureibacillus在R3 组中也显著增加,可以认为E11 添加后可以有效在消化体系内定殖。Methanoculleus的相对丰度在R3 组中也被发现明显升高(p<0.05),这是一类嗜氢产甲烷菌,其含有以H2和CO2为底物的产甲烷途径的编码基因,可以高效利用H2和CO2生成甲烷[29]。在产甲烷代谢途径中,氢营养型产甲烷途径能效更高,嗜氢产甲烷菌Methanoculleus丰度的增加可以有效提高甲烷的生成效率。并且Methanoculleus具有很强的耐氨性,其中铵转运蛋白和甲基铵通透酶的缺失可解释对富含铵或氨的环境的适应性特征[29]。之前的研究表明当TAN 的浓度维持在1 000 mg/L 以上时,这类产甲烷菌会成为优势微生物,本实验的TAN 处于1 000 mg/L 以上,所以这类产甲烷菌在本实验的反应器中丰度较高[30-31]。总之,生物强化使Methanoculleus相对丰度的提升可能是高温厌氧消化产甲烷效率提高的原因之一。

图6 功能微生物添加量对属水平微生物群落的影响Figure 6 Effects of bioaugmentation dosage on microorganism at genus level
3 结论
利用Ureibacillus suwonensisE11 对餐厨垃圾高温厌氧消化系统进行生物强化可以有效促进甲烷产生,其中15% 的添加剂量最佳,累积甲烷产量(以VS 计) 达到575.14 mL/g,相比 于CK 组(452.86 mL/g)提高了27.00%。功能微生物添加量为15% 时不仅有效提高了乙酸浓度,为产甲烷菌提供可利用的底物,还提高了产甲烷菌对乙酸的利用效率,使乙酸最终被完全利用。微生物群落结构演替规律表明E11 可以在厌氧消化系统内定殖,并且该菌株的添加不仅可以提高重要水解细菌的相对丰度,促进消化系统中的水解步骤,还提高了嗜氢产甲烷菌Methanoculleus的相对丰度,从而提高甲烷产率。
