山东沿海红纤维虾形草根际微生物群落结构及功能
2024-01-13张燕英王天雨沈萍萍
张燕英,王天雨,孙 浩,沈萍萍
(烟台大学海洋学院,山东 烟台 264005)
海草床是海洋生物重要的栖息地,然而,由于气候变化和人类活动的影响,全球海草床生态系统严重退化[1],海草床生态系统的保护和生态修复已成为海洋生态领域的研究热点[2]。海草床生态系统拥有多样性丰富、代谢活跃的微生物群落,微生物参与元素生物地化循环与海草宿主建立了复杂且动态变化的互作机制,进而影响海草的营养转化和健康生长[3-4]。研究显示,微生物能够消耗海草生长发育产生的有害代谢产物,如甲醇、乙醇和乙醛等化合物[5],从而有效降低这些有害代谢产物对其生长的影响,如:海草叶片上的溶藻细菌能够产生琼脂水解酶和角叉菜胶酶,杀死附生在海草叶片上影响海草光合作用的真核藻类[6]。世界范围内海草的几次大面积死亡现象均与海草床沉积物中硫化物含量过高有关[7-8],而海草根际的硫氧化微生物能够将硫化氢(H2S)等低价硫化物氧化为硫,再将硫氧化为硫酸盐,从而有效缓解海草生长的环境压力[9]。海草根际的固氮微生物能够将空气中的氮气转化为铵盐,供海草吸收利用[10];此外,海草根际微生物还能够通过产生植物激素调节海草的生长发育[5]。可见,微生物与海草组成共生功能体,在海草床生态系统中发挥重要作用[11]。
目前,海草微生物研究主要集中在鳗草、大洋波喜荡草和泰来草等海草种类[12-14],不同种类海草的微生物类群存在一定差异[15]。分布在黄渤海的红纤维虾形草(P.iwatensis)被世界自然保护联盟(IUCN)评估为易危海草种类,其微生物群落结构还未见报道。本研究采用扩增子测序和宏基因组测序技术,探究黄渤海地区红纤维虾形草根际微生物群落多样性、结构特征及功能多样性,掌握海草根际细菌优势类群、关键类群并分析其功能,推测其对海草生长、繁殖的影响,为筛选海草根际有益菌提供参考,为易危海草物种的生态修复提供理论依据。
1 材料与方法
1.1 采样位点与样品采集
根据红纤维虾形草在黄渤海的分布情况,于 2021 年 5 月份生长季节(低潮时),在荣成俚岛(37°14′39"N,122°35′8"E)和蓬莱长岛(37°55′17"N,120°45′33"E)设两个采样位点进行样品采集。俚岛采样位点位于荣成市俚岛镇,多年平均气温11.3 ℃,年平均日照时数2 578.5h,底质以砾砂为主,平均粒径1 mm,中值粒径0.56 mm,红纤维虾形草分布在海参养殖池塘附近[16]。长岛采样位点位于烟台市蓬莱区长岛海洋生态文明综合试验区,历年平均气温12.1 ℃,年平均降水量为537.1 mm,底质以黏土质粉砂为主,中值粒径介于0.005~0.089 mm之间[17]。
在每个位点设置四个平行采样点,点间距离>50 m,为不同群体样本(图1),使用无菌铲于每个采样点采集沉积物(距根表2 mm内),混合均匀后装于密封袋内,冷冻避光保存带回实验室,保存于-80 ℃冰箱。
图1 样品采集位点
1.2 DNA提取和PCR扩增
取1 g沉积物样品,根据E.Z.N.A.土壤DNA试剂盒 (Omega Bio-tek,Norcross,GA,USA)说明书进行微生物群落总DNA抽提,使用1%的琼脂糖凝胶电泳检测DNA的提取质量,使用NanoDrop2000测定DNA浓度和纯度。
分别使用16S rDNA正向引物338F(5'-ACTCCTACGGGAGGCAGCAG-3')和反向引物806R(5'-GGACTACHVGGGTWTCTAAT-3'),原生动物18S rDNA正向引物TAReukFWD1F(5'-CCAGCASCYGCGGTAATTCC-3')和反向引物TAReukREV3R(5'-ACTTTCG TTCTTGATYRA-3'),真菌ITS rDNA正向引物ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和反向引物ITS2R(5'-GCTGCGTTCTTCATCGATGC-3')进行PCR扩增,PCR反应体系为:5×TransStart FastPfu缓冲液4 μL,2.5 mmol/L dNTPs 2 μL,上下游引物(5 μmol/L)各0.8 μL,TransStart FastPfu DNA聚合酶0.4 μL,模板DNA 10 ng,dH2O补足至20 μL。每个样本3个重复。PCR反应程序为:95 ℃ 预变性 3 min,27个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),然后72 ℃稳定延伸10 min,最后在4 ℃进行保存。
1.3 Illumina Miseq 测序及数据分析
纯化后的PCR产物,利用NEXTflex Rapid DNA-Seq Kit(Bioo Scientific,USA)进行建库,并通过Illumina Miseq PE300平台(上海美吉生物医药科技有限公司)完成微生物16S rDNA、18S rDNA和ITS rDNA测序。对原始数据使用Fastq软件进行质控,并使用FLASH软件进行拼接。使用QIIME2中DADA2处理序列,获得ASV特征表。其中对于18S rDNA结果,去除Rhodophyta红藻门、Streptophyta链型植物、fungus、后生动物等获得最终的原生生物ASV分类表。
所有的数据分析均在美吉生物云平台(https:∥cloud.majorbio.com)上进行,使用Mothur软件(http:∥www.mothur.org/wiki/Calculators)计算Alpha多样性,采用Wilxocon秩和检验进行Alpha多样性的组间差异分析;使用基于Bray-Curtis距离算法的PCoA分析(主坐标分析)检验样本间微生物群落结构的相似性,并结合Adonis非参数检验分析样本组间微生物群落结构差异是否显著;通过R语言对微生物之间相关性进行计算,并使用Gephi软件绘制相关性网络图。
1.4 宏基因组测序与数据分析
用于宏基因组测序分析的DNA质检后,通过Covaris M220(上海美吉生物医药科技有限公司)将DNA片段化,筛选约400 bp的片段,使用NEXTflexTM Rapid DNA-Seq(Bioo Scientific,USA)构建PE文库,使用Illumina NovaSeq(Illumina,USA)测序平台进行宏基因组测序(上海美吉生物医药科技有限公司)。
原始测序数据经Fastp v 0.20.0进行质量控制后用于后续分析。使用Diamond v0.8.35将得到的非冗余基因集的氨基酸序列与NR数据库、eggNOG数据库(v4.5.1)、KEGG数据库(v94.2)进行比对(BLASTP比对参数设置期望值e-value为1e-5),获得基因对应的KEGG功能。使用KO、Pathway、EC、Module对应的基因丰度总和计算对应功能类别的丰度。所有的数据分析均在美吉生物云平台(https://cloud.majorbio.com)上进行,利用CD-HIT软件(http://www.bioinformatics.org/cd-hit/)进行聚类,构建非冗余基因集。使用SOAPaligner软件进行基因丰度的计算,通过R语言进行物种与功能组成分析和物种与功能贡献度分析。
2 结 果
2.1 红纤维虾形草根际微生物群落多样性分析
经过扩增子高通量测序,细菌16S rDNA共获得553 307条有效序列,平均长度420 bp;原生生物18S rDNA共获得368 154条有效序列,平均长度248 bp;真菌ITS rDNA共获得520 982条有效序列,平均长度为373 bp。基于ASV丰度表计算微生物α多样性指数,包括ASV Richness、Ace、Chao和Shannon指数,对样品中微生物群落的丰富度和多样性进行分析。长岛真菌群落的ASV Richness和Chao指数显著高于俚岛,表明长岛区域真菌群落的丰富度较高。两个海域样品的细菌和原生生物的多样性指数差异无统计学意义(表1)。
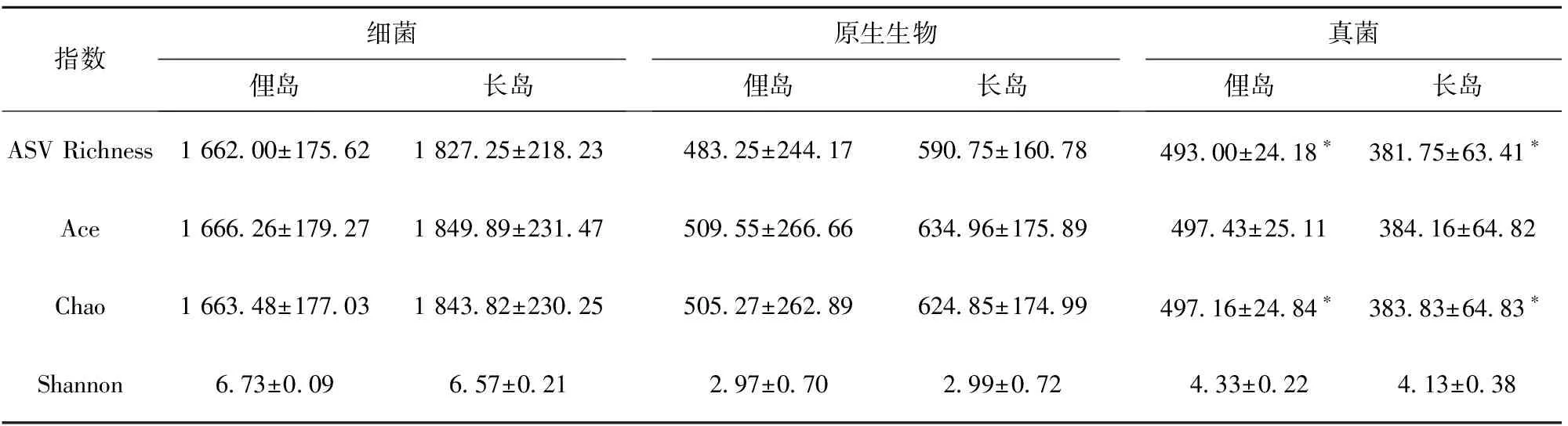
表1 红纤维虾形草根际微生物α多样性指数
主坐标分析(PCoA)结果表明不同海域红纤维虾形草根际细菌、原生生物、真菌类群都在PC1轴上显著分开(图2),表现出明显的区域特异性。多元方差分析(Adonis)结果也表明不同海域间的细菌(R2=0.46,P=0.037)、原生生物(R2=0.45,P=0.033)、真菌群落(R2=0.56,P=0.032)差异均具有统计学意义。长岛海域细菌样品间的相似性比俚岛要高,而真菌类群比俚岛要低。
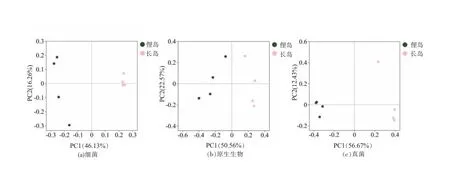
图2 基于Bray-Curtis距离的细菌、原生生物及真菌的主坐标分析(PCoA)
2.2 红纤维虾形草根际微生物群落组成分析
细菌种类在两个海域较为相似(图3(a))。在纲水平,主要类群为拟杆菌纲(Bacteroidia),分别占俚岛和长岛细菌丰度的(18.56±1.16)%和(28.42±1.41)%,其次是γ-变形菌纲(Gammaproteobacteria)、Desulfobulbia和α-变形菌纲(Alphaproteobacteria)。俚岛海域弯曲杆菌纲(Campylobacteria)和厌氧绳菌纲(Anaerolineae)相对丰度较高,分别是长岛的2.1倍和1.6倍;而长岛海域拟杆菌纲和γ-变形菌纲相对丰度较高(图3(a))。两个海域原生生物群落组成差异较大(图3(b))。俚岛原生生物优势类群为类锥体纲(Conoidasida,(2.09±1.31)%)。长岛的优势类群是石莼纲(Ulvophyceae,(26.26±14.04)%)和褐藻纲(Phaeophyceae,(6.19±5.35)%),分别是俚岛样品的32.7倍和24.3倍。此外,长岛海域甲藻纲(Dinophyceae)的丰度是俚岛的1.7倍。真菌群落结构如图3(c)所示,优势类群为粪壳菌纲(Sordariomycetes),分别占俚岛和长岛真菌的(26.69±9.17)%和(10.73±3.11)%,其次为罗兹菌门(Rozellomycota)、担子菌门(Basidiomycota)和酵母纲(Saccharomycetes)。俚岛和长岛海域真菌群落差异较大,俚岛样品中粪壳菌纲、银耳纲(Tremellomycetes)、节担菌纲(Wallemiomycetes)和伞菌纲(Agaricomycetes)的丰度分别是长岛的2.5倍、2.4倍、3.3倍和1.7倍;而长岛海域酵母纲的丰度显著高于俚岛(P=0.033)。
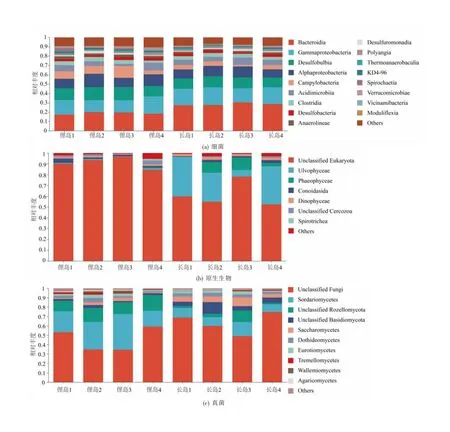
图3 纲水平上不同样品细菌、原生生物及真菌群落组成
在属水平对长岛和俚岛微生物群落进行组间差异比较,结果如图4所示,俚岛红纤维虾形草根际细菌Cocleimonas属和脱硫球茎菌属(Desulfobulbus)的丰度显著高于长岛海域(P<0.05),而伍斯菌属(Woeseia)、水杆菌属(Aquibacter)和海藻杆菌(Algibacter)属丰度显著低于长岛海域。这两海域差异显著的原生生物包括礁膜属(Monostroma)、鹅肠菜属(Endarachne)、异帽藻属(Heterocapsa)、石莼属(Ulva)和原礁膜属(Protomonostroma),这些原生生物在长岛海域的丰度均显著高于俚岛海域(P<0.05)。而真菌中帚枝霉属(Sarocladium)、梅奇酵母属(Metschnikowia)、枝孢属(Cladosporium)和克鲁维酵母属(Kluyveromyces)在两海域存在显著差异,其中长岛梅奇酵母属和枝孢属相对丰度显著高于俚岛海域,而俚岛海域帚枝霉属和克鲁维酵母属丰度显著高于长岛海域(P<0.05)(图4)。
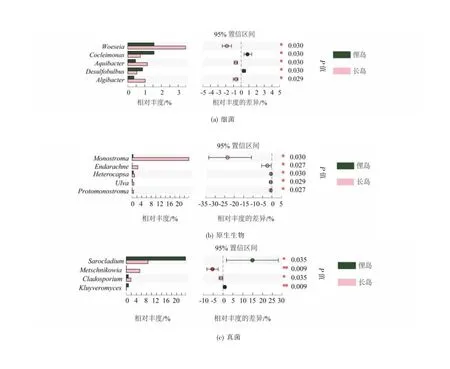
图4 不同海域样品中细菌、原生生物及真菌的差异物种分析
2.3 红纤维虾形草根际微生物生态网络分析
分别选取俚岛和长岛细菌、原生生物和真菌中总丰度前100的ASV,进行斯皮尔曼(Spearman)秩相关分析并筛选相关系数|p|>0.6,且显著水平P<0.05的数据构建相关性网络(图5),网络的拓扑参数见表2,结果显示三个微生物类群的节点数量、模块化指数、网络直径、平均路长度和平均聚类系数较为相似,而细菌的边数量较真菌和原生生物高,说明细菌之间的互作关系更加复杂(图5),原生生物间的正相关关系(俚岛和长岛分别为88.3%和77.43%)高于负相关关系(俚岛和长岛分别为11.7%和22.57%),而细菌和真菌群落不同ASV之间的正相关关系和负相关关系相差不大。
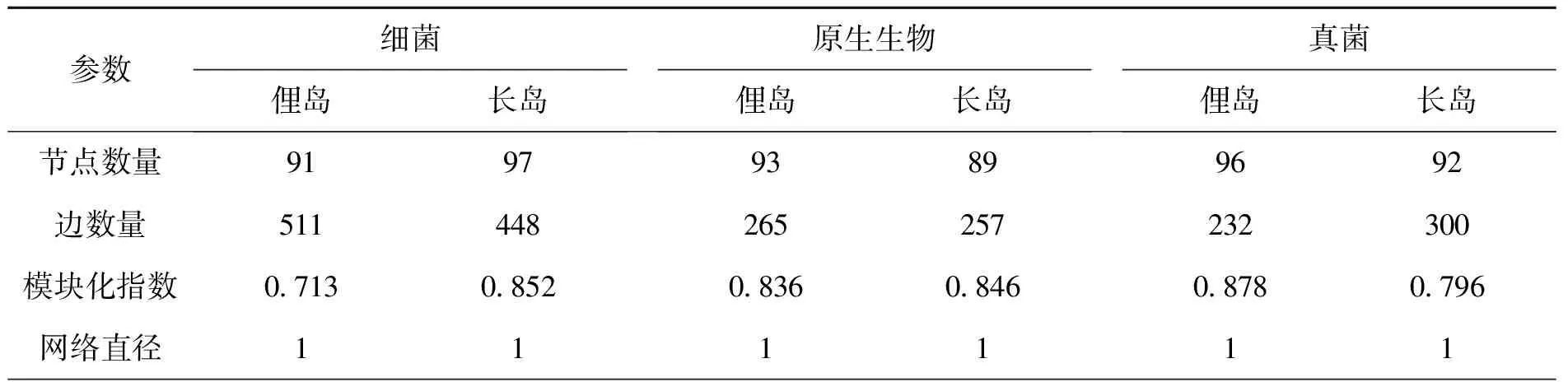
表2 微生物相关性网络图拓扑参数

图5 细菌、原生生物及真菌相关性网络
2.4 红纤维虾形草根际微生物氮代谢和硫代谢功能分析
宏基因组测序共获得原始数据862 743 138条序列,平均序列长度150 bp,经过质控,得到优化序列770 588 866条,平均序列长度150 bp,拼接组装后得到7 173 516条contig用于后续功能分析。
KEGG富集分析表明,微生物参与的相对丰度前五的功能分别是代谢途径(Metabolicpathways)、次生代谢产物的生物合成(Biosynthesis of secondary metabolites)、不同环境下的微生物代谢(Microbial metabolism in diverse environments)、碳代谢(Carbon metabolism)和氨基酸的生物合成(Biosynthesis of amino acids)(图6),这些功能涉及生物生存所必须的主要代谢功能或过程。

图6 红纤维虾形草根际微生物匹配到KEGG数据库的主要代谢通路的功能基因相对丰度
微生物物种与功能贡献度关联分析结果见图7,在属水平上共有17个属在氮代谢和硫代谢中贡献较高,其中Eudoraea只与氮代谢功能相关,主要参与反硝化作用(图7(a))。将长岛和俚岛进行组间差异分析,结果显示参与氮代谢和硫代谢的微生物中有三个属的丰度存在显著差异(图7(c)),其中长岛海域荧光杆菌属(Ilumatobacter)相对丰度显著高于俚岛海域(P<0.05),而硫卵菌属(Sulfurovum)和甲基海洋杆菌属(Methyloceanbacter)相对丰度显著低于俚岛海域(P<0.05)。荧光杆菌属在氮代谢过程中对反硝化作用贡献度较高,在硫循环过程中对硫酸盐还原贡献度较高;硫卵菌属在氮代谢过程中对硝酸盐还原贡献度较高,在硫代谢过程中对硫酸盐还原贡献度较高;甲基海洋杆菌属在氮代谢过程中对反硝化作用、硝酸盐还原和固氮作用的贡献度较高,在硫代谢过程中对硫酸盐还原贡献度较高。
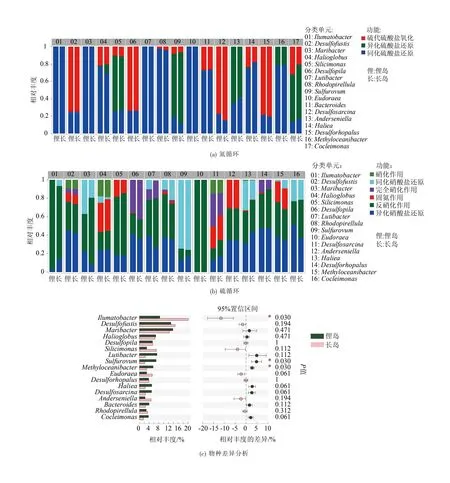
图7 硫、氮循环核心功能微生物贡献度
3 讨 论
海草根际微生物在海草床生态系统中发挥重要作用,微生物群落结构会受到海草物种、海草生长状态和环境因素的共同影响[18]。法国和葡萄牙诺氏鳗草(Zosteranoltii)和红海北部海草床的长萼喜盐草(Halophilastipulacea)根际微生物都以γ-变形菌和δ-变形菌为优势种[15],荣成天鹅湖鳗草(Zosteramarina)和日本鳗草(Zosterajaponica)根际沉积物中微生物以变形菌门和拟杆菌门为优势种[18],本研究中俚岛和长岛红纤维虾形草根际微生物均以拟杆菌门和变性菌门为优势种,说明海草根际微生物核心物种具有一致性。但是不同海域或者相同海域不同海草物种根际微生物群落结构存在差异,通常将这种差异归结为海草根系分泌物的差异和环境因子的差异造成的[15,18]。本研究中长岛和俚岛海域微生物在属水平细菌、真菌和原生生物都存在显著差异类群,进一步说明相同海草种类在不同海域根际微生物群落结构可能会受到环境影响而存在显著差异。在属水平,具有硫酸盐还原作用的脱硫叶菌属(Desulfobulbus)在俚岛海域的丰度显著高于长岛海域,脱硫叶菌属通过推动海草根际硫循环过程,对海草的健康起重要作用[7,9,19]。化能自养菌伍斯菌属(Woeseia)在沉积物氮、硫等元素循环中起重要作用,其生长依赖环境中氧气的充分供应,在长岛海域伍斯菌属的丰度显著高于俚岛,这可能与长岛海域海草根际氧气含量有关。俚岛红纤维虾形草分布在海参养殖池塘附近,海参养殖过程中的呼吸作用和生物扰动提高了沉积物耗氧量,导致俚岛海域海草根际氧气含量降低[20]。此外,微生物之间的相互作用也是造成微生物群落结构差异的原因之一,可以通过微生物相关性网络揭示微生物群落成员之间共享的生态位,或者群落成员之间更直接的共生关系[21]。使用网络分析来探索微生物分类群之间的关系,结果显示长岛区域微生物群落的复杂性高于俚岛,表明长岛区域微生物群落更稳定、细菌群落复杂性最高,可能是群落稳定性的主要贡献者。
除了细菌外,原生生物和真菌也是海草根际微生物的重要组成部分,原生生物和真菌是微食物网的重要组成部分,在能量流动和物质循环中发挥重要作用[22]。但与细菌相比,海草根际原生生物和真菌相关研究较少,本研究中原生生物和真菌丰度和多样性远低于细菌。原生生物分布广泛,其在水生食物网中占有重要地位,具有净化水质和作为水体富营养化的指标等重要作用[23]。本研究中原生生物相对丰度较高的绿藻门礁膜属(Monostroma)和褐藻门鹅肠菜属(Endarachne)均属于光养型原生生物,具有光合色素,能够利用光合作用合成有机物供应其生命活动,是海草床有机碳源的重要组成部分[24]。红纤维虾形草根际真菌优势物种是子囊菌门(Ascomycota)的粪壳菌纲(Sordariomycetes)真菌,与南海新村湾泰来草、海菖蒲和南沙群岛泰来草、喜盐草根际真菌群落存在显著差异[25]。这说明真菌在海草根际难以形成核心微生物类群,也进一步印证了真菌与海草之间的互作性不如细菌强烈[25-26]。
近年来,人类活动导致大量活性氮通过大气和河流最终排入海洋,引起水体富营养化等一系列环境问题[27]。富营养化会引发海草沉积物中有机物含量增加并抑制海草对氧气的需求,导致海草沉积物处于缺氧状态,造成硫化物的大量积累[28]。沉积物中高浓度的硫化物会严重影响海草的光合作用、养分吸收和生长代谢[29]。海草根际微生物参与海草根际硫循环和氮循环,在海草生态系统中发挥重要作用[10,30],且海草根际微生物在驱动硫循环反应过程中往往和海草根际氮循环相互耦合,如微生物的生物固氮反应通常与硫酸盐还原过程关系密切,很多海草根际的硫酸盐还原菌同时具有固氮活性[10]。本研究通过微生物物种与功能的关联分析,揭示有17个属的微生物在氮代谢过程贡献较高,其中16个属同时对硫循环代谢过程的贡献度较高,表明这些微生物同时在海草根际硫和氮循环中发挥重要作用。对两个海域的样品进行差异分析发现,长岛和俚岛海草根际硫循环和氮循环微生物有三个属存在显著差异,包括荧光杆菌属(Ilumatobacter)、硫卵菌属(Sulfurovum)和甲基海洋杆菌属(Methyloceanbacter),它们共同参与硫酸盐还原过程,在缺氧的海草根际环境中通过硫酸盐的还原进行有机质矿化过程,将硫酸盐作为呼吸链末端的电子受体,生成无机硫化物。硫酸盐还原分为同化硫酸盐还原和异化硫酸盐还原两种类型,本研究中荧光杆菌属(Ilumatobacter)和甲基海洋杆菌属(Methyloceanbacter)对同化硫酸盐还原贡献度较高,而硫卵菌属(Sulfurovum)对异化硫酸盐还原贡献度较高,同化硫酸盐还原需要消耗能量,异化硫酸盐还原是一个产能的过程[31],因此,海草根际硫酸盐还原菌的丰度差异可能与可利用能量有关。对海草根际微生物氮代谢分析显示荧光杆菌属(Ilumatobacter)和甲基海洋杆菌属(Methyloceanbacter)对反硝化贡献度较高,硫卵菌属(Sulfurovum)对同化硝酸盐还原贡献度较高。在富营养化的海草床生态系统中,微生物参与的同化硝酸盐还原和反硝化作用可以去除多余的氮,同化硝酸盐还原是利用硝态氮合成细胞物质,本研究中硫卵菌属(Sulfurovum)对同化硝酸盐还原贡献度较高,说明这类细菌在海草根际厌氧环境下能够将硝态氮转化为细胞物质,反硝化的中间产物NO是调节植物生长的信号分子[5,32],因此,在富营养化的海草床生态系统中,参与脱氮过程的微生物在海草健康生长中发挥重要作用。综上,在海草根际参与硫循环和氮循环的微生物占据相对较高丰度,说明这些微生物类群对海草生长以及维持海草根际微环境具有重要意义,也预示海草根系环境可以根据氧含量和能量控制功能微生物群落达到平衡状态,为进一步挖掘微生物资源、进行海草床生态系统的修复提供思路。
