黄馨枝干炭疽病病原菌的鉴定
2024-01-12王旭施文娟孔婷孙荣华
王旭 施文娟 孔婷 孙荣华
(1 镇江市城市绿化管理站,镇江 212000;2 上海市园林科学规划研究院,上海 200232;3 上海城市困难立地绿化工程技术研究中心,上海 200232)*为通信作者
黄馨(Jasminummesnyi),又称云南黄素馨,为常绿藤状灌木,喜温暖湿润的环境,因其适宜于堤岸、花架绿篱或坡地高地悬垂种植,在我国各地得到了广泛栽培[1]。例如,在江苏省镇江市,黄馨是高架、立交桥道路及公园绿地等区域的主栽品种。
近年来,黄馨的病害发生引起了广泛关注。目前,国内外已报道引起黄馨叶部病害的病原菌有C.cassiicola、C.siamense[2-3],但鲜有关于黄馨枝干病害病原菌的研究报道。目前,镇江市的黄馨上出现了一种新的枝干炭疽病,受害枝干在发病初期出现近椭圆形的病斑,随着病程的推进,病斑可从数厘米长发展至整个枝干,病害发生严重时易造成大量枝干枯萎死亡,给当地的生态景观效果带来较大影响。鉴于此,笔者对镇江市黄馨枝干炭疽病的病原菌进行了分离培养和致病性测定,并利用传统的形态学结合多基因联合分析构建系统发育树,进行了致病菌的鉴定,旨在为该病害的诊断及科学防控提供理论研究基础。现将相关研究结果报道如下。
1 材料与方法
1.1 症状观察
2019 年,笔者在江苏省镇江市七里甸街道,观察并记录黄馨枝干炭疽病的发病情况与危害特点,同时采集发病枝条带回实验室进行病原菌分离。
病害调查分级标准参照罗卿权等[4]提出的标准,具体为:0 级,无枝枯;1 级,1 枝无分枝的小枝枯萎;2 级,无分枝的小枝枯萎,数量大于1 枝;3 级,1枝有顶级分枝的枝条枯萎;4级,有顶级分枝的枝条枯萎且枝条枯萎数大于1 枝;5 级,1 枝或1 枝以上基部大枝(至少有两级分枝)枯萎;6级,整株枯萎。
计算公式:株发病率=(发病株数÷总株数)×100%;病情指数=[∑(各级病株数×各级代表值)÷(调查总株数×最高级代表值)]×100。
1.2 病原菌的分离及形态学鉴定
将发病症状典型的病枝用组织分离法进行病原菌分离[4],培养基为马铃薯葡萄糖琼脂培养基(Potato Dextrose Agar,PDA)。取发病枝条病健交界处约3 mm ×3 mm 大小的植物组织,经75% 乙醇消毒15~20 s、5%次氯酸钠消毒2 min、无菌水漂洗3次、晾干后,接种在含1% 乳酸的PDA 平板上,然后将其置于25 ℃恒温培养箱中,在14 h 光照/10 h 黑暗条件下进行恒温培养。培养10 d 后,在光学显微镜下观察病原菌的形态特征,记录分生孢子的形状和大小,挑取孢子,采用玻片萌发法观察附着胞的形态,参照相关文献从形态上进行初步鉴定[5-6]。
1.3 致病性测定
根据柯赫氏法则进行致病性测定。具体方法为:用无菌手术刀划伤枝条表皮,形成长度为1 cm 的伤口,将菌丝块正面朝向伤口,用已浸湿无菌水的脱脂棉保湿,每个菌株重复接种3个枝条,并设置清水对照。接种后5~7 d 观察枝条发病情况,并对发病枝条进行病原菌再分离,完成柯赫氏法则验证。
1.4 多基因联合鉴定
1.4.1 菌株DNA 提取与PCR 扩增
在接种菌株的PDA 平板于25 ℃恒温培养箱、14 h 光照/10 h 黑暗条件下培养5~7 d 后,用灭菌的刀片刮取PDA 平板表面菌丝,置于研钵中,加入液氮研磨,采用CTAB 法提取基因组DNA[7]。对真菌的核糖体内转录区间隔区(internal transcribed spaces,ITS)、肌动蛋白基因(actin,ACT)、3-磷酸甘油醛脱氢酶基因(glyceraldehydes-3-phosphate dehydrogenase gene,GAPDH)、ApMAT基因 (partial mating type Mat1-2 gene)、谷氨酰胺合成酶基因(glutamine synthetase gene,GS)、β-微管蛋白基因(β-tubulin gene,TUB2)进行扩增,引物序列及退火温度等见表1[8-9],引物设计由上海生工生物工程技术服务有限公司完成。PCR 体系总体积为25.0 μL,包括0.1 μL Taq 聚合酶(5 U/μL)、2.5 μL PCR Buffer(含Mg2+)、2.0 μL dNTPs、引物(10 μmol/L)各1.0 μL、1.0 μL 模板DNA,然后用灭菌ddH2O 补足25.0 μL。PCR 产物用1% 琼脂糖凝胶电泳检测。

表1 引物序列、退火温度及PCR 产物长度
1.4.2 聚类分析
采用NCBI 数据库中的Blastn 工具进行序列比对和同源性分析,下载相似性高的序列及其对应模式菌的序列用于聚类分析。将所需序列,用Clustal X 2.0.1 0 进行比对剪切后,按ITS-GAPDHACT-TUB-ApMAT-GS的顺序分别首尾相连接,用MEGA-X 软件以邻接法(neighbor joining,NJ)构建系统发育树,利用自展法 (Bootstrap,1 000次重复)检验各分支的置信度。
2 结果与分析
2.1 病害症状观察及病情调查
黄馨枝干炭疽病在镇江市始发于4月下旬,故本研究于2019 年5 月—9 月进行调查,共调查50 株黄馨。结果表明,5月黄馨枝干炭疽病的病株率为28%、病情指数为12.5,6 月黄馨枝干炭疽病的病株率为48%、病情指数为19.0,7 月黄馨枝干炭疽病的病株率为62%、病情指数为32.0,8月黄馨枝干炭疽病的病株率为78%、病情指数为40.3,9月黄馨枝干炭疽病的病株率为82%、病情指数为42.6。黄馨枝干炭疽病在初发时,受害枝干出现黑褐色、近椭圆形病斑,随着病情的推进,病斑中心凹陷,病斑扩展延长呈长椭圆形或不规则形,且无分枝的小枝受害时,小枝萎蔫;在发病后期,基部枝条上病斑沿枝干扩展成条块状,使枝条上叶片枯萎,直至黄馨整株萎蔫干枯。病原菌不仅可以侵染黄馨较小的分枝,导致分枝枯死,也可侵染黄馨基部主枝,导致黄馨整株枯萎死亡。
2.2 病原菌分离与致病性
从采集的发病症状典型的黄馨病枝中,分离培养得到10 个菌落形态一致的分离物(命名为JM-1),其菌落呈圆形,边缘整齐,初期为白色;气生菌丝为白色至灰白色,后渐变为深灰色、絮状或绒状,培养皿背面呈现不均匀的灰白色。挑取菌落边缘的菌丝转移纯化,将菌株命名为2019JM。为验证菌株2019JM 的致病性,将其接种于黄馨枝条,接种5 d 后发现,接种枝条萎蔫,发病症状与黄馨枝枯病自然状态下的感染症状相同,接种清水的枝条不发病。对接种发病的黄馨枝条进行病原菌再次分离,所得分离物的菌落形态与原分离物JM-1 一致,证明2019JM 为黄馨枝干炭疽病的病原菌。
2.3 病原菌形态学鉴定
将病原菌2019JM 接种在PDA 培养基上,置于25 ℃恒温培养箱中、于14 h 光照/10 h 黑暗条件下,培养5 d 后,病原菌分生孢子呈椭圆形至长椭圆形,两端钝圆或一端略尖,无隔膜,单细胞,无色;分生孢子大小基本相近,大小为(12.1~16.5)μm×(4.0~5.0)μm,见图1-A。分生孢子萌发后,在顶端产生附着胞,附着胞为黑褐色、棍棒形或椭圆形,边缘大多规则,大小为(5~11)μm×(4~8)μm,见图1-B。依据孢子和附着胞的形态特征,将其初步鉴定为炭疽菌属真菌,具体的种需要采用分子手段进行进一步鉴定[5-6]。
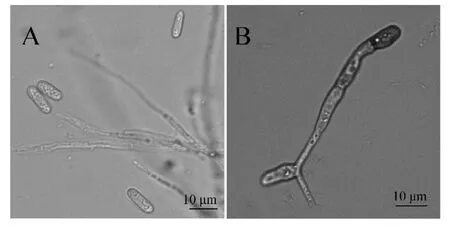
图1 菌株2019JM 的形态特征
2.4 病原菌rDNA-ITS 序列分析
提取病原菌基因组DNA 并对ITS、GAPHD、GS、TUB2、ACT和ApMAT序列片段进行PCR 扩增,将序列提交给NCBI 数据库,获得序列号,具体见表2。以Colletotrichumhippeastri(CBS 241.78)作为外群[6],多基因序列系统发育树的聚类分析结果见图2,菌株2019JM 与GenBank 中的模式菌株暹罗炭疽菌C.siamense(ICMPl8578、LC2940、LC2941、CBS125378) 以很高的支持率聚为一支,表明菌株2019JM 为暹罗炭疽菌。因此,综合依据形态学鉴定和系统发育树分析,本研究将引起江苏省镇江市黄馨枝干炭疽病的病原菌鉴定为C.siamense。
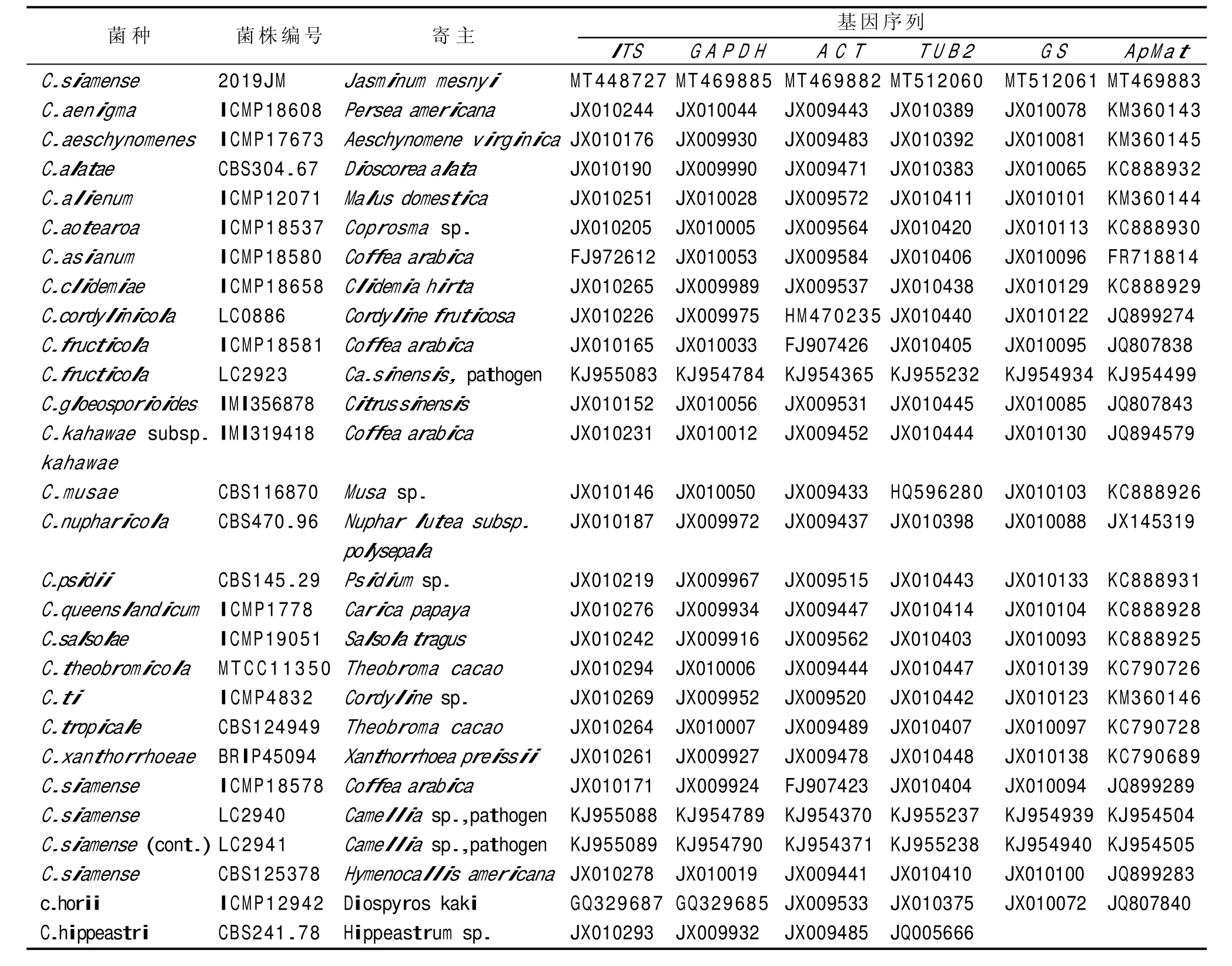
表2 系统发育分析中的菌株信息
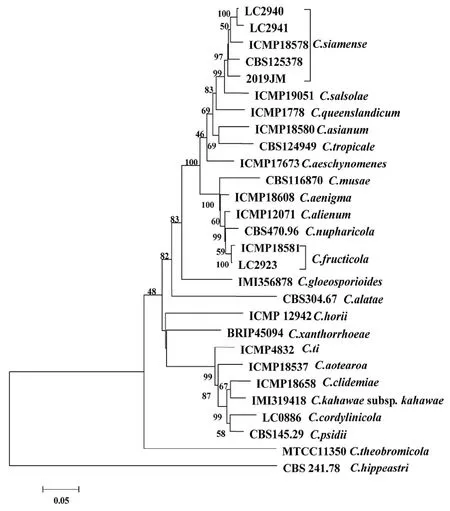
图2 基于多基因序列构建系统发育进化树
3 结论与讨论
炭疽菌属(ColletotrichumCorda)真菌寄主广泛,是世界范围内许多粮食作物及经济作物上的重要致病菌[5-6],但是,炭疽菌属不同种之间的形态特征差异微小,且继代培养性状不稳定,故依据形态特征鉴定炭疽菌不够准确。同时,ITS基因序列难以区分胶孢炭疽菌复合群下近缘相似种之间的差异[10],但依据多基因序列可以有效鉴定胶孢炭疽菌复合群下各种,故目前,多基因分析方法已成为炭疽菌属真菌鉴定的必要手段[5-6]。例如,ApMAT和GS的单基因或者双基因可以将暹罗炭疽菌从胶孢炭疽菌复合群下鉴定出来[5-6]。本研究前期采用单基因分析发现,依据ITS基因发育树的自展支持率较低,不能有效区分各序列的进化关系,而GS、TUB-2、ApMAT基因可以将暹罗炭疽菌的各菌株聚在一起,但是对胶孢炭疽菌复合群下其他种的自展支持率较低,说明只有依赖多基因序列才能有效区分胶孢炭疽菌复合群下各种的进化关系。
暹罗炭疽菌C.siamense最早在泰国咖啡上被发现[10],随后有C.siamense在草莓、葡萄、油茶、辣椒、橡胶等多种寄主植物上发生的报道[6,8,11-14],由于C.siamense具有较强的致病力,故对寄主植物的经济价值和生态效益构成了严重威胁。在镇江市,C.siamense为害黄馨枝干引起的炭疽病一般在4 月下旬开始发生,病害初发时,受害枝条上出现不规则病斑,无分枝的小枝受害时,会造成小枝枯萎;在5 月—6 月,黄馨枝干炭疽病的病株率有所升高,但病害总体发生程度较轻,偶有顶级枝条萎蔫,多表现为无分枝的小枝枯萎和枝干部位出现病斑;7 月—9月是黄馨枝干炭疽病的发病高峰期,且此时夏季高温,受温度胁迫易造成大量枝干枯萎死亡,甚至造成黄馨整株枯死,严重影响了黄馨的生态景观价值。
本研究利用病原菌形态特征结合多基因联合构建系统发育树分析,首次明确了引起镇江市黄馨枝干炭疽病的病原菌为C.siamense,这为该病害的诊断提供了理论依据,且这是国内首次报道C.siamense引起黄馨枝干炭疽病。
有研究认为,炭疽菌易发生复合侵染,即多种炭疽菌可同时侵染同一寄主植物。另据文献报道,辣椒炭疽病的致病菌有4 种;李杨等[14]在海南省的油茶上分离到3种不同的炭疽菌。因此,关于黄馨枝干炭疽病是否存在优势菌株和复合侵染现象也有待进一步研究确认。
