线粒体调控在阿尔茨海默病铁死亡中的作用研究进展*
2024-01-09孟炜寒
孟炜寒, 杜 悦, 朱 乐, 曾 燕, 魏 珍,2△
武汉科技大学 1脑科学先进技术研究院 2职业危害识别与控制湖北省重点实验室,武汉 430065
阿尔茨海默病(Alzheimer’s disease,AD)是一种以渐进性认知功能丧失和行为损害为主要特征的中枢神经系统退行性疾病,是最为常见的痴呆类型,约占70%[1]。目前,全球约有5000万AD患者,到2050年将达到1.52亿[2]。高龄是AD的主要危险因素,然而AD患病已逐渐年轻化[3],有病例报道最年轻的AD确诊患者为19岁[4]。阿尔茨海默病的主要临床症状表现为记忆困难,学习障碍以及精神疾病样行为(如抑郁、易怒、妄想等),病情严重者可能出现瘫痪,生活不能自理[5],给患者家庭以及社会带来巨大的精神压力和经济负担。阿尔茨海默病的发生发展极为复杂,涉及APP蛋白的异常剪切、Tau蛋白的过度磷酸化修饰、ApoE4基因多态性、胶质细胞促发的免疫炎症、微生物感染、睡眠、年龄以及社会环境因素(比如受教育程度)等,至今未能清晰解释AD的发生机制,当然也无有效治疗药物[6]。
随着研究的深入,除了经典的淀粉样蛋白级联假说,近年来一些新的AD致病假说被提出,例如线粒体级联假说[7],金属离子紊乱假说[8],脑肠轴假说[9]等。线粒体结构和功能变化可能造成神经元铁死亡(ferroptosis)是AD致病机制中的重要一环,本文主要综述了细胞中铁代谢以及线粒体内脂代谢、氨基酸代谢异常在AD神经元铁死亡中的作用及机制。
1 铁死亡的基本特征
铁死亡(ferroptosis)是一种具有铁依赖性的新定义的程序性细胞死亡,且具有脂质依赖性与铁依赖性[10]。发生铁死亡的细胞往往出现细胞膜起泡但不破裂,线粒体体积减少、双层膜密度增加、嵴减少或消失等形态学特征。细胞内铁离子依赖的活性氧(reactive oxygen species,ROS)过量积累,谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)的抗氧化作用减弱,使ROS生成与清除之间的稳态被打破,从而促进细胞膜磷脂过氧化引发细胞发生铁死亡,而这一过程受到许多基因调控[11]。最近的研究表明,铁死亡与许多疾病的病理生理过程密切相关,如肿瘤、神经系统疾病、缺血再灌注损伤、肾损伤和血液疾病[12]等。
1.1 铁死亡的细胞形态
铁死亡细胞在显微镜下呈空的胞质组成的透明圆气球形[13],细胞膜起泡而不破裂,细胞核大小正常。铁死亡细胞不具有细胞凋亡的典型特征,不会出现凋亡小体、细胞萎缩和染色质凝集等现象[14],也不会形成自噬小体。它也不具有经典的坏死形态,如细胞器与细胞质的肿胀[11]。而在亚显微结构上,铁死亡细胞主要表现出线粒体形态变化,如体积缩小,双层膜密度增加,线粒体外膜(outer mitochondrial membrane,OMM)断裂,线粒体脊的正常结构被破坏等[14]。这些形态学特征有助于区分铁死亡与其他细胞死亡模式,如细胞凋亡、坏死、焦亡和自噬。
1.2 铁死亡的生化与基因调控过程
铁死亡关键的生化过程即膜脂质过氧化。促进该过程的分子或事件会诱导铁死亡发生,而抑制或缓解该过程的则阻碍其发生。由于细胞内氧化还原活性二价铁(Fe2+)数量的增加[15],过量的二价金属阳离子易发生芬顿反应(Fenton reaction),产生过多氧自由基,使细胞膜上的多不饱和醚磷脂(polyunsaturated ether phospholipids,PUFA-ePLs)过氧化。这一过程受到GPX4调控,GPX4将过氧化脂质(LOOH)转化为相应的脂质醇(CHO)[16],从而缓解过氧化脂质的积累。GPX4同时消耗细胞内抗氧化储备还原型谷胱甘肽(glutathione,GSH),将其转换成氧化型谷胱甘肽(glutathiol,GSSG),因此,细胞内GSH的可利用率影响GPX4的活性,进而影响过氧化脂质的还原。GSH由半胱氨酸(cysteine,Cys)和谷氨酸(glutamate,Glu)合成,细胞内这两种氨基酸的浓度受氨基酸逆向转运系统Xc-(System Xc-)精细调控。系统Xc-是由4F2重亚基和xCT轻亚基组成的异源二聚体,分别由溶质载体-家族7成员11(solute carrier family 7 member 11,SLC7A11)和溶质载体家族3成员2(solute carrier family 3 member 2,SLC3A2)基因编码。线粒体是细胞内ATP的主要产地,也是ROS的主要来源。铁死亡抑制蛋白1(ferroptosis suppressor protein 1,FSP1)因子将辅酶Q10(CoQ10)还原为泛醇,从而限制ROS产生和脂质过氧化[17]。
在遗传学上,细胞铁死亡是由铁代谢相关基因如转铁蛋白及受体(transferrin/transferrin receptor 1,Tf/TfR1)、铁蛋白重链(ferritin heavy chain,FHC)、铁转运蛋白1(ferroportin 1,FPN1),脂质合成和修饰基因如脂质合成酰基辅酶a合成酶家族成员2(acyl-CoA synthetase long chain family member2,ACSF2)、酰基辅酶a合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4),以及氧化应激基因如ATP合酶F0复合体C3亚基(ATP synthase F0 complex subunit C3,ATP5G3)、柠檬酸合成酶(citrate synthase,CS)共同驱动的[18]。有研究发现,铁调节蛋白2(iron-regulatory protein 2,IRP2)转录后水平调节铁代谢基因表达,敲低IRP2将沉默铁代谢基因的表达。IRPs能结合目标mRNA上的铁反应元件(iron response element,IRE)从而调节细胞内铁稳态[19]。当细胞缺铁时,IRPs与IRE结合的亲和力增高,抑制铁蛋白mRNA的翻译,同时稳定TfR1 mRNA。当细胞铁充足时,IRPs与IRE结合的亲和力降低,增加铁蛋白合成并促进TfR1 mRNA降解[20]。
1.3 铁死亡的局限性
近年来,铁死亡的作用在越来越多的疾病中被发现,例如癌症、炎症、缺血再灌注损伤疾病以及神经系统性疾病等。但在一些疾病的相关研究中,铁死亡的作用仍具有局限性。在癌症中,Erastin等药物诱导的铁死亡可抑制肿瘤细胞的生长,然而,铁死亡损伤可以在肿瘤微环境中引发炎症相关的免疫抑制,从而有利于肿瘤生长。铁死亡对肿瘤生物学的影响程度尚不清楚,但是一系列研究发现癌基因(例如RAS和TP53)的突变、应激反应途径(如NFE2 L2信号传导,自噬和缺氧)、上皮间充质转化以及激活铁死亡的治疗反应之间存在重要相关性[21]。在神经系统疾病中,铁死亡在AD、帕金森症(Parkinson’s disease,PD)以及亨廷顿病(Huntington’s disease,HD)、肌萎缩侧索硬化症(amyotrophic lateral sclerosis,ALS)中都被发现可能发挥重要作用[22]。研究表明,在AD、PD和HD临床病理中已发现脑铁积累和脂质过氧化[23];而铁死亡抑制剂被发现在HD细胞模型以及PD的MPTP小鼠模型中具有保护作用[24];在PD和ALS患者的Ⅰ期临床试验中使用抗铁死亡化合物也有良好的疗效[25]。关于铁死亡与神经退行性疾病的潜在关系得到了越来越多的发掘,但是关于铁死亡在神经退行性疾病中的具体作用机制尚未得到准确的验证。
2 AD细胞中铁稳态失衡且线粒体功能障碍
铁的代谢是铁死亡过程中一个重要的调节点,其中许多代谢酶和代谢物已被作为铁死亡调节剂的靶点。“游离”铁与过氧化氢易诱发芬顿反应[26],产生大量羟基自由基,发生氧化作用,从而促进细胞内脂质、蛋白质和核酸的过氧化尤其是脂质过氧化并引起细胞铁死亡。因此,铁稳态的维持对抵抗细胞死亡十分必要。维持细胞内铁稳态是一个复杂的过程,依赖于多种铁代谢相关蛋白的协调。研究发现多种铁代谢相关蛋白在AD患者和小鼠模型的大脑中异常表达,导致铁沉积并促进AD进展[27]。Tf及其细胞表面受体TfR1主要负责细胞内铁的吸收。Trf1循环异常可导致神经退行性病变并伴有脑铁沉积的神经性病变(neurodegeneration with brain iron accumulation,NBIA)[28]。细胞内铁的转出依赖于FPN,而FPN是目前已知唯一的铁外输转运体。FPN表达减少时会使细胞内二价铁离子蓄积,引发铁死亡以及氧化应激,神经元功能受损[29]。在神经元中,淀粉样蛋白前体蛋白(amyloid precursor protein,APP)连接到FPN并稳定FPN的表达。APP也是一种铁氧化酶,可以氧化Fe2+为Fe3+并将其移出细胞。二价金属转运蛋白1(divalent metal transporter 1,DMT1)是另一种重要的铁吸收蛋白,其过度表达会积累细胞内不稳定的亚铁达到铁过载的状态,导致黑质中铁的积累和多巴胺能神经元的丢失[30]。铁蛋白是细胞内主要的铁储存蛋白,由FTL1(轻铁蛋白)和FTH1(重铁蛋白)组成。铁蛋白主要调节还原状态(Fe2+)的活性铁和氧化状态(Fe3+)的非活性铁之间的平衡以保存多余的铁,防止细胞和组织受到氧化损伤[31]。当细胞内铁蛋白表达异常,游离铁含量增多时,会导致细胞内铁稳态失衡引发铁死亡。核受体共激活因子4(nuclear receptor coactivator 4,NCOA4)是一种选择性受体,负责与铁蛋白结合并将其运输到溶酶体进行降解并释放活性铁(Fe2+),此过程被称为铁蛋白吞噬(ferritin phagocytosis)[32]。铁死亡可以通过铁蛋白吞噬促进铁和ROS积累来触发。当NOCOA4表达下调时,会导致铁蛋白吞噬现象减少,细胞内游离铁增加,加速细胞内铁死亡现象发生(图1)。一些证据表明,铁蛋白吞噬介导的铁死亡是导致神经退行性疾病的关键机制之一,破坏铁蛋白吞噬功能会导致铁代谢缺陷,可能导致过度氧化应激,导致心脑血管疾病、肿瘤和神经退行性疾病等疾病的发生[33]。在PD相关视网膜变性中,α-突触核蛋白(α-synuclein)会破坏铁蛋白吞噬,导致铁蛋白积聚和视网膜铁过载[34]。NCOA4也参与了维持神经元内线粒体的稳态平衡和线粒体质量控制,是神经元线粒体健康的重要调节因子之一[35]。例如,右美托咪定(Dexmedetomidine,DEX)是一类具有神经保护作用的药物,可以通过NCOA4介导的铁蛋白自噬来逆转甲氨蝶呤(Methotrexate,MTX)诱导的海马HT4细胞系的神经毒性和炎症[34]。
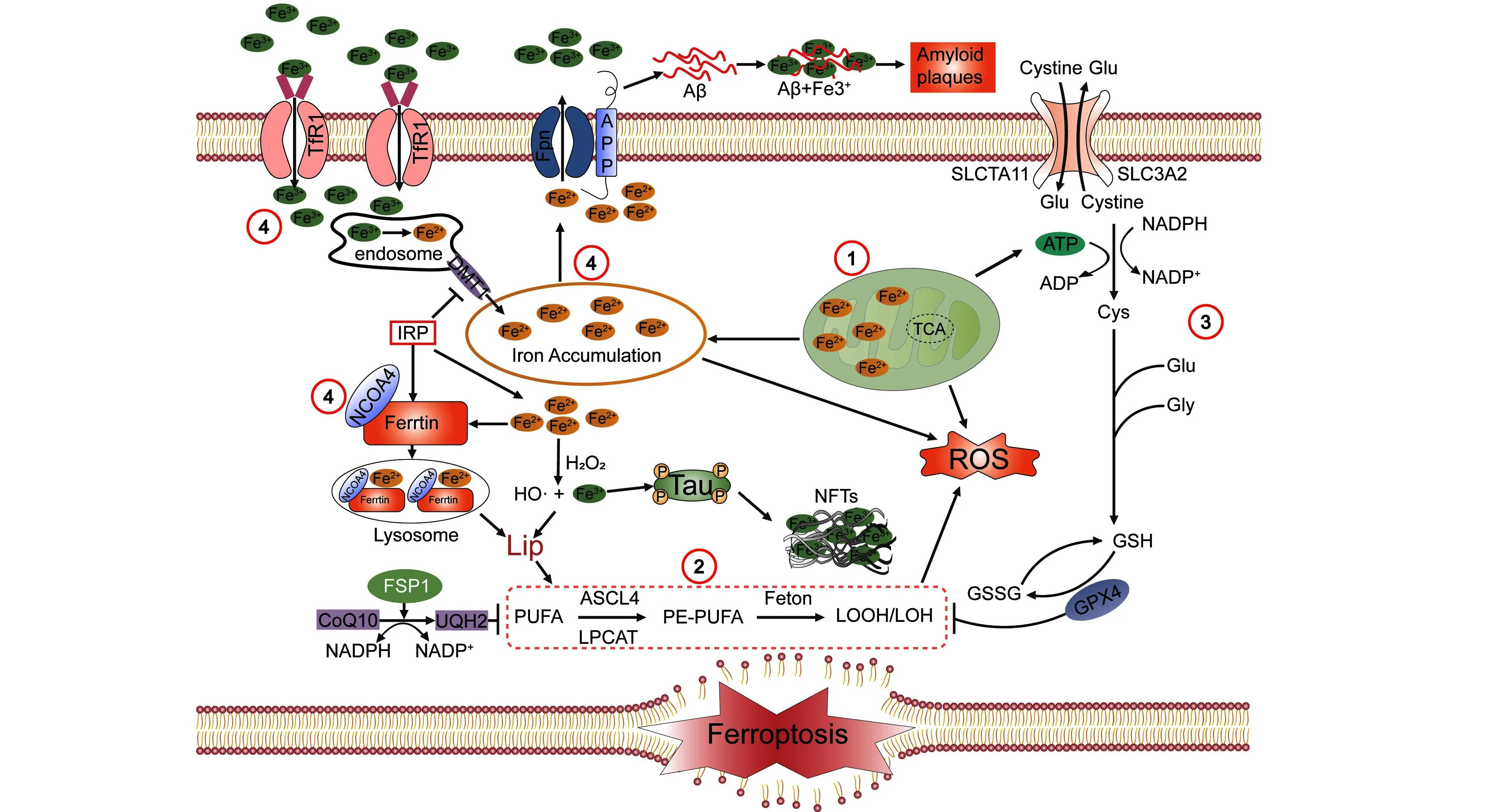
在阿尔茨海默病中铁死亡调控机制主要包括:①线粒体结构破坏或功能障碍会导致ATP产量降低、ROS升高;②脂质氧化的代谢酶活性或表达增加,脂质过氧化增强:③半胱氨酸、谷氨酸和甘氨酸合成GSH受阻,或者GSH利用受阻,抗氧化能力弱;④铁离子输入转运体增加,输出转运体减少,导致铁累积
早在上世纪60年代就有研究者指出有证据表明大脑皮层铁的积累与AD的发展之间存在关联。研究者在AD患者脑中中观察到游离铁含量升高以及铁蛋白水平升高,且铁超负荷与AD的认知能力下降成正比[36]。淀粉样蛋白β(Aβ)和过度磷酸化tau是AD的两个病理标志,也可以促进铁在大脑中的沉积,形成AD发展的恶性循环[37]。AD中存在的APP功能受损可引发细胞内Fe2+和Cu2+水平升高,并减少细胞外Cu2+,从而促进其积累。细胞内Fe2+和Cu2+过量累积会导致ROS增加引起氧化应激。此外,铁、铜和锌对Aβ具有较高的结合亲和力,可以促进其聚集。增加的神经元铁、铜和锌也与Tau蛋白结合并促进NFT的形成。因此,在斑块和神经纤维缠结(neurofibrillary tangle,NFT)中发现了过量的重金属[38]。由此可见铁稳态失衡在AD中已被得到强力的证实。
铁稳态失衡不仅导致铁死亡,也是导致线粒体功能障碍的重要因素。通常线粒体功能障碍引起的疾病(如肿瘤、慢性炎症、神经退行性疾病等)都是由线粒体内自由基产生过多导致的,而铁稳态失衡则可能导致自由基水平升高,使线粒体受到更多的损伤[39]。例如,铁稳态失衡会导致ROS增加,ROS可分解线粒体膜上的酶并破坏线粒体膜,导致线粒体膜电位下降[40]。同时,线粒体内的铁离子还可能与硫基团结合形成自由基,进一步强化氧化应激反应,导致线粒体膜及膜上酶的结构和功能受到破坏,从而引发细胞死亡[41]。因此铁稳态失衡可能通过增加ROS而加重线粒体功能障碍。线粒体和钙离子的紧密联系使其能够快速的负责很多代谢功能,钙离子浓度能够影响线粒体内酶的活性,影响线粒体功能,这个过程称为线粒体级联反应(mitochondrial cascade hypothesis)[42]。然而,级联反应可能会导致神经系统的脆弱性并引起一系列疾病,如阿尔茨海默病[43]。铁离子的稳态失衡也可能干扰细胞内钙离子浓度,导致钙和线粒体之间的相互作用受到影响,从而出现线粒体失控和细胞功能障碍[44]。
总的来说,铁稳态失衡可能通过多重途径引起线粒体功能障碍从而调控包括AD在内的多种神经性疾病的发生发展。
3 脂质过氧化与线粒体障碍相互作用促进AD的发生发展
氧化损伤是由于ROS的过度产生和抗氧化剂中和或消除其有害影响的能力失衡造成的。ROS介导的脂质过氧化是导致铁死亡的关键步骤[45]。脂质过氧化链式反应的产物具有很高的生物活性破坏力,它能损害DNA、蛋白质和酶的活性,并作为激活信号通路的启动因子,启动细胞死亡[46]。而膜磷脂(phospholipids,PLs)的过氧化是引发铁死亡的直接事件[47],尤其是多不饱和脂肪酸(polyunsaturated fatty acids,PUFAs)的过氧化[48]。脂质组学研究表明,磷脂酰乙醇胺类膜磷脂上的花生四烯酸和腺苷脱氨酶是铁死亡过程中发生过氧化的主要PUFAs。PUFAs通过酰基辅酶A合成酶长链家族成员-4(acyl-CoA synthetase long-chain family member 4,ACSL4)酯化为PUFA-CoA,然后被溶血磷脂酰胆碱酰基转移酶-3(lysophosphatidylcholine acyltransferase3,LPCAT3)修饰,以便整合到细胞膜中才能被氧化并成为铁死亡信号[49]。因此,ACSL4和LPCAT3是调控铁死亡的关键酶,如果ACSL4和LPCAT3缺失,铁死亡将会被抑制[50]。此外,脂质过氧化的主要产物4-羟基壬烯醛(4-hydroxynonenal,4-HNE)可诱导ROS的产生,从而进一步反馈激活脂质过氧化反应和铁死亡发生,因此4-HNE的增加也被认为是铁死亡的判断指标[51]。线粒体通过脂肪酸代谢和谷氨酰胺分解为铁死亡提供所需的特定脂质前体。此外,在线粒体外产生的脂质过氧化物可通过氧化应激的传播引起线粒体内脂质过氧化和线粒体损伤,从而破坏线粒体对铁稳态的调节,最终导致铁死亡的发生[52]。
许多神经退行性疾病(AD、PD、HD等)的发展已被充分证明与发生过脂质过氧化有关。在AD小鼠和AD患者受损的星形胶质细胞中检测到脂质过氧化产物4-HNE水平显著升高[53]。研究表明,Aβ肽可激活NADPH氧化酶(NADPH oxidase,NOX)引起脂质过氧化,而脂质过氧化产物又促进了APP的加工,从而加剧了Aβ肽的积累[54]。而且AD患者大脑中也存在脂质过氧化产物和Aβ斑块的位置共存现象并且铁离子增多导致的ROS升高会导致大量低聚Tau的形成。最新研究指出,肠道微生物群可以通过PUFAs相关的神经炎症调节阿尔茨海默病病理和认知障碍[55]。研究人员发现,AD患者以及模型小鼠的肠道微生物群明显增多,其中拟杆菌能增强宿主大脑中促炎性PUFA的代谢并激活小胶质细胞和AD病变[55]。脂氧合酶(lipoxygenase,LOX)[56]、环氧合酶(cyclooxygenase,COX)[57]、细胞色素c(cytochrome c)[58]是参与脂质过氧化的重要代谢酶,它们均被发现在AD细胞中异常表达或活性增强。例如,AD患者大脑中12/15-脂氧合酶(12/15-LOX)的酶活性显著增加[59],高活性的12/15-LOX降低了AD模型小鼠的记忆和学习能力。基于铁死亡通路中的脂质过氧化在AD病理进展中的重要作用,一些AD治疗药物的研发靶向此过程。最新研究表明,中药材红景天可通过下调TFR1和ACSL4蛋白表达,抑制铁死亡的发生,从而减少Aβ斑块的积累并挽救神经元损伤来减缓认知障碍[60]。
脂质过氧化与线粒体功能障碍紧密联系且相互作用于神经系统疾病。线粒体电子传递链上的氧化磷酸化过程是大多数哺乳动物细胞内ROS的重要来源。线粒体移植实验发现含有阿尔茨海默病患者线粒体DNA的细胞系表现出更多的活性氧和自由基的产生[61]。局部过量ROS的产生不仅会导致线粒体损伤,还会促进铁死亡[62]。细胞内抗氧化机制依赖于线粒体内GSSG与GSH的相互转换,谷胱甘肽过氧化物酶4(GPX4)作为铁死亡核心调控因子能催化GSH转换成GSSG,同时伴随毒性脂质过氧化物(LOOH)转换成脂质醇(CHO)从而抵抗细胞膜上脂质过氧化反应[16](图1)。值得注意的是,定位于线粒体内膜的二氢乳清酸脱氢酶(dihydroorotate dehydrogenase,DHODH)可以通过生成一种自由基捕获型抗氧化剂(radical trapping antioxidant)CoQH2来阻止脂质过氧化,从而抑制铁死亡,此过程是GPX4非依赖性的,因此若同时靶向GPX4和DHODH将会提高相关疾病的治疗效果[63]。最近多项研究表明转位于线粒体的缺氧诱导因子(transcription factor hypoxia inducible factor,HIF)也参与线粒体脂质代谢从而调控细胞铁死亡[64]。AD中磷酸化的Tau蛋白已被证明可以阻止线粒体运输,导致突触的能量剥夺和氧化应激,进而导致神经退行性变[65]。减少氧化应激、脂质过氧化和铁过载,可进一步抑制铁死亡,从而削弱对海马神经元和突触可塑性的损伤,最终恢复认知功能[66]。而且AD病理中发现许多铁死亡证据,包括铁积累过多,脂质过氧化物和ROS水平升高,GSH和GPX4水平降低等。
总而言之,脂质过氧化作为铁死亡与AD病理的共同特征,线粒体在两者之间的桥梁作用不可忽视,但关于三者之间更加准确的调控机理仍有待研究,未来厘清其中的分子机制有助于探索AD预防和治疗的新靶点。
4 线粒体内氨基酸代谢异常诱发AD铁死亡发生
系统Xc-是磷脂双分子层中广泛存在的氨基酸逆向转运体,是由SLC7A11和SLC3A2两个亚基组成的异二聚体[67]。系统Xc-分别以1∶1的比例控制细胞内胱氨酸(cystine)和谷氨酸的进出[68]。胱氨酸在细胞中被还原为半胱氨酸(Cysteine,Cys),后者与谷氨酸、甘氨酸形成三肽的GSH(图1)。抑制Xc-系统活性可以阻碍胱氨酸的吸收从而减少GSH的合成,导致GPX4活性降低和脂氧合酶活化,细胞抗氧化能力下降,脂质活性氧积累,最终发生氧化损伤和铁死亡[69]。也有报道指出,在NFT的形成过程中,铁的累积可以通过ROS累积,介导氨基酸作用促进Tau的形成。铁可以通过形成通过磷酸化氨基酸残基介导的分子间配位复合物来产生低聚Tau,通过Cys-Cys的结合介导Tau的磷酸化[70]。
线粒体内的氨基酸代谢,也可以产生NADPH等还原物为呼吸链氧化磷酸化提供底物以补充ATP生成,为细胞提供能量。研究表明,线粒体功能障碍与氨基酸代谢异常存在密切联系[71]。例如,精氨酸代谢异常可损害线粒体稳态[72],N-乙酰半胱氨酸(N-acetylcysteine,NAC)可以拯救胎盘中的线粒体损伤以及铁死亡[73]。在线粒体内,半胱氨酸剥夺(cysteine deprivation,CD)诱导的铁死亡需要谷氨酰胺分解(将谷氨酰胺转化为谷氨酸),通过α-酮戊二酸脱氢酶(alpha-ketoglutarate dehydrogenase,KGDH)复合物促进线粒体内三羧酸循环。KGDH不仅参与谷氨酸水平的调节,还充当氧化还原传感器以维持线粒体氧化还原状态[74]。有研究发现,线粒体活性增加会降低γ氨基丁酸的信号传导会导致社交行为障碍与神经元稳态被打破[75]。GSH作为细胞内的主要抗氧化剂,在线粒体铁硫(iron-sulfur,FeS)簇生物合成中是必不可少的,通过共价连接FeS簇作为CGFS型(Ⅱ类)戊二醛(glutaredoxins,Grxs)的辅助因子,以帮助FeS前体从线粒体输出[76]。
越来越多的证据表明GSH的失调可能与神经变性相关联。AD患者大脑中GSH水平降低[77],血清中Cys的水平显著降低[78]。低水平的Cys以及GSH会促进AD细胞铁死亡。而GSH作为抗氧化剂,共同作用于铁死亡以及AD的病理过程也有大量研究。研究发现,当细胞内GSH降低时,会导致Fe2+转运受损,进而引发铁死亡。同时Fe2+转运受损会导致线粒体功能受损从而可能导致AD的发生[79]。同时低水平的GSH也会造成细胞内其它金属离子紊乱(例如铜)从而增加神经细胞的死亡[13]。
5 总结与展望
随着全球AD发病率的逐年攀升,寻找新的有效的药物靶点迫在眉睫。本文从线粒体调控铁死亡的病理机制入手,从铁稳态失调、脂质过氧化以及氨基酸代谢紊乱三个方面展开讨论,综述了线粒体功能障碍调控铁死亡在AD发病中作用的研究进展,以期为AD寻找更有效的治疗方案提供新思路。针对AD铁死亡的药物研发目前还处于基础研究阶段,而一些铁螯合剂已经用于临床缓解铁沉积,消除脑内自由基。随着对AD病理机制的认知加深,未来有望针对多靶点共同调控网络,并结合中药复方疗法,采用联合治疗方案,为根治AD提供新的可能。
