小球藻的基因工程改造研究进展
2024-01-06张甜甜陈必链郑明敏
张甜甜,陈必链,2,郑明敏,2*
(1. 福建师范大学生命科学学院,福建 福州 350117;2. 福建师范大学,工业微生物教育部工程研究中心,福建 福州 350117)
小球藻(Chlorella)是一种单细胞真核绿藻,具有光合作用能力,生长繁殖速度快,广泛分布在淡水、湖泊和海洋等水体中,易于大规模培养,是生产生物燃料[1]、高价值化合物[2-3]、水产养殖功能饵料及重组治疗蛋白[4]的潜在宿主。从改造的小球藻中,可以提取蛋白、油脂等营养物质。由于小球藻能够利用二氧化碳[5],因此其也被研究用于缓解温室效应等环境问题。
随着基因工程技术的发展,加上小球藻具有易培养、生长快速等优势,为其成为外源蛋白生产的理想宿主提供了可能性。许多研究者开始利用基因工程技术在小球藻细胞中进行外源基因的表达,以期生产外源蛋白或提高小球藻生产高附加值产品的能力,图1展示了运用不同方法进行小球藻的遗传转化操作。构建遗传转化体系是开展基因工程研究工作的基础,有研究表明,小球藻可以克服其他细菌、真菌、植物和动物表达系统在生产重组蛋白中的缺点[6]。
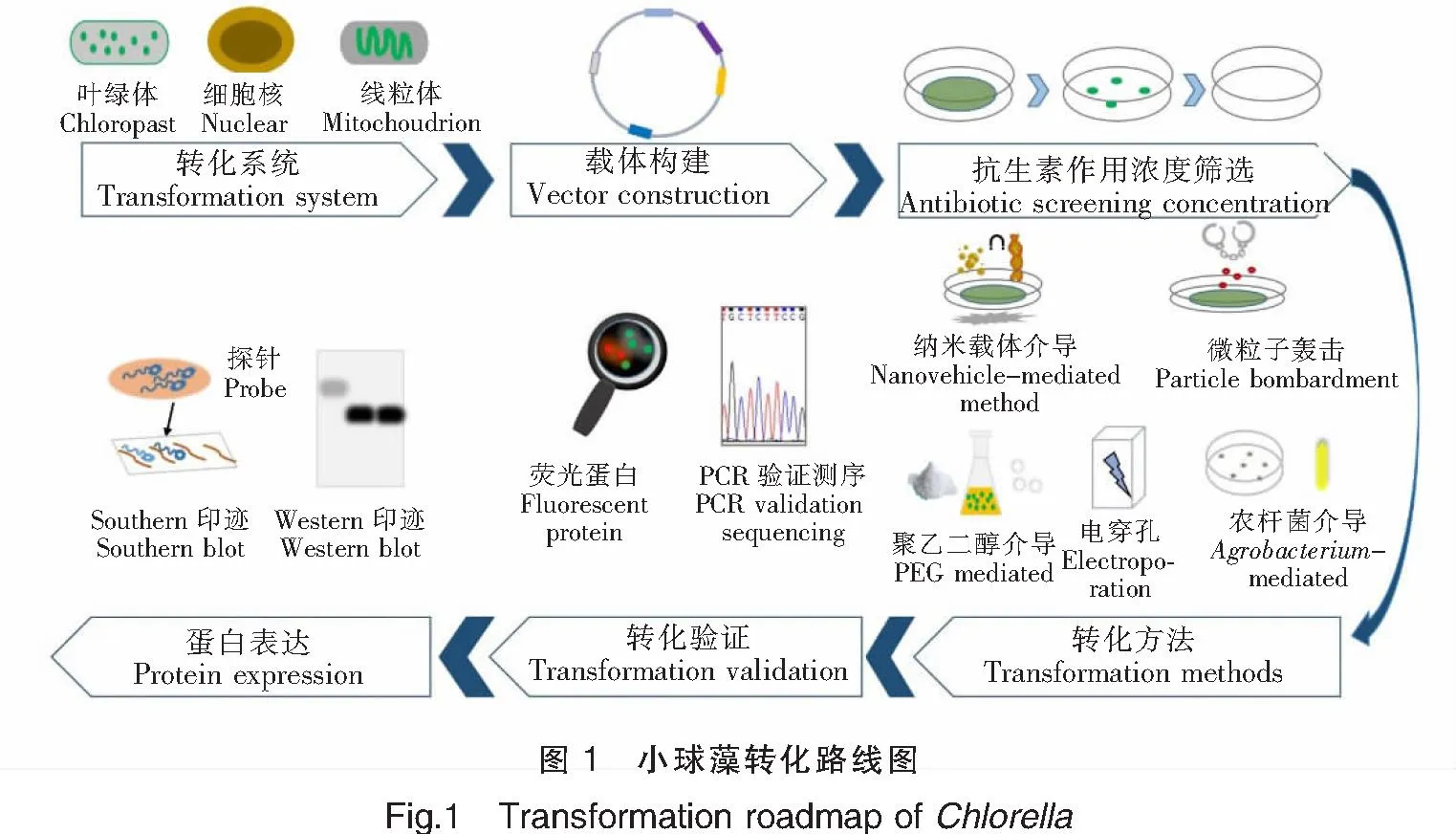
作为外源基因或蛋白表达的潜在宿主,小球藻具有良好的发展前景和优势。但由于缺乏可靠高效的转化体系,蛋白表达不稳定以及高价值化合物产量相对较低,阻碍了其作为生物反应器应用的发展。因此,了解小球藻基因工程转化体系的建立和发展,对今后的真核基因表达具有积极意义。本文阐述了近年来在真核微藻小球藻中的基因工程改造进展,详细介绍了小球藻遗传转化体系的转化系统、转化方法、影响转化的因素及相关蛋白的表达,以期为后续小球藻的基因工程改造和功能蛋白生产提供技术参考。
1 小球藻转化系统
微藻中小球藻已实现细胞核和叶绿体系统的转化开发,但对于线粒体系统的转化,仅在莱茵衣藻(Chlamydomonasreinhardtii)中实现。开发转化系统的难点在于需要同时克服四个问题:1)DNA能够被转移到胞内;2)至少需要表达一个标记或报告基因;3)宿主细胞内能够复制DNA;4)被转化的细胞能够恢复和增殖。除了核转化外,小球藻叶绿体转化也逐渐受到关注。针对不同的靶细胞器转化系统,需要制定合适的转化体系,包括转化方法、启动子、标记/报告基因的选择。
1.1 核转化系统
对于小球藻的遗传转化系统,核基因组转化是最早且被应用最广泛的方法。目前已有超过12种不同的小球藻藻种成功地进行了核转化[7]。由于在细胞核中表达的基因能够编码多种功能蛋白,与核基因组严格相关的功能不能通过细胞器基因工程来修改,因此细胞核成为许多基因工程研究的主要目标[8]。对于小球藻而言,相对于细胞器转化,目前人们对核基因组的转化更为了解,操作也更为方便。Kim J等[9]利用CRISPR-Cas9系统成功编辑了小球藻的核基因组,生成硝酸还原酶(Nitrate reductase,NR)和腺嘌呤磷酸核糖转移酶(Adenine phosphoribosyl transferase,APT)基因突变体。Gomma A E等[10]利用SV40大T抗原转化功能域,通过overlap PCR技术制备线性基因表达盒,实现了该抗原成分在小球藻细胞中的稳定表达,建立了一个无抗生素标记核转化系统。Run C等[11]构建含有增强型绿色荧光蛋白(eGFP)基因的载体,通过优化电穿孔转化的相关参数,在DNA水平上成功鉴定了NptⅡ和eGFP基因,实现了蛋白核小球藻的稳定核转化。虽然小球藻的核基因组转化表达系统已逐渐成熟,但其也存在易出现基因失活、基因沉默、表达效率低等情况[12]。
1.2 叶绿体转化系统
针对植物细胞而言,其遗传物质不仅存在于细胞核中,还存在于叶绿体和线粒体中。1988年Boynton J E等[13]使用轰击法成功实现了莱茵衣藻的叶绿体转化,为随后小球藻的叶绿体转化提供了可能。Wang K等[6]通过基因枪法将两个抗菌肽基因传递到普通小球藻(C.vulgaris)的叶绿体基因组中,成功构建了小球藻叶绿体转化体系。在叶绿体转化系统中,外源DNA两侧通常有与整合位点两端同源的区域,通过同源重组将外源基因整合到叶绿体基因组的特定位点上。需要注意的是,这些同源区域要足够短,以尽量减少自发重组。叶绿体转化系统具有基因拷贝数多、表达可靠、无基因沉默现象等优点,但外源DNA的进入需要穿过叶绿体膜等障碍,这可能是小球藻叶绿体转化体系还不够成熟的原因之一。
1.3 线粒体转化系统
关于动物和植物线粒体的研究已有所完善和进步,但在细胞体内稳定遗传的线粒体转化只在酿酒酵母(Saccharomycescerevisiae)和莱茵衣藻中可以实现[14]。Hu Z等[15]将编码增强型绿色荧光蛋白(Enhanced green fluorescent protein,EGFP)的基因转入莱茵衣藻线粒体中并实现了表达。而Yoo B C等[16]设计了一种携带外源DNA的“编辑质粒”,通过基因枪法将质粒成功导入酵母线粒体和莱茵衣藻叶绿体中。但是由于单个细胞可能存在多个线粒体及植物线粒体基因组通常也存在不稳定性、缺乏强选择性标记等[14]缺陷,线粒体转化系统也没有得到普遍应用。
2 转化方法
目前,已有关于小球藻的转化方法的研究,但选择合适的转化方法仍然是实现外源蛋白在小球藻中表达的关键步骤之一。1991年,Jarvis E E等[17]使用聚乙二醇(PEG)介导法在椭圆小球藻(C.ellipsoidea)中实现了荧光素酶(Luciferase)基因的瞬时表达,这是在小球藻中首次实现核转化。之后,对于小球藻转化方法的研究逐渐增多。通过开发并优化一种可靠高效的转化方法,可以大大提高小球藻的转化效率。本文总结了5种转化方法,分析并阐述其优缺点和应用。
2.1 聚乙二醇介导法
聚乙二醇(Polyethylene glycol mediated method,PEG)介导法,又被称为玻璃珠法,是通过利用聚乙二醇与玻璃珠协同作用,造成细胞膜产生瞬间空隙,从而使外源 DNA 得以进入细胞的方法[18]。PEG介导法使用设备简单、操作方便,对宿主细胞的损伤较小[7],是最早用于使外源基因转移到微藻的方法之一,最初被用于模式藻莱茵衣藻的转化[19],之后在一些小球藻种的转化体系中也得到了应用。Hawkins R L等[20]利用PEG介导法,将带有胞外分泌信号序列和外源DNA的质粒转化到小球藻细胞中,在小球藻中实现了人生长激素(Human growth hormone,HGH)基因的瞬时表达,并发现玻璃珠的大小也会影响转化效率。杨博[18]利用PEG介导法,首次将增强型绿色荧光蛋白报告基因转化到普通小球藻基因组中,最后获得(356±30)个克隆子/μg DNA,建立了一套稳定而可靠的小球藻外源基因转化体系。然而,PEG介导法的转化效率通常较低[21],且只能用于无细胞壁细胞。因此,在使用该方法时,需要选用无细胞壁突变体作为受体细胞,或使用物理、化学、生物等方法[22]去除细胞壁以获得原生质体。目前,多数研究采用复合酶解液或物理方法(如研磨或超声破碎)对小球藻的细胞壁进行破坏和去除。
2.2 电穿孔法
电穿孔是一种常用的细胞遗传转化方法,通过施加特定强度的脉冲电场,改变细胞膜的表面结构,在细胞膜上打开微孔,以便外源 DNA 通过此孔进入受体细胞,并在恢复状态后整合到细胞染色体组中[23]。电穿孔法是目前小球藻遗传转化研究中应用最为广泛的方法,具有操作简便、转化效率较高等优点。
电穿孔法的转化效率受到多种因素的影响,包括细胞壁、电场强度、脉冲时间、质粒浓度等。Chow K C等[24]发现,在较高的电场强度下,获得的转化效果更好,但是电压过高会导致重组质粒无法稳定整合到细胞染色体组中。Muoz C F等[25]在开发预测微藻最佳电穿孔条件的方法时发现:较低的细胞浓度、光强和小DNA片段的传递可以产生更高的转化效率。但有些种属,比如三角褐指藻(Phaeodactylumtricornutum)[26],由于其具有硅质化的细胞壁,因此无法使用电穿孔直接进行转化。针对这种情况,通常采用制备原生质体或使用渗透液渗透细胞壁,如使用山梨醇和甘露醇预先处理受体细胞,改变细胞壁通透性,以此增加质粒 DNA 进入细胞的概率。
2.3 微粒子轰击法
微粒子轰击法,又叫基因枪法,是一种可以将DNA递送到完整细胞的有效方法。它通过利用高压氦气或氮气瞬间产生的冲击力,向细胞中发射黏附有质粒DNA的金属(钨或金)微粒子,从而穿透细胞壁进入细胞[27]。Chen Y[28]等最初在椭圆小球藻中利用微粒子轰击实现了β-葡萄糖苷酸酶(β-Glucuronidase,GUS)基因的瞬时表达,表明微粒子轰击是一种用于小球藻转化的可行方法。Wang K等[6]通过直径约550 nm的金载体微颗粒将重组质粒轰击转化至普通小球藻的叶绿体中,获得了(212±48)个克隆子/μg DNA的转化值。赵熙宁[29]通过微粒子轰击介导蛋白核小球藻(C.pyrenoidosa)的转化,最终获得了高蛋白产量的优质蛋白核小球藻工程株。
微粒子轰击法的影响因素主要包括微粒子直径、轰击距离和DNA浓度等。虽然需要使用昂贵的基因枪设备,但操作简单、适用范围广。叶绿体基因组转化的唯一有效方法是微粒子轰击或生物法[19]。但使用的金属颗粒一般带有生物毒性,可能会对受体细胞产生损伤。
2.4 农杆菌介导法
农杆菌介导法是一种常用于植物遗传转化的方法,其原理是将目的基因插入到根瘤农杆菌(A.tumefaciens)Ti质粒上的T-DNA 区,通过农杆菌侵染将目的基因导入到植物细胞的核基因组中[30]。近些年来,研究发现该方法也可用于小球藻的遗传转化。Sharma P K等[31]首次报道了一种在索罗金小球藻(C.sorokiniana)中高效稳定的农杆菌介导转化体系,最终获得了(220±5)个克隆子/106细胞的转化值。冯兴标等[32]通过农杆菌介导法有效提高了小球藻中虾青素的含量。与其他方法相比,农杆菌介导法具有操作简便、宿主范围广泛、可转移大片段DNA,以及准确将低拷贝数转基因整合到转录活性区域等显著优势[33]。
在农杆菌附着植物细胞的过程中,植物细胞缺口会释放酚类化合物如乙酰丁香酮(Acetosyringone,AS),然后诱导毒力基因Vir进行表达,从而将外源DNA转移到细胞基因组中[30]。Yang B等[7]研究发现小球藻只有在加入AS后才能进行转化。Sharif N等[34]的研究结果表明,细菌密度为OD600=1.0、共培养温度为25 ℃、共培养培养基pH为5.5、共培养3 d、乙酰丁香酮浓度为100 μmol/L是农杆菌转化普通小球藻的最佳条件。在农杆菌介导的转化体系中,可对农杆菌浓度、小球藻细胞状态、共培养条件及乙酰丁香酮浓度等参数进行优化,以提高小球藻的转化效率。
2.5 纳米载体介导法
纳米载体介导法是一种新型的小球藻遗传转化方法。磁性纳米粒子是从金属氧化物中制备的基因载体。它可以通过修饰表面与大量核酸分子静电吸附结合,形成复合体,在磁场作用下渗透到细胞中,从而将目的基因传递到核基因组中。曹苏珊等[35]使用磁性纳米颗粒介导法成功将报告基因EGFP传递到核基因组,在椭圆小球藻中获得了EGFP绿色荧光的瞬时表达。磁性纳米颗粒与质粒紧密结合形成的静电吸附可以防止外源基因被胞内核酸酶酶解,从而避免了细胞转化效率的降低[36]。作为一种新型的转化方法,该技术具有转化速度快、操作方便、表达稳定等特点,但也存在转化效率低、成本高等缺陷。
3 转化影响因素
3.1 细胞壁
微藻的细胞壁是由碳水化合物、碳氢化合物、蛋白质和其他成分组成的复杂多聚体[8]。小球藻的细胞壁随着生长发育会逐渐形成两层结构,厚度约为17~21 nm[37-38],这种细胞壁是外源DNA进入微藻细胞的一个巨大物理屏障。
若使用农杆菌或PEG介导法进行转化,需要获得细胞壁缺陷型藻株或制备原生质体。酶解法是制备藻细胞原生质体的常用方法,具有条件温和、对细胞损伤程度小等优点。纤维素是大多数小球藻种细胞壁的主要成分[39]。研究表明,同时使用纤维素酶和崩溃酶预处理小球藻,可以显著提高原生质体制备率[39]。Yang B等[5]使用了含有纤维素酶、离析酶和果胶酶的酶混合液预处理小球藻的细胞壁,最终得到了超过80%的原生质体产率。尽管破坏细胞壁不是所有藻类成功转化的必要条件,但它是细胞渗透到足以有效穿透外源DNA和细胞死亡之间的微妙平衡[8],可以有效提高转化效率。在莱茵衣藻中,高效率转化通常只报道在无细胞壁突变体或已去除细胞壁的原生质体细胞中[8]。因此,在小球藻基因转化研究中,细胞壁的处理是提高转化效率的关键步骤之一。
3.2 选择标记和报告基因
在转化体系中,通常会将带有某一特定抗性的选择标记或带有荧光报告基因的质粒与目的DNA融合后,转化到细胞中,便于后续阳性转化子的筛选。选择标记是指将编码某一抗生素的抗性基因导入细胞中表达,从而使转化细胞具有对该抗生素的抗性特质。表1列出了目前常用于小球藻遗传转化筛选的抗生素抗性选择标记。
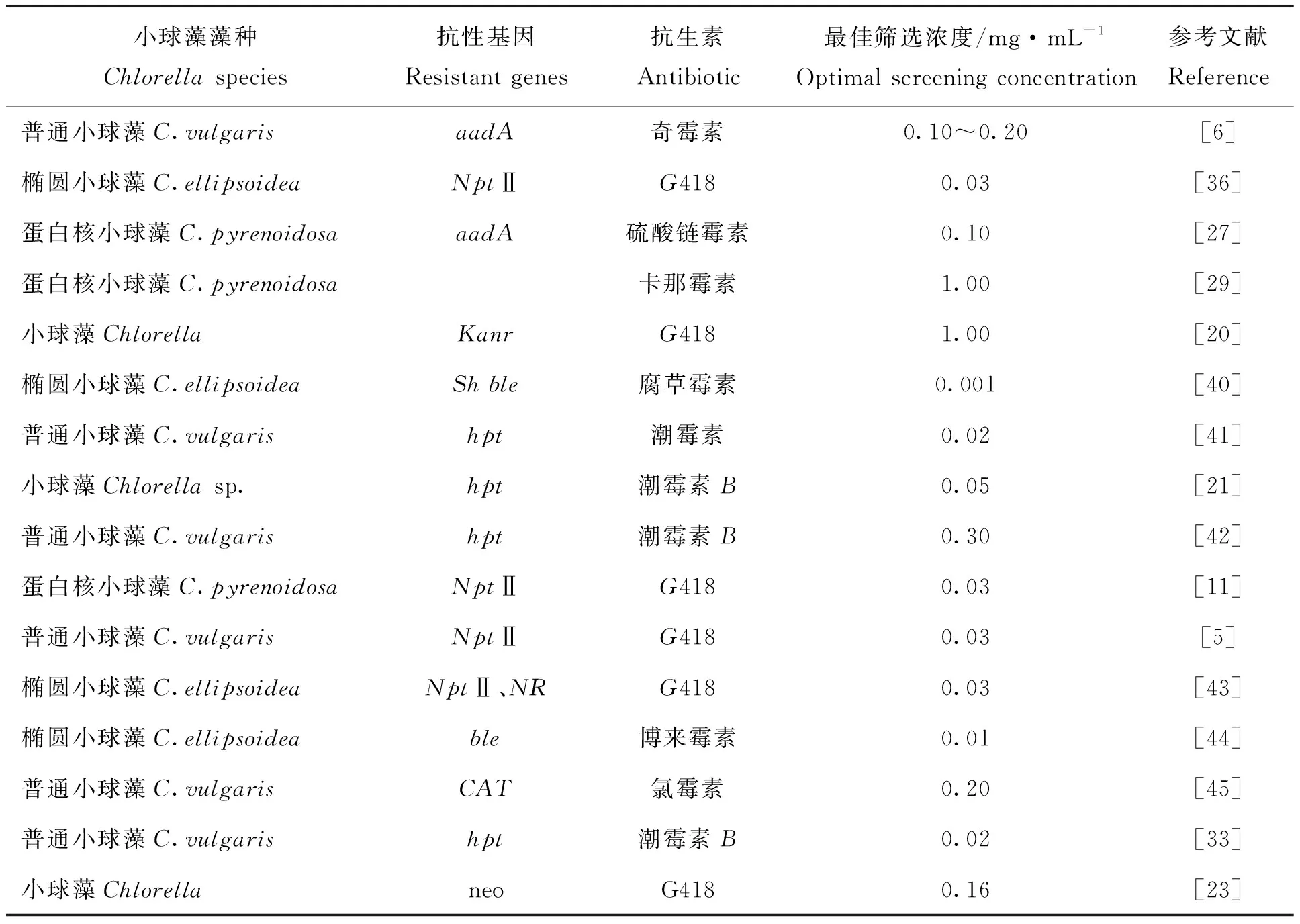
表1 用于不同种类小球藻的抗生素抗性基因及最佳筛选浓度Tab.1 The resistant genes and optimal screening concentration of antibiotic for different species of Chlorella
不同的藻种对抗生素有不同的敏感浓度,在选择抗生素时,需要提前考虑藻的种类和其本身的抗性,并设置一系列抗生素浓度梯度,观察抑制野生型藻株生长的最佳作用浓度,这是小球藻基因工程改造中的重要部分。
报告基因是指编码具有酶活性或易于从各种细胞蛋白中识别的蛋白质[7]。例如在荧光显微镜下发出绿色荧光的绿色荧光蛋白[11]、增强型绿色荧光蛋白和蓝绿色荧光蛋白[38]。成功表达荧光报告基因的细胞,在一定的激发波长下,会发出相应的荧光,以此来验证质粒是否整合成功。但考虑到小球藻细胞具有固有的光合色素和一些荧光物质,如叶绿体红色荧光[46],因此荧光报告基因需要在强启动子的驱动下才能达到高表达水平,以此区别于背景荧光。
除了肉眼可以观察到的显性选择标记外,还有一些隐性选择标记可用于筛选转化子。NR基因编码了一种介导硝酸盐向亚硝酸盐转化的酶,NR基因突变体不能利用硝酸盐,因此在仅以硝酸盐为氮源的培养基中无法生长。另外,APT基因编码一种催化腺嘌呤转化为腺苷酸的蛋白质,APT突变体会导致细胞在含有2-氟腺嘌呤的培养基中不受抑制地生长[9]。这种通过基因突变导致翻译提前终止,蛋白无法表达的隐性选择标记可以减少外源基因的插入和抗生素的使用,具有提高经济效益和减少环境污染等优点。
3.3 启动子和增强子
启动子是基因表达的关键因子,强启动子会调控基因的高表达[47]。花椰菜花叶病毒35S (CaMV35S)启动子是小球藻中应用最广泛的组成型启动子之一[7],已被成功用于普通小球藻[5]、椭圆小球藻[17]、蛋白核小球藻[48]。来自植物的组成型启动子也可以发挥作用。如玉米的泛素启动子已成功驱动GUS报告基因在小球藻中的高效表达[44],而水稻肌动蛋白的Actin1启动子也被认为可以促进小球藻中的基因表达。Niu Y F等[45]克隆了NR基因的启动子和终止子,驱动诱导氯霉素乙酰转移酶(Chloramphenicol acetyltransferase,CAT)报告基因的表达,结果显示CAT的表达会受到硝酸盐的诱导,提出了一种诱导型启动子在小球藻中的应用体系。
一般来说,外源启动子的表达效率低于内源启动子。在小球藻中被鉴定并分离出的光系统Ⅰ蛋白D (psaD)启动子,是首次报道的可以驱动真核微藻和高等植物基因表达[49]的小球藻内源性启动子。Shin J H等[50]从小球藻核基因组中分离到两个氮饥饿诱导型启动子,通过添加信号肽,成功实现了人粒细胞集落刺激因子(hG-CSF)多肽的表达和分泌。为了高效驱动基因表达,研究者会在构建质粒时加入一些调控元件,如增强子来激活启动子发挥功能。研究表明在中性粒细胞肽(Neutrophil peptide-1,NP-1)基因上游区域添加一个ω增强子后,可以增加基因表达[51]。在Liu J等[52]构建的小球藻(C.zofingiensis)转化体系中,将内源性八氢番茄红素去饱和酶启动子的第一个内含子序列克隆后,添加到启动子序列上游,转化效率提高了91%。这些研究证实了增强子或调控元件在促进小球藻转基因表达中可以发挥重要的作用。
4 蛋白表达
小球藻拥有完整的真核细胞表达系统,因此可以对外源蛋白进行翻译后修饰和加工。此外,小球藻具有生长周期短、食用安全性高、生产效率高等优势,被广泛应用于基因工程领域。表2列出了不同种类小球藻转化系统中的转化率或蛋白表达量。
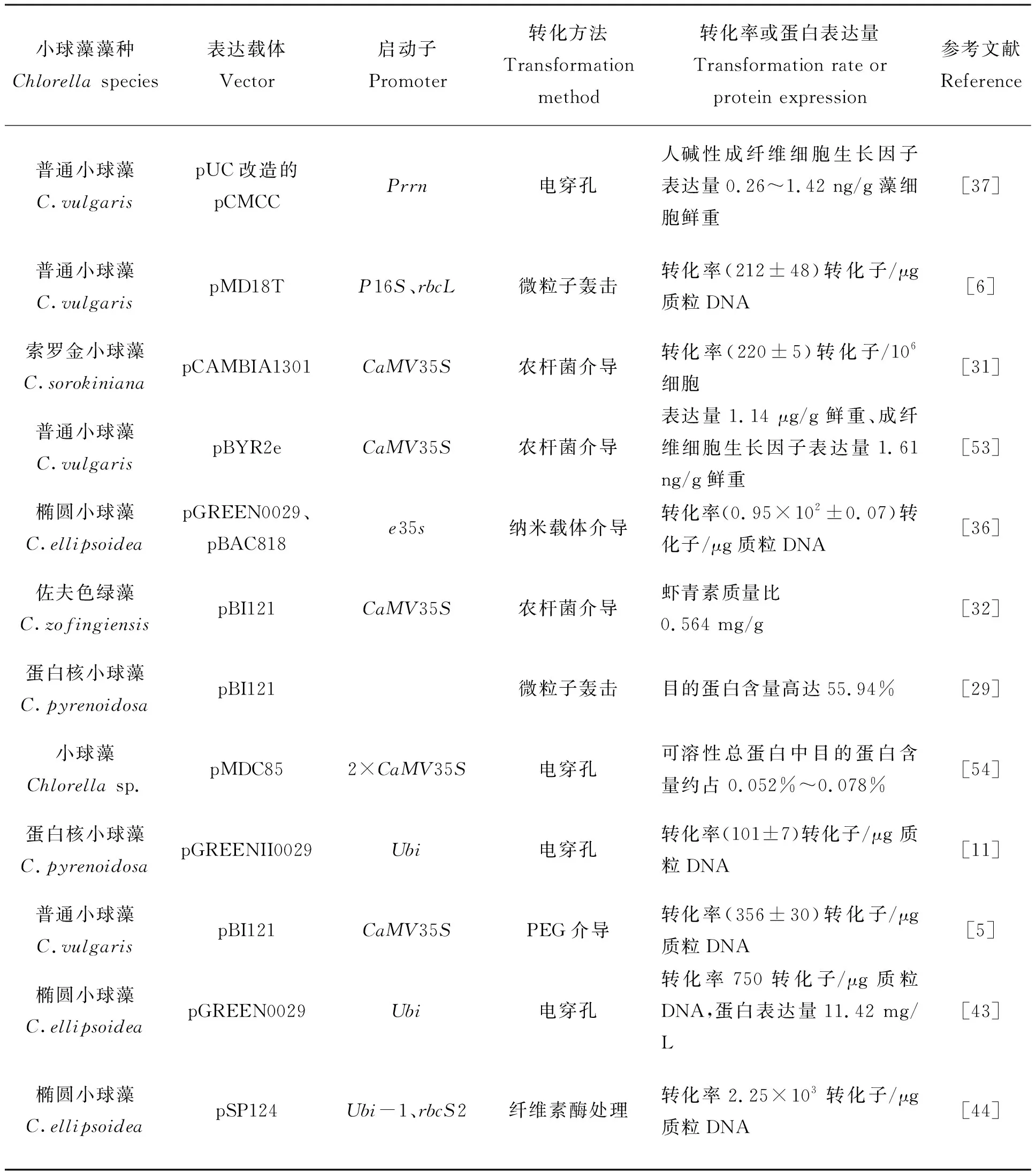
表2 不同种类小球藻的转化率或蛋白表达量Tab.2 The transformation rate or expressed proteins in different species of Chlorella
Hawkins R L等[20]发现在小球藻中表达的异源蛋白HGH的产量约为200~600 ng/mL。虽然比报道的大肠杆菌(Escherichiacoli)中的HGH(0.5~2.4 μg/mL)产量低。但该研究为在小球藻中表达生产外源蛋白提供了良好的开端。
小球藻中表达的重组蛋白可用于生产天然饵料。Kim D H等[40]在椭圆小球藻中稳定转化了比目鱼生长激素(Flounder growth hormone,fGH)基因,最后测定fGH的蛋白量超过了400 μg。将转化后的小球藻喂养比目鱼30 d后,其生长速度提高了25%。Reddy P H等[48]将含有传染性法氏囊病病毒(Infectious bursal disease virus,IBDV)VP2基因的表达载体转化到蛋白核小球藻中,Western blot分析证实了IBDV VP2蛋白可以在蛋白核小球藻中表达,该蛋白可以保护鸡免受鸡传染病病毒IBDV的侵害。这些研究证实了小球藻中外源蛋白稳定表达的可行性,为工业化生产水产养殖业的食用蛋白提供了可靠依据。
目前也有许多针对小球藻进行的外源抗菌肽的表达研究。Koo J等[41]将牛乳铁蛋白N叶基因进行密码子优化,在普通小球藻中实现了乳铁蛋白的稳定表达。乳铁蛋白是一种可以在非免疫防御系统中起作用的抗菌肽,对革兰氏阳性和阴性细菌都具有抗菌活性。何艺宾[54]利用电穿孔转化法,在转基因小球藻中实现了抗菌肽融合基因的稳定表达,并证实了该抗菌肽能够抑制嗜水气单胞菌(Aeromonashydrophila)和金黄色葡萄球菌(Staphylococcusaureus)的生长。
小球藻作为表达治疗性蛋白质的潜在平台也引起了人们的关注。Bai L L等[43]将突变的防御素(Mutated NP-1)NP-1基因转化到椭圆小球藻中,并分离出了具有生物活性的防御素。藏花酸是稀有药用植物藏红花的主要活性成分之一,是一种潜在的药用药物。Lou S等[42]通过将藏花酸合成途径中的关键基因crtRB和ZCD1引入普通小球藻中,成功产生了具有活性的藏花酸。虽然小球藻能够进行复杂的蛋白质折叠和修饰,产生的活性蛋白质可以用作抗原和抗体[55],但其生产量往往低于细菌或植物系统。适当添加信号肽序列[50]和核糖体结合位点(RBS)序列[37]或许有助于建立高效的蛋白表达和分泌系统。
CRISPR/Cas9通过使用gRNA序列将核酸酶(Cas9)引导到受体细胞的基因组中,可以精确编辑原核和真核物种的基因组。尽管在莱茵衣藻中首次成功应用了CRISPR/Cas9介导的基因编辑技术,但结果显示其转化效率很低,这可能是因为Cas9对藻细胞具有细胞毒性[56]。除了较低的转化效率外,需要用于筛选的明显表型也是目前CRISPR/Cas9基因编辑系统在微藻中使用的局限之一[8]。通过蛋白质组学技术[56]和高效基因表达框的改良,可能有利于基因编辑技术在小球藻转化中的应用。
5 总结与展望
小球藻是高值生物产物和功能蛋白质的一种重要来源。通过利用各种技术工具对小球藻进行基因组编辑和蛋白表达,扩展其作为表达宿主,具有经济性和环境可行性。目前,电穿孔和基因枪法是小球藻转化的常用方法,其中基因枪法是用于叶绿体基因组转化的最有效方法。常用于植物转化系统的农杆菌介导法,不需要提前制备原生质体,在小球藻转化体系中也有巨大的应用潜力。启动子、功能表达载体和合适的选择标记是小球藻转化体系的重要组成部分。构建和利用植物病毒载体,密码子优化、插入调控元件、使用内源性启动子等操作可以提高外源基因插入的效率和稳定性,并增加蛋白表达量。
小球藻作为一种生命活动简单的真核微藻,在功能营养饵料和改善生态环境方面都有可观的前景。尽管不断的探索和研究使小球藻基因工程改造领域取得了相当大的进展,但在其成为成熟的表达系统之前仍有许多障碍需要克服。开发一种适合小球藻高效、稳定表达蛋白的转化体系,对于生产高价值产品和功能性营养品具有重要而深远的意义。
