山西高原主要林型生物量异速生长模型
2024-01-05严俊霞段兰兰
贺 丽, 严俊霞†, 段兰兰, 刘 菊, 王 琰
(1.山西大学黄土高原研究所,030006,太原; 2.山西省林业和草原科学研究院,030012,太原)
森林生态系统对水土保持和水源涵养起着重要作用,也是陆地生态系统最大的碳库,储存陆地生态系统有机碳的76%[1]。森林生物量的准确估算对评估森林在全球气候变化中的减排效应、预估碳汇潜力以及增加区域碳收支都具有重要意义[2]。
森林生物量估算方法可分为直接和间接估算。前者准确性高,但费时费力、成本高,对森林破坏性大,不适合大面积森林生物量的估算[2]。异速生长模型作为一种间接方法,通过易测因子(如树干直径和高度)来估算树木的整体或部分生物量[3],被广泛运用于不同森林生态系统生物量的估算中[4]。
森林生物量的大小及其分配不仅与树种有关,而且受立地条件、气候条件以及海拔等因素的影响。即使在同一区域,不同森林类型的生物量模型的参数亦可能有较大的差异;因此,建立适用于特定地区、特定环境条件下的生物量生长模型,对准确估算地区森林生物量、减少估算误差非常重要。利用异速生长模型估算森林生物量的研究较多,我国不少学者针对全国和区域尺度建立生物量方程[5-7]。山西地处黄土高原东部,全省森林面积321.1万hm2。准确估算山西高原森林生物量和碳储量对于山西乃至全国的碳收支估算都具有现实意义;然而,利用异速生长模型对山西高原不同植被生物量估算的整体研究仍比较缺乏。已有研究多集中在太岳山的华北落叶松、油松、辽东栎等[8-9]。笔者以山西高原10个森林生态站典型树种的解析木数据为基础,研究不同树种不同组分的生物量分配比例;利用典型变量,即胸径(D)和树高(H),为山西高原不同林型建立生物量异速生长方程;比较模型的拟合精度,得到不同林型生物量估算的最优模型。
1 研究区概况
山西省位于华北地区(E 110°14′~114°33′、N 34°34′~40°44′),总面积15.67万km2。地势表现为由东北斜向西南的平行四边形,以山地丘陵为主。土壤类型主要为栗钙土、栗褐土和褐土。气候属温带大陆性季风气候,雨热同期,光照充足。年平均气温介于4~14 ℃之间。年降雨量358~621 mm之间,多集中在6—8月。无霜期120~220 d。山西省植物资源丰富,植被垂直地带性和纬度地带性特征明显,主要森林类型有针叶林(寒温性针叶林和温性针叶林)、针阔叶混交林、落叶阔叶林[10]。
2 材料与方法
2.1 样地设置
山西省生态林业区划将全省划分为北部风沙区、西部黄土丘陵区、吕梁山土石山区、中南部盆地区、东部土石山区5个一级区[11],其中中南部盆地区大中城市相对集中,不是山西森林的主要分布区域,在确定森林生态定位网络监测时,不作为主要森林生态类型区;将其余4个生态林业区根据立地类型、平均气温(T<7.5 ℃、≥7.5 ℃~10 ℃、>10 ℃)和年均降雨量(P<400、≥400~500和>500 mm)梯度分为11个森林生态类型区(图1);依据森林生态类型区的森林资源分布特点、《森林生态站工程项目建设标准(试行)》,建立10个省级森林生态站,与3个国家森林生态站共同构成山西森林生态系统监测网络体系[12]。13个森林生态站中,3个国家站的森林类型已在10个省级站中全部涵盖,因此笔者的试验样地依托10个省级森林生态站进行。
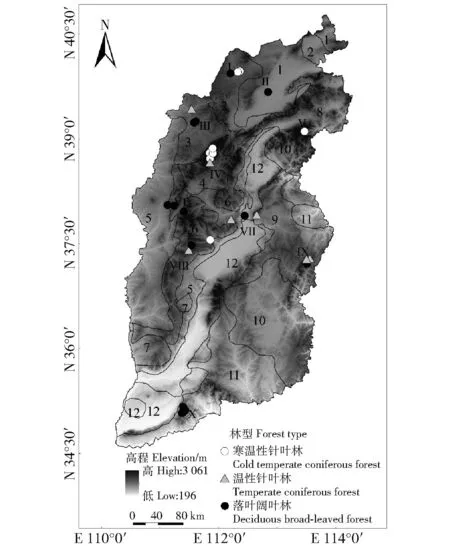
1~12为北部风沙区(1、2),西部黄土丘陵区(3、5、7),吕梁山土石山区(4、6),东部土石山区(8、9、10、11)和盆地(12);Ⅰ~Ⅹ依次为右玉辅站、金沙滩杨树局站、偏关辅站、芦芽山站、五台辅站、临县辅站、太原站、关帝山站、太行辅站和中条山站。The number 1-12 indicate the northern sandstorm area ( 1 and 2), the western loess hilly area (3, 5 and 7), the Lüliang Mountain soil and stone mountain area (4 and 6), the eastern soil and stone mountain area (8, 9, 10 and 11), and basin area (12) , respectively. 2): Ⅰ-Ⅹ represent Youyufu station, Yangshuju station of Jinshatan, Pianguanfu station, Luyashan station, Wutaifu station, Linxianfu station, Taiyuan station, Guandishan station, Taihangfu station and Zhongtiaoshan station, respectively. 图1 采样点示意图Fig.1 Schematic diagram of the sampling sites
10个省级森林生态站包括山西省全部森林生态类型,主要优势树种(组)面积占森林生态区所有植被类型的88.59%(不包括灌木林区)[12]。因此在每个生态站根据优势树种(组)的种类与数量,设置3~8个面积为1 hm2的试验样地,10个生态站共设置50个固定的标准试验样地,涵盖山西省的主要森林类型,样地内主要植被类型优势树种的样木数量占所有样地样木数量的89.19%[12]。
2.2 采样方法
在减少破坏的前提下,充分考虑代表性,结合各样地优势树种的起源和生长环境、各龄级树种数量及分布面积,在每个试验样地内选取1个 20 m×20 m的代表性区域,进行每木检尺,记录树种、胸径、树高。根据所记录树种的种类和数量、胸径和树高的中位数值,在涵盖山西省主要森林植被类型所包含9个优势树种的基础上,按照样地中不同龄级树种的分配比例,在每个样地选取 3~4 株样木进行破坏性采样,50个样地共选取典型性样木97棵,包含山西省3个森林植被类型(寒温性针叶林(CCF)、温性针叶林(TCF)、落叶阔叶林(DBF))9个主要优势树种的不同龄级阶段。对所有97棵样木解析后,把不同森林生态类型区的相同树种合到一起进行统计分析。所建立的模型力求能代表山西森林不同林型的树种生物量。各树种样木的基本信息见表1。
各样木的地上部分生物量采用收获法:将树木进行破坏性采样,以树桩高度10 cm为界分为地上和地下2部分。地下部分生物量测定采用全挖法:在树桩周围挖掘半径1.5 m、深1.5 m的圆柱体,收集圆柱体内的所有根系。将树干在1.3 m处切割,此后每隔2 m一段进行分隔,直到树冠。在现场对所有树干、树根、树冠分别进行称量鲜质量。然后树根和树冠选取200~500 g样品、每段树干选取底端3~5 cm厚的圆盘样品分别称量鲜质量,带回室内烘干后称取干质量,分别计算树干、树根、树冠的含水率。利用含水率分别计算样木的树干、树根和树冠干质量,即生物量。所有取样树种分为3种林型、9个树种(表2),分别进行不同部位生物量与胸径、树高因子的关系方程拟合。
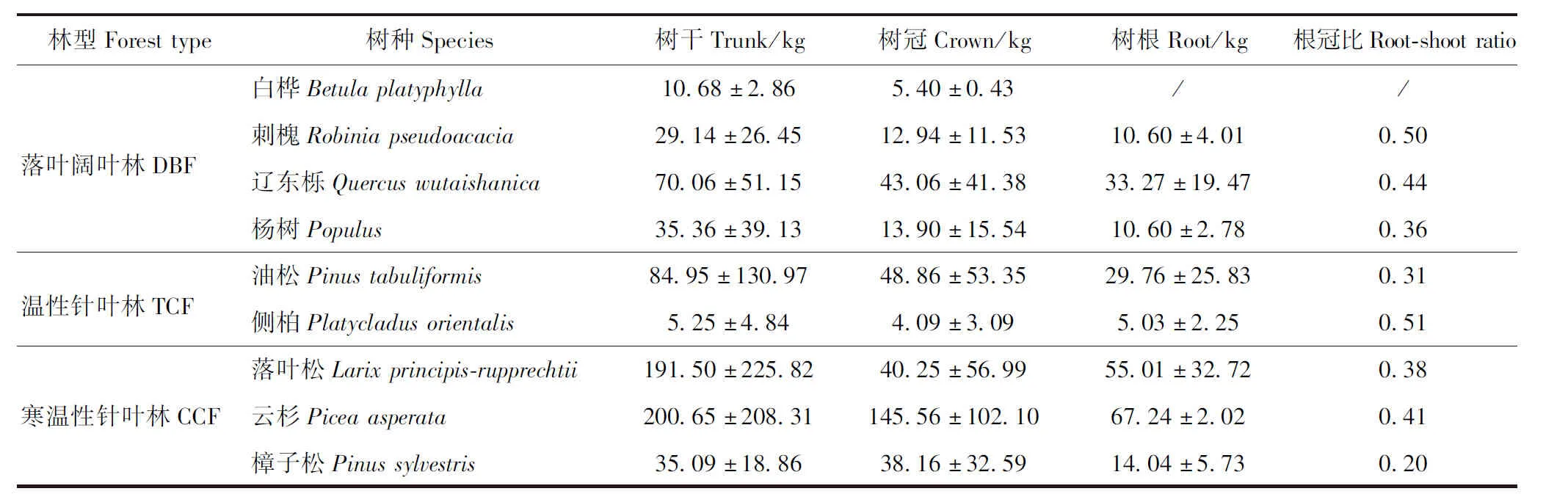
表2 各树种不同部位生物量Tab.2 Biomasses of different parts of the tree species
2.3 研究方法
笔者采用生物量估算常用的幂函数模型分别对树木不同部位的生物量与胸径、树高的关系进行拟合:
lnW=a+blnD;
(1)
lnW=a+bln(D2H);
(2)
lnW=a+blnD+clnH;
(3)
lnW=a+blnH。
(4)
式中:W为不同部位的生物量,kg;D为胸径,cm;H为树高,m;a、b、c为拟合参数。采用决定系数(R2)、均方根误差(Root mean squared error,RMSE)、平均绝对误差(Mean absolute error,MAE)、赤池信息准则(Akaike information criterion,AIC)、偏差(Bias)指标选择最优的生物量估算模型。根冠比(Root-shoot ratio,RS)反映地下和地上生物量的分配比例 ,用地上和地下生物量的比值来表示。
数据分析采用SYSTAT 12和SPSS 22.0软件,图形绘制采用Origin 2018软件。
3 结果与分析
3.1 各组分生物量分配
除樟子松外,其他8种树种生物量分配比例均为树干最大,但树种间有较大差异(图2)。8种树种树干生物量占总生物量的比例范围为37.05%~61.64%,平均为47.22%;其中,落叶松最大,为61.64%。树冠和树根生物量的比例在不同树种间差异相对较小;总体看,油松、云杉、樟子松和辽东栎树冠生物量占总生物量的比例大于树根的比例。不同树种树冠和树根生物量占总生物量的比例范围分别为8.98%~46.27%和16.68%~43.80%,平均值分别为25.59%和46.27%;落叶松树冠比例最小;刺槐的树根生物量比例在8种树种中最大,为43.80%,樟子松最小,为16.68%。不同树种根冠比例范围为11.99%~88.00%,其中侧柏比例最大,樟子松最小;地上部分占总生物量范围为56.20%~83.32%,其中刺槐比例最小,樟子松最大。随着胸径的增加,树干、树冠和树根生物量呈增加趋势(图3)。
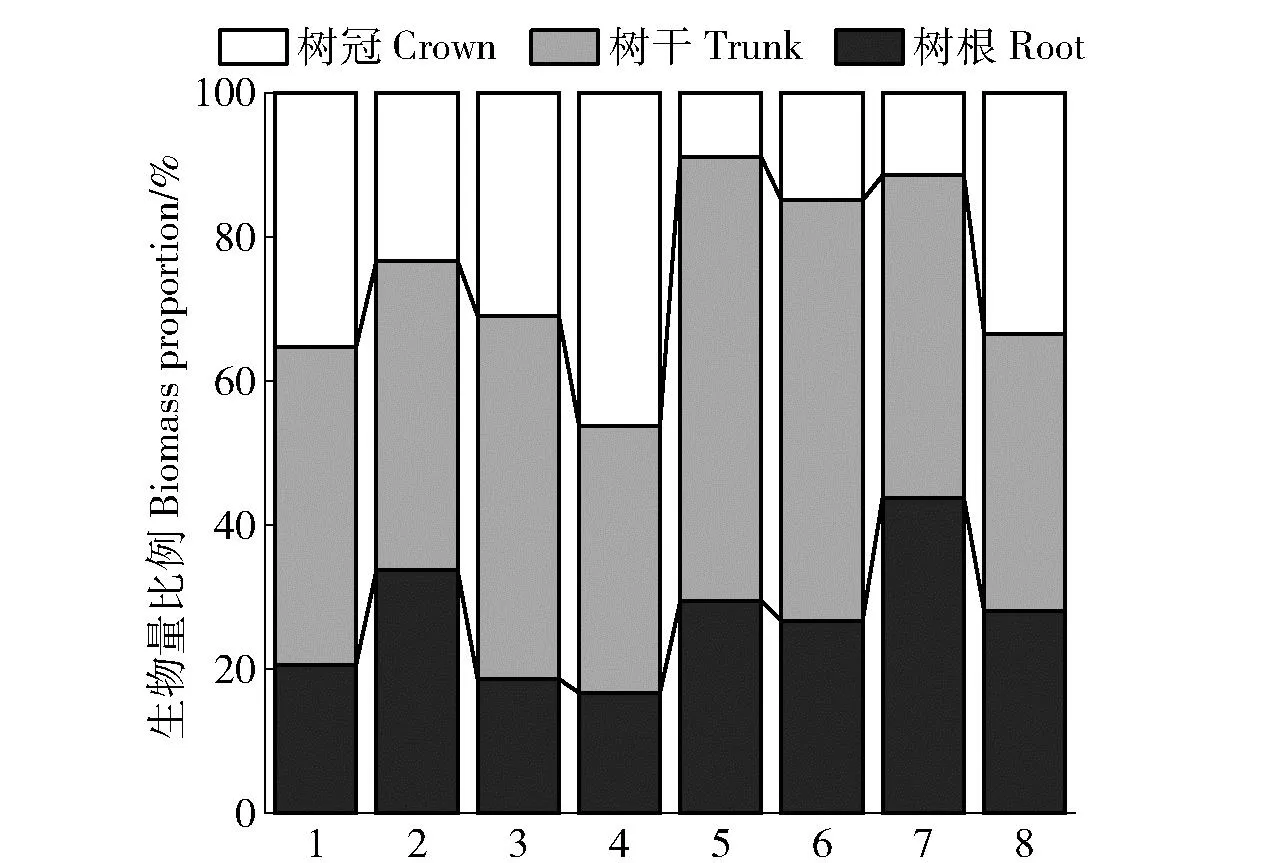
1~8分别为油松、侧柏、云杉、樟子松、落叶松、杨树、刺槐和辽东栎。The 1-8 indicate the Pinus tabuliformis, Platycladus orientalis, Picea asperata, Pinus sylvestris, Larix principis-rupprechtii, Populus, Robinia pseudoacacia and Quercus wutaishanica, respectively图2 不同树种生物量分配比例Fig.2 Biomass allocation ratios of different tree species

图3 树冠、树干和树根生物量与胸径之间的关系Fig.3 Relationship between biomass of crown, trunk and root and DBH
3.2 生物量异速生长模型
采用异速生长方程估算树干和树冠生物量时,用D、H及其组合作为预测变量的模型均达到极显著水平(P<0.001),可以解释树干和树冠生物量变化的65%~98%(表3)和34%~88%(表4)。不同林型的最优模型表现形式不一致,3种林型树干的最优模型分别为模型3(DBF和CCF)和模型2(TCF),树冠的最优模型均为模型3。

表3 不同林型树干生物量异速生长模型Tab.3 Allometric model of trunk biomass in different forest type

表4 不同林型树冠生物量异速生长模型Tab.4 Allometric model of canopy biomass in different forest type
由于树根生物量取样数据量少于树干和树冠的数据,因此分为DBF和针叶林(CF)2种林型计算分析。各种模型中,DBF只有模型4达到显著水平(P<0.05),而CF的4个模型均达到显著性水平(P<0.05),D和H以及它们的组合对根生物量的解释能力在54%~70%之间(表5)。对于CF而言,模型3的MAE最低,表明模型3是估算CF树根生物量的最优模型。

表5 不同林型树根生物量异速生长模型Tab.5 Allometric model of root biomass in different forest type
3.3 生物量模型拟合结果对比
为验证本文模型的精度,将结果与文献[7]的估算结果进行对比,笔者利用TCF和CCF的所有数据筛选出针叶林树干和树冠的生物量最优估算模型分别为lnW=0.04ln(D2H)-2.10和lnW=2.37lnD-0.92lnH-0.83。将笔者筛选的最优模型和李海奎等[7]建立的中国森林生物量模型得到的预测值与解析木实测值进行比较(图4)。结果表明,本研究的模型和中国森林的模型表现出不同程度的高估或低估。在不同林型中,本研究的模型对生物量的预测都偏高,Bias在0.54%~7.08%之间;在DBF中,中国森林的模型对树干和树根的生物量预测偏低,对树冠生物量的预测偏高,在针叶林中,中国森林的模型对树冠和树根生物量的预测偏低,对树干生物量的预测偏高,不同林型的Bias在0.63%~30.00%之间。在不同部位,模型均达到显著性水平(P<0.05)。本研究的模型拟合的R2均比中国森林的高,估算的偏差(Bias)比中国森林的更低,更接近于生物量实测值(图4)。
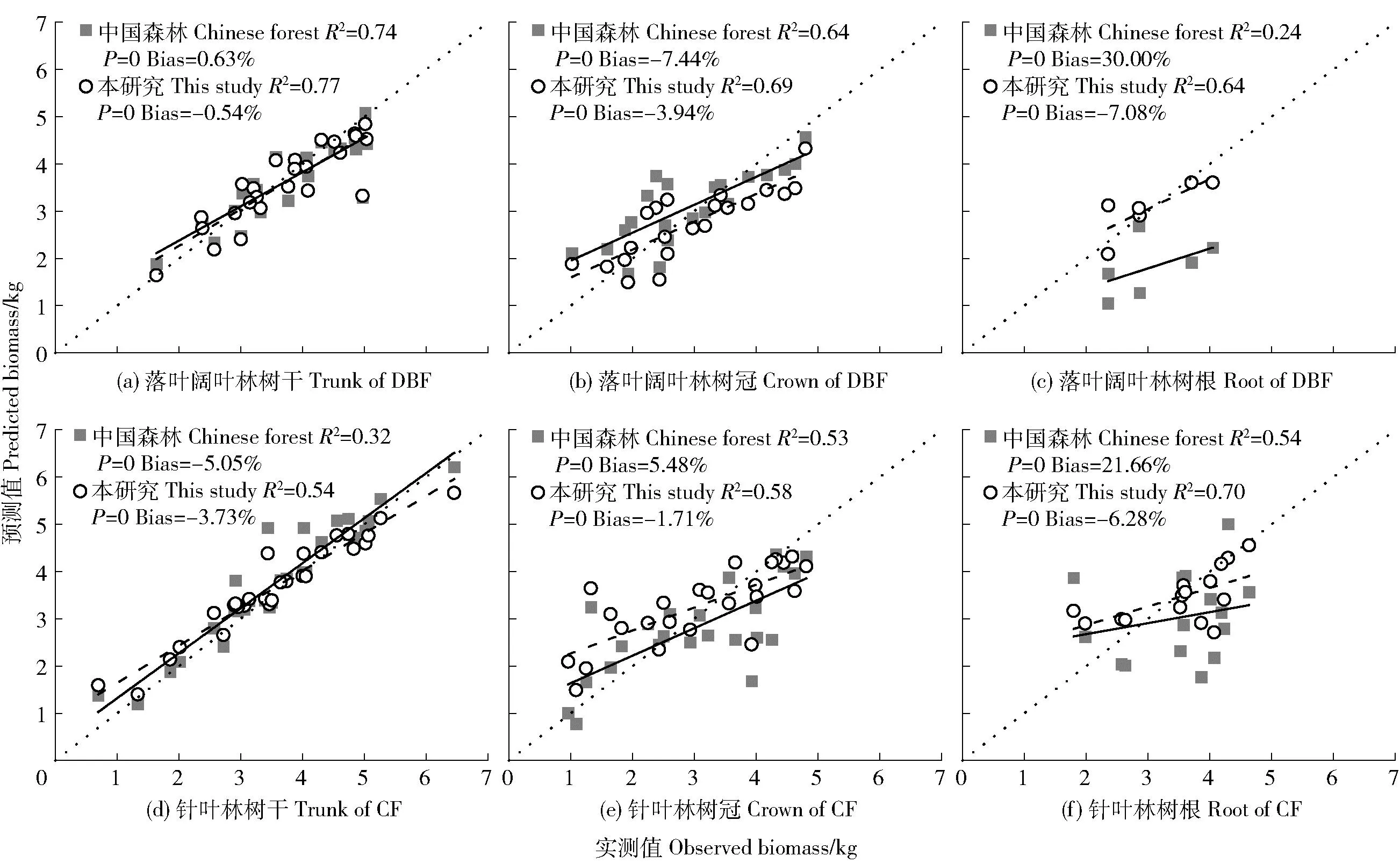
图4 落叶阔叶林和针叶林树干、树冠和树根生物量模型预测与实测值对比Fig.4 Comparison between the simulated and measured biomasses of tree trunk, crown and root in DBF and CF
4 讨论
4.1 各组分生物量分配
森林生物量受生物和非生物因素的共同影响[13],其在各组分的分配通常受到外部环境、树种差异、林龄等因素的影响。低温能降低土壤养分和水分的可利用性,引起植物根系生物量和分配比例的增加[14];Reich等[15]分析大量的森林生物量数据后,认为森林在低温环境下会优先将生物量分配给根。本研究中不同树种根冠比为11.99%~88.00%,高于Houghton等[16]对巴西亚马逊森林生物量根冠比的研究(13.4%~26.3%),主要是因为研究区处于中纬度内陆地区,属温带大陆性季风气候,年均温低于巴西亚马逊森林生态系统,地上部分生长较慢,根冠比相对较大。
关帝山、五台山中山和亚高山、五台山山间盆地、塞罕坝4个典型分布区落叶松的比例范围分别为46.32%~62.55%、12.03%~28.06%、7.77%~10.85%和13.46%~15.62%[17]。笔者研究中树干部分积累较多的生物量,其占总生物量平均比例为47.22%,其次是树冠和树根,分别占25.59%和27.18%,树干生物量占地上生物量比例较大。随着树木生长,更多的生物量将分配到树干上,使树木更加稳定,并通过树干将更多的水分和养分输送到需要进行光合作用的组分中[2],而且有利于占据林冠上层空间[13];朱江等[8]对山西太岳山落叶松生物量格局的研究发现,随着胸径的增加,地上生物量和地下生物量呈上升趋势,表明树种在幼树时将更多的生物量分配给树干快速到达林冠层,到达一定高度后分配更多的生物量给树枝以获取更多光照[3]。地上不同部位生物量逐渐增加,而根系为维持地上部分的稳定而逐渐生长,树根生物量也逐渐增加。
4.2 异速生长模型
在异速生长模型中,D作为最常用和最容易被精确测量的指标被广泛应用。加入其他变量如H,通常会提高模型估算的准确性[18]。然而在一些森林中,由于树高的测量具有较大的不确定性,对于模型中加入H来预测树木生物量的影响存在争议。一些研究认为没有改善,甚至恶化[19],而有些研究则认为会改善[20]。汪珍川等[21]对广西省主要树种生物量异速生长模型的研究表明,由于D信息获取简单且精确,利用D可较好地预测广西10类主要树种(组)各器官生物量;Chave等[4]发现,与只包含D为自变量的生物量模型相比,加入H后估算生物量的误差从16%降低到6%。笔者通过筛选模型发现,加入H后3种林型各组分的生物量异速生长模型的精度都得到不同程度的提高,不同林型以D2H或D+H为自变量有较好的拟合效果。
笔者建立的树冠生物量异速生长模型解释率低于树干。这是由于在野外采样过程中,树枝和树叶比树干更加复杂,测定比较困难[3],树叶对于光照、水分、养分和土壤条件高度敏感[22],其生物量受到树冠密度和枝条位置的影响,导致误差较大,树干生物量积累的结构组织上相对较高,而在枝叶上相对较低。随着树干的绝对质量逐渐增加,枝叶的绝对质量逐渐趋于稳定,并且在树木达到一定大小时作为凋落物脱落[23],树冠部分在生物量分配中的作用较小,但排除枝叶会导致模型不准确。
3个部位中,树根生物量模型表现较差,尽管提高拟合参数的准确性非常重要,但由于树根一般在土壤深处,挖掘难度大、所需的成本较高且损失较多,采样数量往往有限,Niiyama等[24]在估算马来西亚半岛龙脑香林的根系生物量时,提出树根生物量估算的准确性和精度受到用于建立异速生长模型数据量的影响。
5 结论
不同树种各部位之间生物量比例不同,除樟子松外,其余树种的树干生物量占总生物量比例最大。随着胸径的增加,树干、树冠和树根生物量也逐渐增加。除寒温性针叶林的树干生物量最优模型是以D2H为自变量外,其余林型树干和树冠的最优模型均是以D+H为自变量的模型;落叶阔叶林和针叶林的树根生物量最优模型分别是以H、D+H作为自变量的模型。
将笔者筛选出的各部位最优模型与中国森林的模型对比发现,笔者建立的模型对生物量的估算更接近于实测值,更适合用于估算山西省森林的生物量。异速生长方程的精确性与其本身的使用范围有关,应用中应谨慎选择。
