稻蛙生态种养模式碳足迹评价
2024-01-01丁玉凌徐书含刘文超方凯凯王玲玲邹丽芳曹林奎沙之敏






摘要: 【目的】开展稻蛙生态种养模式碳足迹系统性研究,为该模式的可持续发展提供科学依据和优化建议。【方法】采用生命周期评价法,对常规水稻种植 (Traditional rice monoculture, TR ) 、绿色稻蛙种养(Green rice-frogcoculture, GF) 和有机稻蛙种养 (Organic rice-frog coculture, OF) 模式进行碳足迹评价。【结果】GF 和OF 的单位面积碳足迹分别为5 985.20 和5 632.99 kg CO2e·hm−2,相比TR 分别降低5.98% 和11.51%,单位利润碳足迹分别下降45.10% 和45.87%,单位营养密度碳足迹分别下降14.63% 和12.17%。GF 和OF 净生态系统经济效益提高了50% 以上,其中GF 的高达71.77%。与TR 相比,GF 的产量差异不显著,OF 的产量降低了16.73%;GF 和OF 的CH4 排放分别增加了58.13% 和131.18%,N2O 排放分别降低了41.06% 和63.12%;GF 和OF 具有更高的全球增温潜势和温室气体排放强度,其中,OF 的影响更为显著。GF 和OF 中,有机物料如紫云英、菜籽饼和有机肥的投入表现为净碳固定效应。碳足迹构成和敏感性分析显示, TR 的碳足迹构成中温室气体排放(57.07%) 和化肥施用(32.88%) 占比较高;OF 的碳排放和固定均高于GF,CH4 排放和尼龙防护网使用对GF 和OF 的贡献最大。不确定性分析表明,GF 和OF 的单位面积碳足迹分别为5 907.29 和5 647.25 kg CO2e·hm−2,低于TR,变异系数小于7.5%。【结论】稻蛙共作模式(GF 和OF) 相较于TR 模式具有更低的碳足迹以及更突出的碳固定效应和环境友好等特点,为气候变化背景下我国水稻产业的绿色低碳发展提供了新的科学依据。
关键词: 稻田生态系统;碳足迹;生命周期评价;绿色稻蛙;有机稻蛙
中图分类号: S966.3;S511;X82 文献标志码: A 文章编号: 1001-411X(2024)06-0939-10
气候变化引发的全球性生态环境灾难正受到越来越多关注。全球农田在支持人口爆炸性增长的同时,也排放了大量的温室气体 (Greenhouse gas,GHG)。水稻作为一种重要的粮食作物,维持了世界约一半人口的生存,同时稻田的气体排放贡献了农田温室气体排放的50% [1]。在水稻种植过程中,甲烷 (CH4) 通过淹水稻田中的厌氧发酵释放,在温室气体排放中占据主导地位,而氧化亚氮 (N2O) 则源于氮肥的施用。我国作为水稻种植和稻米消费大国,稻田排放的CH4 约为全球稻田CH4 排放的1/5[2]。未来受到气候变化和土地扩张的双重影响,预计水稻种植的温室气体排放强度将超过现有强度的2 倍[3]。因此,控制稻田温室气体排放对于减缓全球气候变化至关重要,科学评估稻田生态系统的碳排放,并开发稻田温室气体减排措施,对我国应对气候变化、实现“双碳”目标具有重要意义。
稻蛙生态种养是一种将水稻种植和蛙类养殖相结合的综合农业模式,利用稻田作为蛙的栖息地,通过蛙的活动来减少农药和化肥的施用,实现生态位互补。稻蛙生态种养模式因其突出的经济效益在我国具有一定的规模,2022 年稻蛙、稻鳖、稻鳅、稻螺4 种模式的应用面积达到8×104 hm2[4]。近年来,稻蛙生态种养模式逐渐发展出集成有机肥替代、绿肥施用和生物农药施用的有机稻蛙共作模式,在绿色生产的基础上进一步提高了环境承载力[5]。以往研究表明,稻蛙生态种养相比于单一的水稻种植具有多种优势,包括控制稻田病虫害、促进养分累积和抑制氨挥发[6-8]。Fang 等[9] 的研究表明,稻蛙种养最多可以降低水稻生长季51% 的CH4 排放,并协同促进水稻的生长。另一项研究发现,稻蛙种养可以减少1.84%~22.33% 的N2O 排放[10]。因此稻蛙生态种养具有减缓稻田温室气体排放的潜力,可以为水稻生产的环境效益和可持续性提供解决方案。
碳足迹通常指活动或产品在生命周期中直接或间接产生的温室气体的总排放量,是系统性衡量某种碳减排方案在特定应用场景下各阶段固碳减排表现的重要工具。由于农业生产过程复杂,除了作物生产过程中的直接碳排放外,农资生产及农业活动带来的能源消耗引起的间接碳排放也不容忽视,基于排放因子估算法的生命周期评估 (Lifecycle assessment, LCA) 方法被普遍应用于估算农业碳源或碳汇[11]。国际上一般采用单位面积和单位产量二氧化碳当量 (CO2e) 作为作物种植和生产的碳足迹评估单位,而在针对稻田生态种养模式的碳足迹研究中,单位利润和单位营养密度单元 (Nutrientdensity unit, NDU) 二氧化碳当量则可以被用来反映动物产品的经济属性和食物属性[12]。虽然稻田碳足迹核算已被广泛应用于评估各种水稻生态种植模式和技术,然而目前的研究仍然存在碳固存不明确和数据来源不确定性较大的问题,将农资投入中对土壤有机碳影响较大的有机物料的碳固定纳入碳足迹计算并采取敏感性分析和不确定性分析可以进一步改善评估结果的准确性[13-14]。
目前针对水稻与其他水生养殖动物共作的碳足迹研究结果都表现出较好的碳减排潜力。Ling 等[15]比较了稻虾模式和不同稻田复种模式的碳足迹,发现无论是否结合饲料投入和秸秆还田,稻虾模式的碳足迹、直接碳排放和间接碳排放均维持在较低的水平。Feng 等[16] 的研究结果显示,稻鸭共作的单位面积碳足迹相比单一水稻种植降低9%,其中对CH4 排放近22% 的抑制可能导致了较低的碳足迹。崔文超等[17] 计算了青田稻鱼共生系统的碳足迹,发现不同经营模式的单位面积碳足迹和单位产值碳足迹之间存在博弈,为稻鱼模式经济效益和环境效益的协同实现提供了理论依据。一项针对稻蛙生态种养模式的综合性分析显示,相较于双季稻和稻鸭共作,稻蛙种养承受环境压力更小,能值可持续指标表现良好,可持续发展潜力大[18]。不少研究都表明稻蛙种养可以有效抑制温室气体排放,然而现阶段仍然缺乏生命周期框架下不同稻蛙种养模式的碳足迹及其组成研究。
综上,本研究以上海青浦现代农业园区发展有限公司不同种植模式的田间试验为案例,采用LCA 方法,结合产量、利润和营养密度单元,分析不同模式的碳足迹及其组成。通过敏感性分析和不确定性分析,探究影响稻蛙种养碳足迹的关键环节,以进一步优化稻蛙生态种养模式,探索我国稻田生态系统低碳管理策略,促进农业可持续发展。
1 材料与方法
1.1 研究区概况
研究地点位于上海青浦现代农业园区 (121°01′E,30°08′N) ,气候类型为亚热带湿润型季风气候,年平均气温15.5 ℃,年平均降雨量1 056 mm。试验年份为2018 年,水稻品种为‘青角307’。试验地土壤类型为青紫泥水稻土,基本土壤理化性质如下:土壤pH 为6.53;土壤容重为1.06 g·cm−3;土壤有机碳质量分数为18.46 g·kg−1;土壤全氮、全磷和全钾质量分数分别为1.60、0.81 和11.18 g·kg−1;土壤速效氮、速效磷和速效钾质量分数分别为9 0 . 0 0 、52.60 和166.70 mg·kg−1。
1.2 试验设计
试验采用单因素随机区组设计,共设3 个处理,分别为常规水稻种植 (Traditional rice monoculture,TR)、绿色稻蛙种养(Green rice-frog coculture, GF)、有机稻蛙种养(Organic rice-frog coculture, OF) 模式,每处理均设置3 个重复。所有处理冬季种植紫云英Astragalus sinicus,夏季种植水稻Oryzasativa,不同模式的紫云英和水稻种植密度一致。水稻生长季氮肥总施用量为300 kg·hm−2,TR 处理施用尿素75 kg·hm−2 和复合肥225 kg·hm−2,GF 处理施用紫云英22.5 kg·hm−2、菜籽饼127.5 kg·hm−2、尿素75 kg·hm−2 和复合肥75 kg·hm−2,OF 处理施用紫云英22.5 kg·hm−2、菜籽饼127.5 kg·hm−2 和生物有机肥150 kg·hm−2 (均以施氮量计)。详细的试验设计和田间管理措施参见文献[19]。
供试蛙为虎纹蛙Rana rugulosa,GF 和OF 处理的蛙投放密度分别为每公顷6 000 和13 500 只,每只蛙投入时为 15~25 g。放蛙前使用尼龙网制作围网和天网,以防止鸟类捕食及蛙逃逸。在水稻分蘖期将蛙放入稻田的固定区域进行驯化,第1 周内设置饵料台给蛙喂食,适应后散养在水稻田中。GF 和OF 处理蛙生长期饲料总投入量分别为66.67和150.00 kg·hm−2。水稻收割后,GF 和OF 处理分别收获投入量一半的成品蛙。
1.3 碳足迹分析
1.3.1 系统边界
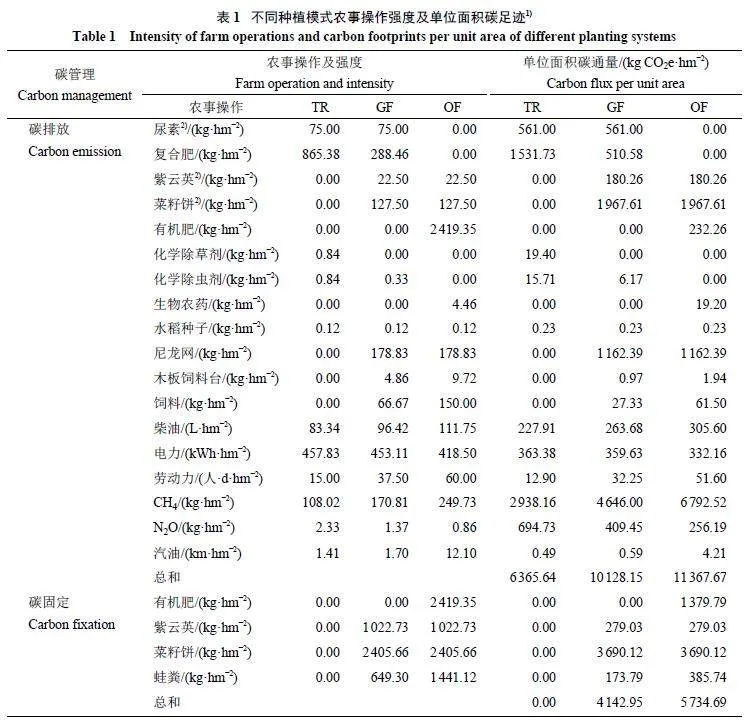
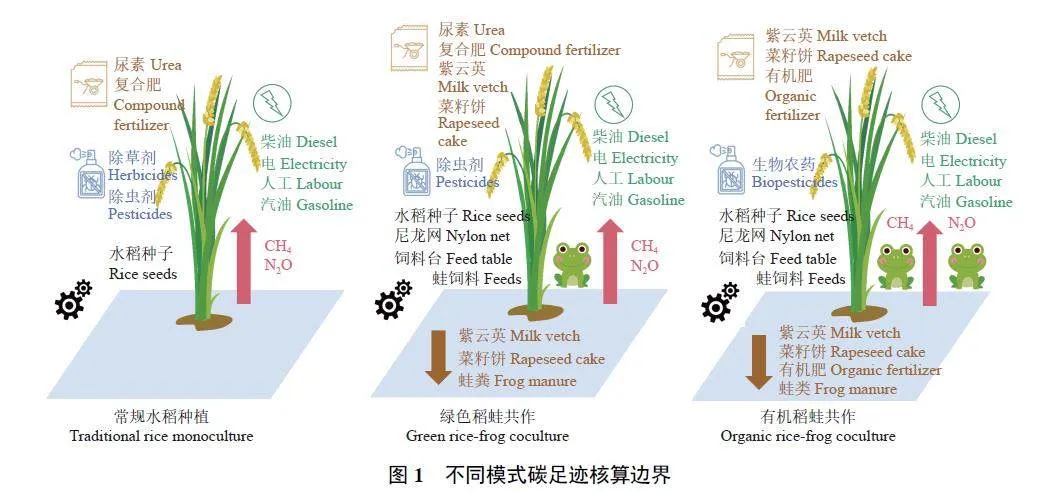
本研究基于ISO14067 框架,采用LCA 方法,参考ISO14040 生命周期评价标准,并运用排放因子计算水稻生长季生命周期中的碳排放和碳固定,单位折算为CO2e。核算边界包括种植前各投入品生产中的碳排放、种植过程中能源消耗引起的碳排放、温室气体直接排放和投入品碳固定,以及收获后的运输和加工阶段能源消耗引起的碳排放。投入品生产中的碳排放包括绿肥 (紫云英)、菜籽饼、复合肥、尿素、有机肥、化学除草剂、化学除虫剂、生物农药、水稻种子、尼龙网、蛙饲料、木板饲料台的生产过程中所产生的碳排放。种植过程中和收获后的能源消耗包括柴油使用、电力消耗、工人劳动和汽油消耗。温室气体主要包括CH4 和N2O。碳固定指的是生物质投入后当年保留在土壤中的碳,包括紫云英、菜籽饼、有机肥和蛙粪。各模式的详细碳足迹核算边界如图1 所示。
1.3.2 核算公式
本研究中全球增温潜势 (Globalwarming potential, GWP) 、温室气体排放强度(Greenhouse gas emission intensity, GHGI) 和净生态系统经济效益 (Net ecosystem economic benefits,NEEB) 的计算公式为:
GWP = 27.2ECH4 +298EN2O, (1)
GHGI = GWP=Y, (2)
NEEB = I +S-C -GWP×TaxC。(3)
式 (1) 中:ECH4为 CH4 排放量,kg·hm−2;EN2O为N2O 排放量,kg·hm−2;27.2 和298 为IPCC 第六次报告中CH4 和N2O 的GWP[20]。
式(2) 中:Y 为稻米产量,kg·hm−2。
式(3) 中:I 为收入,元;S 为补贴,元;C 为成本,元。经调查,本案例中,TR、GF 和OF 模式的收入分别为48 551.58、73 125.99 和77 261.72 元,补贴分别为17 280、20 130 和20 130 元,成本分别为41 400.0、51 412.5 和59 550.0 元。TaxC 为碳交易市场价,取值0.103 7 元·(kg CO2e)−1 [21]。
单位面积碳通量的计算公式为:
式(4) 中:CF、CE 和CS 分别为单位面积碳足迹、碳排放和碳固定,kg CO2 e·hm−2。
式(5) 中:IAi 为第i 项农事操作的强度;EFi为第i 项农事操作对应的碳排放因子。尿素、紫云英肥、菜籽饼的碳排放因子分别为7.48、8.01、15.43 kg CO2e·kg−1(基于氮含量计算),复合肥、有机肥、化学除草剂、化学除虫剂、生物农药、水稻种子、尼龙网、木板饲料台、饲料的碳排放因子分别为1.77、0.10、23.10、18.70、4.30、1.84、6.50、0.20、0.41 kg CO2 e·kg− 1 ,柴油的碳排放因子为2.74kg CO2e·L−1,电的碳排放因子为0.79 kg CO2e·kWh−1,人工的碳排放因子为0.86 kg CO2e·人−1·d−1,汽油的碳排放因子为0.35 kg CO2e·km−1 [15, 22-27]。
式(6) 中:IAj 为涉及到碳固定的第j 项农事操作的强度,FIXj 为涉及到碳固定的第j 项农事操作对应的碳固定因子。紫云英、菜籽饼、有机肥、蛙粪的碳固定因子分别为0.273、1.534、0.570 和0.268kg CO2e·kg−1 [24, 28-30]。
由于各模式利润和营养产出不一,采用多类型碳足迹指标,其计算公式为:
CFP = CF=(I +S -C), (7)
CFN = CF= (NDUr×Y +NDUf ×Sell)。(8)
式(7) 中:CFP 为单位利润碳足迹,kg CO2e·元−1。
式( 8 ) 中: C F N 为单位营养密度碳足迹,kg CO2e·NDU−1;NDUr 为稻米营养密度单元;NDUf为蛙营养密度单元;Sell 为蛙出售量,kg·hm−2。稻米和蛙营养密度的计算方法建立在蒋榕等[14] 的基础上,即食物可食部蛋白质、脂肪和膳食纤维含量除以日摄入量后除以所含热量,每100 g 稻米、蛙肉计算出的NDU 分别为0.231 4、1.028 2,数据来源为国家食物与营养咨询委员会。
1.4 敏感性分析
对参与GF 和OF 模式单位面积碳足迹计算的尿素、复合肥、紫云英、菜籽饼、有机肥、化学除草剂、化学除虫剂、生物农药、水稻种子、尼龙网、木制饲料台材料、柴油、电力、劳动力、汽油用量,CH4、N2O 排放量和蛙粪产量共18 个参数进行敏感性分析,令其在基线基础上变化±20% 和±40%,比较单位面积碳足迹结果的变化,使其变化幅度越大的参数越敏感。
1.5 不确定性分析
蒙特卡洛法是一种广泛应用于评估特定变量的不确定性影响的方法[31]。假设各农事操作强度参数符合三角分布,采用蒙特卡洛随机抽样模拟进行100 000 次迭代,计算3 种模式单位面积碳足迹的不确定性结果,形成碳足迹频数分布直方图,计算平均值、标准差和变异系数。
1.6 统计分析
利用Microsoft Excel 3.0.0 进行数据汇总和碳足迹计算,使用R4.2.1 进行不确定性分析,使用IBM SPSS Statistics 24 进行方差分析,显著性检验为T u k e y 检验,利用G r a p h P a d P r i s m 9 . 0 . 0 、R4.2.1 和Microsoft Powerpoint 3.0.0 绘图。
2 结果与分析
2.1 不同种植模式的水稻产量及温室气体排放量
TR 和GF 模式的水稻产量分别为8 827.56 和8 650.38 kg·hm−2,OF 模式的水稻产量相对较低,为7 350.69 kg·hm− 2 (图2A)。3 种模式的CH4 和N 2O 累积排放量呈相反的趋势 (图2B 和2 C ) ,GF 和OF 模式的平均CH4 排放量相比TR 模式分别显著增加58.13% 和131.18%,达到170.81 和249.73 kg·hm−2,而平均N2O 排放量相比TR 模式分别显著降低41.06% 和63.12%,达到1.37 和0.86 kg·hm−2。OF 模式的GWP 和GHGI 均最高,相比TR 模式分别增加94.03% 和133.01%,达到7 048.71 kg CO2e·hm−2 和0.96 kg CO2e·kg−1,GF 模式其次,GWP 和GHGI 相比TR 模式分别增加39.16% 和42.01%,分别达到5 055.46 kg CO2e·hm−2和0.58 kg CO2e·kg−1。
2.2 不同种植模式的稻田系统碳通量及组成
G F 和O F 模式的单位面积碳足迹分别为5 985.20 和5 632.99 kg CO2e·hm−2,相比TR 模式( 6 365.64 kg CO2 e·hm− 2 ) ,分别下降5.98% 和11.51% (图3A)。TR 模式的单位利润碳足迹为0.26kg CO2e·元−1,而GF 和OF 模式的单位利润碳足迹分别为0.14 和0.15 kg CO2e·元−1 (图3B)。TR 模式的单位营养密度碳足迹为3.12 kg CO2e·NDU−1,而GF 和OF 模式的单位营养密度碳足迹分别为2.74和2.66 kg CO2e·NDU−1 (图3C)。GF 模式的净生态系统经济预算最高,为41 319.24 元·hm−2,OF 模式紧随其后,为37 110.77 元·hm−2,TR 模式最低,为24 054.85 元·hm−2 (图3D)。
表1 显示了3 种种植模式的单位面积碳通量构成。TR 模式的总碳排放低于2 种稻蛙种养模式,其中,CH4 排放和复合肥施用的碳排放最多,分别为2 938.16 和1 531.73 kg CO2e·hm−2。GF 模式的总碳排放低于OF 模式,总碳固定相比OF 模式也较少。对于GF 模式,CH4 排放、菜籽饼施用和尼龙网使用引起的碳排放最多,分别为4 646.00、1 967.61 和1 162.39 kg CO2e·hm−2,菜籽饼施用后带来的碳固定较高,为3 690.12 kg CO2e·hm−2。对于OF 模式,甲烷排放、菜籽饼施用和尼龙网使用引起的碳排放最多,分别为6 792.52、1 967.61 和1 162.39 kg CO2 e·hm− 2 ,菜籽饼施用和有机肥施用后带来的碳固定较高,分别为3 690.12 和1 379.79 kg CO2e·hm−2。
在所有的农事操作中,2 种稻蛙模式的紫云英绿肥、菜籽饼肥和有机肥施用同时具有碳排放和碳固定作用,而这3 项农事操作的净碳足迹皆表现为碳固定。由于2 种稻蛙模式的紫云英和菜籽饼施用量相同,2 项操作分别可以带来98.77 和1 722.51 kg CO2e·hm−2 的净碳固定。有机肥施用为OF 模式贡献了1 147.53 kg CO2e·hm−2 的净碳固定。
剔除了紫云英绿肥、菜籽饼肥和有机肥施用后的碳排放构成如图4 所示。TR 模式中,对于碳排放相对贡献较大的CH4 排放和复合肥施用分别占比46.16% 和24.06%,N2O 排放和尿素施用紧随其后,分别占比10.91% 和8.81%,能源消耗也具有一定的贡献,电力消耗占比5 . 7 1 % ,柴油消耗占比3.58%。GF 模式中,对于碳排放贡献最大的分别为CH4 排放和尼龙网使用,占比58.22% 和14.57%,尿素施用、复合肥施用和N2O 排放其次,分别占比7.03%、6.40% 和5.13%,能源消耗也具有一定的贡献,电力消耗占比4.51%,柴油消耗占比3.30%。OF 模式中,对于碳排放贡献最大的分别为CH4 排放和尼龙网使用,占比75.58% 和12.93%,电力消耗、柴油消耗和N2O 排放其次,分别占比为3.70%、3.40% 和2.85%。
2.3 不同种植模式碳足迹的敏感性分析
不同稻蛙种养模式单位面积碳足迹的敏感性分析结果如图5 所示。在18 个输入参数中,CH4 排放最为敏感。当CH4 排放量在±40% 范围内变动时,GF 模式的单位面积碳足迹的变化范围为4 126.80~7 843.60 kg CO2e·hm−2,OF 模式的变化范围更高,达到2 915.98~8 350.00 kg CO2e·hm−2。在GF 模式中,菜籽饼肥施用量和尼龙网使用量也较为敏感,当这2 个参数在±40% 范围内变动时,单位面积碳足迹的变化范围分别为5 296.19~6 674.21 和5 520.24~6 450.15 kg CO2e·hm−2。在OF 模式中,菜籽饼肥施用量、尼龙网使用量和有机肥施用量较为敏感,当这3 个参数在±40% 范围内变动时,单位面积碳足迹的变化范围分别为4 943.98~6 321.99、5 168.03~6 097.94和5 173.97~6 092.00 kg CO2e·hm−2。其余参数对于2种稻蛙种养模式的碳足迹核算结果的影响均较小。
2.4 不同种植模式碳足迹的不确定性分析
不确定性分析的结果如图6 所示。TR 模式的单位面积碳足迹最高,为6 311.48 kg CO2e·hm−2,GF 模式其次,为5 907.29 kg CO2e·hm−2,OF 模式最低,为5 647.25 kg CO2e·hm−2。TR、GF 和OF 模式的变异系数分别为5.73%、7.16% 和7.26%,均小于10%,表明本案例中3 种模式的单位面积碳足迹结果的不确定性较低。
3 讨论与结论
本研究发现,OF 模式的产量相比TR 模式显著下降16.73%,这可能是由于绿肥和有机肥的输入相比化肥的养分释放更慢,且不利于水稻偏好的铵态氮累积,OF 模式中更大量的蛙带来了更多动物源有机质的输入,考虑到研究区域的土壤为青紫泥,基底有机质为31.83 g·kg−1,较为肥沃,对于有机质输入的反应较为迟钝;也可能是有机肥相比化肥,在水稻分蘖期氮素供应不足,导致有效分蘖数减少,这一系列因素可能导致对产量的负面影响[32]。
Fang 等[9] 的研究发现,在水稻生长季节,稻蛙种养可以使CH4 排放量减少21.68%~51.21%。而本研究发现,GF 和OF 模式分别使得CH4 排放量增加58.13% 和131.18%。Yuan 等[33] 对比了化肥施用、有机肥部分替代和有机肥完全替代化肥施用对稻田CH4 排放的影响,发现有机肥完全替代处理的CH4 排放量最高,可能源自有机肥对土壤碳底物有效性的提高。虽然蛙游、爬、跳等活动可能会通过增强稻田水体扰动、提高稻田水体溶解氧浓度而促进CH4 氧化[34],但是蛙通过排泄向稻田中输入的新鲜有机质对CH4 产生也有一定的贡献[35]。结合2种稻蛙种养模式中不同量级的有机肥输入,可能是导致1.5~2.0 倍CH4 增排的原因。本研究还发现,GF和OF 模式分别显著抑制了41.06% 和63.12% 的N2O 排放,这与一项关注稻田生态种养的meta 研究中稻蛙种养可以显著降低67% 的N2O 排放的结论[36] 相一致。值得注意的是,有机肥施用可能也有利于N2O 排放的减少,如Kong 等[37] 研究发现,有机肥替代通过抑制硝化过程中的氨氧化活性和反硝化过程中的亚硝酸盐还原活性,加强稻田反硝化过程中N2O 的还原。虽然化学农药对于N2O 排放的影响还不甚清晰,Das 等[38] 的研究提示,在有利于反硝化的潮湿田地中,部分化学农药的施用 (如异硫氰酸甲酯) 有可能会贡献N2O 排放。由于稻田温室气体排放中CH4 占据了主要地位,本研究中2 种稻蛙种养模式的GWP 都高于TR 模式,同时GHGI 都高于TR 模式,不过GF 模式相对具有更低的GWP 和GHGI 增长。
在过去的30 年中,随着物资投入和能源消耗的增加,中国稻米生产的碳足迹大幅增加,增长幅度位列所有类型食物中的第1 位[39],亟需发展水稻低碳生产技术。本研究发现,在稻田中投入蛙并集成绿肥还田、有机肥施用、生物农药替代等生态技术可以降低5.98%~11.51% 的单位面积碳足迹,蛙的出售可以带来一定的经济收入,应用稻蛙生态种养模式可以降低45.10%~42.87% 的单位利润碳足迹,即使具有GWP 增长的风险,稻蛙生态种养对NEEB 的促进作用仍然超过了50%。研究表明,稻蛙模式的经济效益在各复合种养模式中位列第2,如果将蛙皮、蛙分泌物进行二次开发利用,可以带来更多经济效益[40]。OF 模式的NEEB 低于GF 模式,可能是因为其GWP 相对较高,另外本研究未考虑有机稻蛙种养可能对稻米品质的正面影响以及有机蛙稻米的溢价效应,因此OF 模式的经济效益还有提升的空间。由于蛙肉是优质蛋白质来源,考虑到稻蛙生态种养模式的营养效益,本研究发现GF 和OF 模式的单位营养密度产出碳足迹相比TR模式降低12.17%~14.63%,相比文献中稻虾共作模式也更低[14],这可能是因为蛙肉的综合营养含量略高于小龙虾。总之,不同维度的碳足迹指标的结果表明,稻蛙生态种养模式在可以实现固碳减排的基础上还有着良好的经济效益和食品营养产出功能。
从碳足迹构成来看,本研究发现,2 种稻蛙生态种养模式的碳排放都高于TR 模式,但约占碳排放量一半的碳固定使得2 种模式的净碳排放较低,尤其是OF 模式。本研究的碳足迹核算边界内考虑了有机物料的投入带来的碳固定,这在其他计算农业碳足迹的文献中是有例可循的[41],具体表现在紫云英还田、菜籽饼施用、有机肥施用和蛙粪输入后对土壤有机碳的增加,虽然紫云英、菜籽饼和有机肥的生产过程产生了间接碳排放,但与它们的碳固定量相互抵消后,表现出净碳固定的效应,这些生态技术的固碳作用对于稻田生态种养模式碳足迹的降低具有较大贡献。需要注意的是,本研究中碳固定的估算基于有机物料的碳含量,未考虑到降解速率,因此存在一定程度上的高估,然而已有的不少证据都表明绿肥和有机肥的输入对稻田土壤中碳固存的潜力巨大[42-43]。就碳排放构成而言,水稻种植期间的温室气体排放,尤其是CH4 排放,无论在TR 模式还是在稻蛙生态种养模式中都占主导地位(gt;45%)。TR 模式中另一大碳排放较高的项目是化肥的施用,这符合曹黎明等[44] 关于上海市水稻生产碳足迹的研究结果,可能与我国化肥的平均温室气体排放系数较高有关[45]。在2 种稻蛙生态种养模式中,除了CH4 排放和菜籽饼施用的间接碳排放较高(这部分被碳固定所抵消) 外,蛙配套设施的建造也带来了一定的碳排放,例如为了防备蛙被鸟类或黄鼠狼吃掉所搭建的尼龙材质围网和天网,形成的间接碳排放在2 种模式的碳足迹中占比均超过10%,这可能与尼龙这种化工产品的排放系数较高有关,另一方面可能是因为对网的网眼大小和分布面积有较高的需求,会导致材料用量较大。
综上所述,稻蛙生态种养增加了CH4 排放,但对N2O 排放具有抑制效果。GF 和OF 模式的单位面积、单位利润和单位营养密度碳足迹相比常规水稻种植分别降低了超过5%、40% 和12%,且NEEB较高,这表明稻蛙生态种养模式具有显著的环境、经济和营养效益。稻蛙生态种养模式中有机物料的施用带来了显著的碳固定,约占碳排放量的40.91%~50.45%,因此其碳足迹相比常规模式更低。相比GF 模式,OF 模式的CH4 排放增加了46.20%,但有机肥施用的碳固定使其单位面积碳足迹降低5.88%。由于CH4 排放仍然是碳足迹中重要的组成成分 (48.22%~75.58%),蛙管理过程中如尼龙网使用造成了较多的间接碳排放(12.93%~14.57%),未来稻蛙生态种养模式需要引入更多的CH4 减排措施和更为绿色环保的蛙管理措施,以降低碳排风险。
参考文献:
[1]QIAN H, ZHU X, HUANG S, et al. Greenhouse gas emissions and mitigation in rice agriculture[J]. NatureReviews Earth amp; Environment, 2023, 4(10): 716-732.
[2]唐志伟, 张俊, 邓艾兴, 等. 我国稻田甲烷排放的时空特征与减排途径[J]. 中国生态农业学报(中英文), 2022,30(4): 582-591.
[3]VAN GROENIGEN K, VAN KESSEL C, HUNGATEB. Increased greenhouse-gas intensity of rice productionunder future atmospheric conditions[J]. Nature ClimateChange, 2013, 3: 288-291.
[4]于秀娟, 郝向举, 党子乔, 等. 中国稻渔综合种养产业发展报告(2023)[J]. 中国水产, 2023, 573(8): 19-26.
[5]鲁艳红, 廖育林, 聂军, 等. 紫云英利用下有机稻—蛙生产模式及其效应比较[J]. 湖南农业科学, 2017(8): 11-14.
[6]谢洪科, 邹朝晖, 刘功朋, 等. 不同蛙类及其密度对水稻性状和主要害虫的影响[J]. 江西农业学报, 2014, 26(6):21-25.
[7]周雪芳, 朱晓伟, 陈泽恺, 等. 稻蛙生态种养对土壤微生物及无机磷含量的影响[J]. 核农学报, 2016, 30(5): 971-977.
[8]陈慧妍, 沙之敏, 吴富钧, 等. 稻蛙共作对水稻−紫云英轮作系统氨挥发的影响[J]. 中国生态农业学报(中英文), 2021, 29(5): 792-801.
[9]FANG K, DAI W, CHEN H, et al. The effect of integratedrice-frog ecosystem on rice morphological traits andmethane emission from paddy fields[J]. Science of theTotal Environment, 2021, 783: 147123.
[10]FANG K, GAO H, SHA Z, et al. Mitigating globalwarming potential with increase net ecosystem economicbudget by integrated rice-frog farming in easternChina[J]. Agriculture, Ecosystems amp;Environment, 2021,308: 107235.
[11]罗怀良. 国内农业碳源/汇效应研究: 视角、进展与改进[J]. 生态学报, 2022, 42(9): 3832-3841.
[12]夏龙龙, 颜晓元. 中国粮食作物生命周期生产过程温室气体排放的研究进展及展望[J]. 农业环境科学学报,2020, 39(4): 665-672.
[13]HOLKA M, KOWALSKA J, JAKUBOWSKA M. Reducingcarbon footprint of agriculture: Can organic farminghelp to mitigate climate change?[J]. Agriculture, 2022,12(9): 1383.
[14]蒋榕, 徐强, 李京咏, 等. 稻虾共作模式碳足迹评价的敏感性和不确定性分析[J]. 中国生态农业学报(中英文),2022, 30(10): 1577-1587.
[15]LING L, SHUAI Y J, XU Y, et al. Comparing rice productionsystems in China: Economic output and carbonfootprint[J]. Science of the Total Environment, 2021,791: 147890.
[16]FENG L, WANG R, WANG R, et al. Life cycle assessmentof rice-duck co-culture systems[J]. EcosystemHealth and Sustainability, 2024, 10: 0166.
[17]崔文超, 焦雯珺, 闵庆文. 不同土地经营模式的稻鱼共生系统环境影响评价[J]. 中国生态农业学报(中英文),2022, 30(4): 630-640.
[18] 钟颖, 沙之敏, 顾麦云, 等. 基于能值分析的稻蛙生态种养模式效益评价[J]. 中国生态农业学报(中英文),2021, 29(3): 572-580.
[19]FANG K, YI X, DAI W, et al. Effects of integrated ricefrogfarming on paddy field greenhouse gas emissions[J].International Journal of Environmental Research andPublic Health, 2019, 16(11): 1930.
[20]MASSON-DELMOTTE V, ZHAI P, PIRANI A, et al.Climate change 2021: The physical science basis. Contributionof working group I to the sixth assessment reportof the intergovernmental panel on climate change[R].Cambridge: Cambridge University Press, 2021.
[21]LI B, FAN C H, ZHANG H, et al. Combined effects ofnitrogen fertilization and biochar on the net global warmingpotential, greenhouse gas intensity and net ecosystemeconomic budget in intensive vegetable agriculture insoutheastern China[J]. Atmospheric Environment, 2015,100: 10-19.
[22]CHEN P, YANG J, JIANG Z, et al. Prediction of futurecarbon footprint and ecosystem service value of carbonsequestration response to nitrogen fertilizer rates in riceproduction[J]. Science of the Total Environment, 2020,735: 139506.
[23]GONG H, LI J, SUN M, et al. Lowering carbon footprintof wheat-maize cropping system in North China Plain:Through microbial fertilizer application with adaptivetillage[J]. Journal of Cleaner Production, 2020, 268:122255.
[24]ABDULKAREEM R. 绿肥还田对双季稻系统温室气体排放及其相关土壤微生物的影响[D]. 北京: 中国农业科学院, 2020.
[25]MANTOAM E, ANGNES G, MEKONNEN M, et al. Energy,carbon and water footprints on agricultural machinery[J]. Biosystems Engineering, 2020, 198: 304-322.
[26]U. S. Environmental Protection Agency. Inventory of U.S. greenhouse gas emissions and sinks: 1990-2018[R/OL]. [2024-06-01]. https://www.epa.gov/ghgemissions/inventory-us-greenhouse-gas-emissions-and-sinks-1990-2018.
[27]中华人民共和国生态环境部. 2018 年度减排项目中国区域电网基准线排放因子[R/OL]. (2020-12-29)[2024-06-01]. https://www.mee.gov.cn/ywgz/ydqhbh/wsqtkz/202012/t20201229_815384.shtml.
[28]熊荣波, 孟艳, 李屹, 等. 高含固率菜籽饼与牛粪−羊粪混合厌氧发酵产沼气特性[J]. 环境工程学报, 2021,15(7): 2427-2435.
[29]YUAN J, SHA Z, HASSANI D, et al. Assessing environmentalimpacts of organic and inorganic fertilizer ondaily and seasonal greenhouse gases effluxes in ricefield[J]. Atmospheric Environment, 2017, 155: 119-128.
[30]陈慧妍. 稻蛙种养模式的氨挥发特征[D]. 上海: 上海交通大学, 2021.
[31]TU Q, MCDONNELL B. Monte Carlo analysis of lifecycle energy consumption and greenhouse gas (GHG)emission for biodiesel production from trap grease[J].Journal of Cleaner Production, 2016, 112: 2674-2683.
[32]任科宇, 徐明岗, 张露, 等. 我国不同区域粮食作物产量对有机肥施用的响应差异[J]. 农业资源与环境学报,2021, 38(1): 143-150.
[33]YUAN J, YUAN Y, ZHU Y, et al. Effects of differentfertilizers on methane emissions and methanogenic communitystructures in paddy rhizosphere soil[J]. Science ofthe Total Environment, 2018, 627: 770-781.
[34]方凯凯. 稻蛙种养对稻田甲烷排放的影响及其机理研究[D]. 上海: 上海交通大学, 2021.
[35]XIA L, LAM S, YAN X, et al. How does recycling oflivestock manure in agroecosystems affect crop productivity,reactive nitrogen losses, and soil carbon balance?[J]. Environmental Science amp; Technology, 2017,51(13): 7450-7457.
[36]WANG C, SHI X, QI Z, et al. How does rice-animal coculturesystem affect rice yield and greenhouse gas? Ameta-analysis[J]. Plant and Soil, 2023, 493(1): 325-340.
[37]KONG D, ZHANG X, YU Q, et al. Mitigation of N2Oemissions in water-saving paddy fields: Evaluating organicfertilizer substitution and microbial mechanisms[J].Journal of Integrative Agriculture, 2024, 23(9): 3159-3173.
[38]DAS S, WANG W, REEVES S, et al. Non-target impactsof pesticides on soil N transformations, abundancesof nitrifying and denitrifying genes, and nitrous oxideemissions[J]. Science of the Total Environment, 2022,844: 157043.
[39]LIN J, HU Y, CUI S, et al. Carbon footprints of food productionin China (1979 —2009)[J]. Journal of CleanerProduction, 2015, 90: 97-103.
[40]李冬雪, 宋星陈, 熊玉唐, 等. 西南山区稻蛙复合种养的研究与实践[J]. 农学学报, 2018, 8(12): 6-12.
[41]HORRILLO A, GASPAR P, ESCRIBANO M. Organicfarming as a strategy to reduce carbon footprint in dehesaagroecosystems: A case study comparing differentlivestock products[J]. Animals, 2020, 10(1): 162.
[42]TIAN K, ZHAO Y, XU X, et al. Effects of long-term fertilizationand residue management on soil organic carbonchanges in paddy soils of China: A meta-analysis[J]. Agriculture,Ecosystems amp; Environment, 2015, 204: 40-50.
[43]KAMRAN M, HUANG L, NIE J, et al. Effect of reducedmineral fertilization (NPK) combined with greenmanure on aggregate stability and soil organic carbonfractions in a fluvo-aquic paddy soil[J]. Soil and TillageResearch, 2021, 211: 105005.
[44]曹黎明, 李茂柏, 王新其, 等. 基于生命周期评价的上海市水稻生产的碳足迹[J]. 生态学报, 2014, 34(2): 491-499.
[45]陈舜, 逯非, 王效科. 中国氮磷钾肥制造温室气体排放系数的估算[J]. 生态学报, 2015, 35(19): 6371-6383.
【责任编辑 庄 延】
基金项目:上海市水稻产业体系项目(沪农科产字(2022) 第3 号)
