以组合生物合成策略为导向的C-核苷类多氧霉素的研究
2023-12-30刘元园李天竹陈文青
刘元园,李天竹,熊 莉,龚 蓉,陈文青
武汉大学药学院,组合生物合成与新药发现教育部重点实验室,湖北 武汉 430071
核苷类抗生素(Nucleoside antibiotics)是一类重要的微生物次级代谢产物,具有抗细菌、抗真菌、抗病毒和除草等多种功效,故在医药、化工以及农业等领域被广泛应用[1-2];根据糖苷键的差异,可将其分为N-核苷和C-核苷类抗生素[3]。
多氧霉素(polyoxin)由可可链霉菌阿苏变种(Streptomyces cacaoi var. asoeinsis)和金产色链霉菌(Streptomyces aureochromogenes)产生[4],由核苷骨架(nucleoside skeleton)、 氨甲酰多聚草氨酸(carbamoylpolyoxamic acid,CPOAA) 及聚肟酸(polyoximic acid,POIA)构成(图1)。由于多氧霉素与UDP-N-乙酰葡萄糖胺的结构类似,故为几丁质合成酶的竞争性抑制剂,可阻断真菌细胞壁的生物合成。因此,对多种植物真菌病害具有良好的防治效果[5]。马来亚霉素(malayamycin)为重要的C-核苷类抗生素,由马来西亚链霉菌DSM 14072(Streptomyces malaysiensis DSM 14072)产生,因其具有独特的全氢呋喃并吡喃双环C-核苷结构(图1),且能有效抑制小麦颖枯病和白粉病等植物病原真菌而被广泛关注[6-8]。近期有研究鉴定了S.malaysiensis DSM 14072 中马来亚霉素的生物合成基因簇,并初步揭示了其生物合成机制[9]。
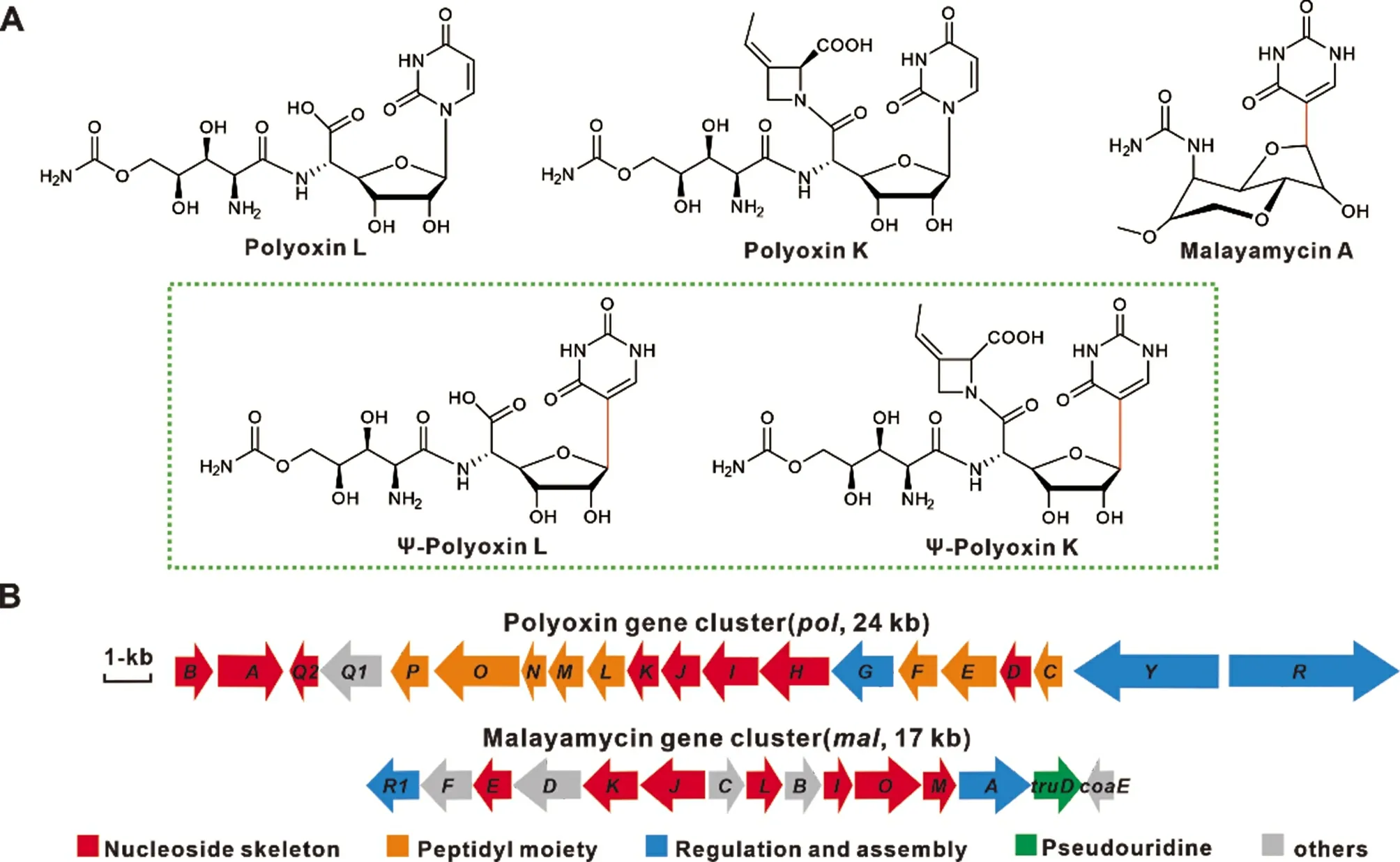
图1 多氧霉素和马来亚霉素的化学结构式及生物合成基因簇
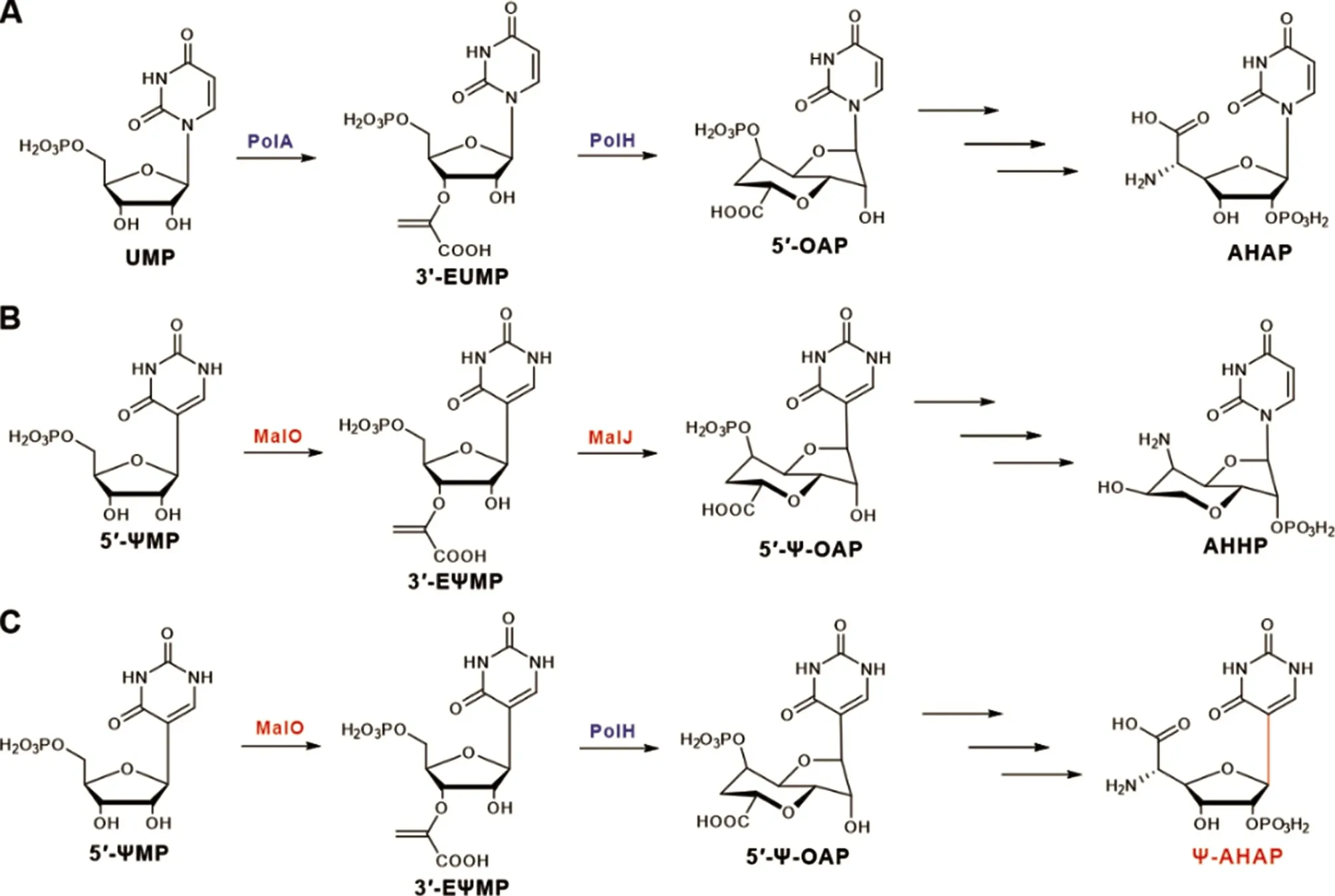
图2 多氧霉素、马来亚霉素和杂合抗生素的核苷骨架生物合成途径
多氧霉素具有显著的抗真菌活性,但其结构中N-糖苷键的稳定性相对较差,会影响其在农业中的应用。本研究采用合成策略将马来亚霉素基因簇中负责C-核苷的truD 和malO 基因整合到多氧霉素相关的突变株中,设计合成了新型C-核苷类多氧霉素Ψ-polyoxin L 和Ψ-polyoxin K,为今后基于二者的结构设计合成多重杂合抗生素奠定了基础。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒
E.coli DH10B、E.coli ET12567(pUZ8002)、E.coli BW25113(pIJ790)、S.aureochromogenes YB172、SA1、SA2、S. malaysiensis DSM 14072、长柄链格孢菌(Alternaria alternate)和皮状丝孢酵母(Trichosporon cutaneum 3071)均保存于本实验室。实验所用质粒及其特征见表1 所列。

表1 本研究所用质粒
1.1.2 工具酶、试剂及药品
限制性内切酶及T4 DNA 连接酶购自New England Biolabs 公司;高保真PrimeSTAR® Max DNA 聚合酶购自Takara 公司;rTaq DNA 聚合酶和DNA Marker 购自鼎国生物;多片段一步法快速克隆试剂盒购自翌圣生物;质粒提取和DNA 胶回收试剂盒购自Omega 公司;甲酸(质谱级)和甲醇(质谱级)购自上海安谱;相关培养基购自OXOID 公司;其他试剂(分析纯)均购买国药。
1.1.3 引物
本研究中所用引物如表2 所列。

表2 本研究所用引物
1.2 常规方法
大肠杆菌与链霉菌的遗传操作方法分别参考了相关文献[13],常规分子的克隆操作均按照说明书进行。
1.3 链霉菌菌株的发酵、发酵样品的生物测定以及LC-MS 检测
链霉菌的摇瓶发酵方法参考相关文献[14],相关发酵样品的处理与生物测定按照参考文献进行[12]。
1.4 利用PCR-targeting 技术同框缺失pJTU4774中polA 基因
将pJTU4774(含有阿伯拉霉素抗性)转至E.coli BW25113(pIJ790)后于30 ℃条件下进行培养,后续具体实验操作参考相关文献[12]。
2 结果与分析
2.1 新型C-核苷类多氧霉素的生物合成途径设计
最近研究表明,马来亚霉素基因簇中有多氧霉素核苷骨架生物合成所需的所有相关同源基因,并与之一一对应。在多氧霉素的合成过程中,以UMP为起始底物,形成含有尿苷的生物合成过程中间体;而马来亚霉素的生物合成中,以ΨMP 为起始底物,形成含假尿苷的类似中间体。磷酸烯醇式丙酮酰转移酶PolA 催化UMP 形成3'-EUMP,而MalO 则只能接受ΨMP 为底物形成3'-EΨMP,以确保马来亚霉素中假尿苷结构的特殊性。随后,在S-腺苷甲硫氨酸自由基蛋白PolH(MalJ)、磷酸激酶PolQ2(MalE)、磷酸水解酶PolJ(MalL)、α-酮戊二酸/铁依赖加氧酶PolK(MalM)和转氨酶PolI(MalK)的催化下,形成AHOAP(5'-amino-6'-hydroxyoctosyl acid 2'-phosphate)。在多氧霉素中,α-酮戊二酸/铁依赖加氧酶PolD 通过催化C-C 键断裂将AHOAP 转化为AHAP(Aminohexuronic acid 2'-phosphate)。而在马来亚霉素生物的合成途径中,AHOAP 在α-酮戊二酸/铁依赖加氧酶MalI 和脱氢酶MalB 的共同催化下形成AHHP(5'-amino-6'-hydroxylheptosyl 2'-phosphate)[15-17]。
考虑到多氧霉素和马来亚霉素核苷骨架生物合成过程的相似性,以及多氧霉素中各结构单元的生物合成相对独立,本研究计划中断相关模块的生物合成过程。在此过程中,将保留多氧霉素和马来亚霉素各自生物合成杂合抗生素所需的模块,并将这些相关模块组装形成目标杂合抗生素。
2.2 定向生物合成Ψ-polyoxin L
在前期研究中, 本实验室构建了S.aureochromogenes YB172 中polF 基因(推测与POIA生物合成有关)同框缺失和polA 基因被tsr 抗性基因中断的突变菌株SA2,借助SA2 进行Ψ-polyoxin L 的相关研究。首先将truD 克隆至整合型载体pIB139 获得质粒pIB139/truD。此外,将malO 基因克隆至pJTU968 的NdeI 和EcoRI 酶切位点。然后用MfeI 和EcoRI 酶切获得带有红霉素抗性基因启动子ermEp*的malO 片段,并将含有ermEp*启动子的malO 片段克隆至pIB139/truD 的EcoRI 酶切位点上,验证正确后获得质粒pIB139/truD&malO。
将构建的质粒pIB139/truD&malO 通过属间接合转移导入突变菌株SA2 中,筛选得到重组菌株SA2::pIB139/truD&malO,命名为PA1。将PA1 和负对照SA2::pIB139 进行发酵,发酵完毕后经草酸调节pH 至3-4,并以A. alternate 为指示菌测定生物活性,结果如图3A 所示:与SA2::pIB139 相比,PA1恢复了微弱的抗真菌活性,表明PA1 菌株可能产生了具有抗真菌活性的新化合物。随后进行了LCHRMS 检测分析,相较于SA2::pIB139,重组菌PA1的发酵液中能检测到质荷比([M+H]+)为478.139 9的化合物,与Ψ-polyoxin L 的理论值478.141 6 基本一致;其二级断裂碎片为460.187 1、417.216 9、399.148 7 和288.094 3 等,与Ψ-polyoxin L 的理论断裂碎片基本一致。实验结果表明,本研究成功产生了具有抗真菌活性的新型杂合抗生素Ψ-polyoxin L。
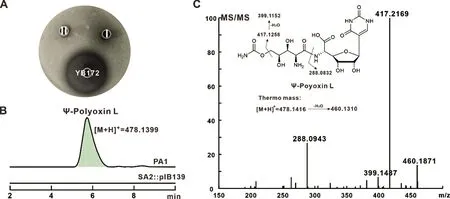
图3 重组菌株PA1 代谢产物的生物活性测定与LC-HRMS 检测分析
2.3 定向生物合成Ψ-polyoxin K
有研究表明,含有完整结构单元的多氧霉素组分相较于不含POIA 结构单元的组分对T. cutaneum等真菌的抑制活性更强[18]。因此,本研究拟将马来亚霉素中的C-核苷引入含有完整结构单元的多氧霉素组分中以获得杂合抗生素Ψ-polyoxin K。前期实验室保藏了S. aureochromogenes YB172 中polA 基因被tsr 中断的突变菌株SA1,因此将上述构建的质粒pIB139/truD&malO 通过属间接合转移SA1 中筛选得到重组菌株SA1::pIB139/truD&malO,命名为PA2。将PA2 和负对照SA1::pIB139 进行发酵,发酵完毕后经草酸调节pH 至3-4,以T. cutaneum 为指示菌进行生物活性测定,结果如图4A 所示:相较于SA1::pIB139,PA2 恢复了较强的抗真菌活性。LCHRMS 检测分析显示:相较于SA1::pIB139,重组菌PA2 的发酵液中不仅能够检测到Ψ-polyoxin L,还能够检测到质荷比([M+H]+)为587.1922 的化合物,其二级断裂碎片为559.170 3、526.317 9、460.130 2、442.002 4、397.0745 和288.095 7 等,与Ψ-polyoxin K 的理论断裂碎片基本一致。实验结果表明重组菌株PA2 定向产生了Ψ-polyoxin L 和Ψ-polyoxin K,并且Ψ-polyoxin K 显示出了较强的抗真菌活性。
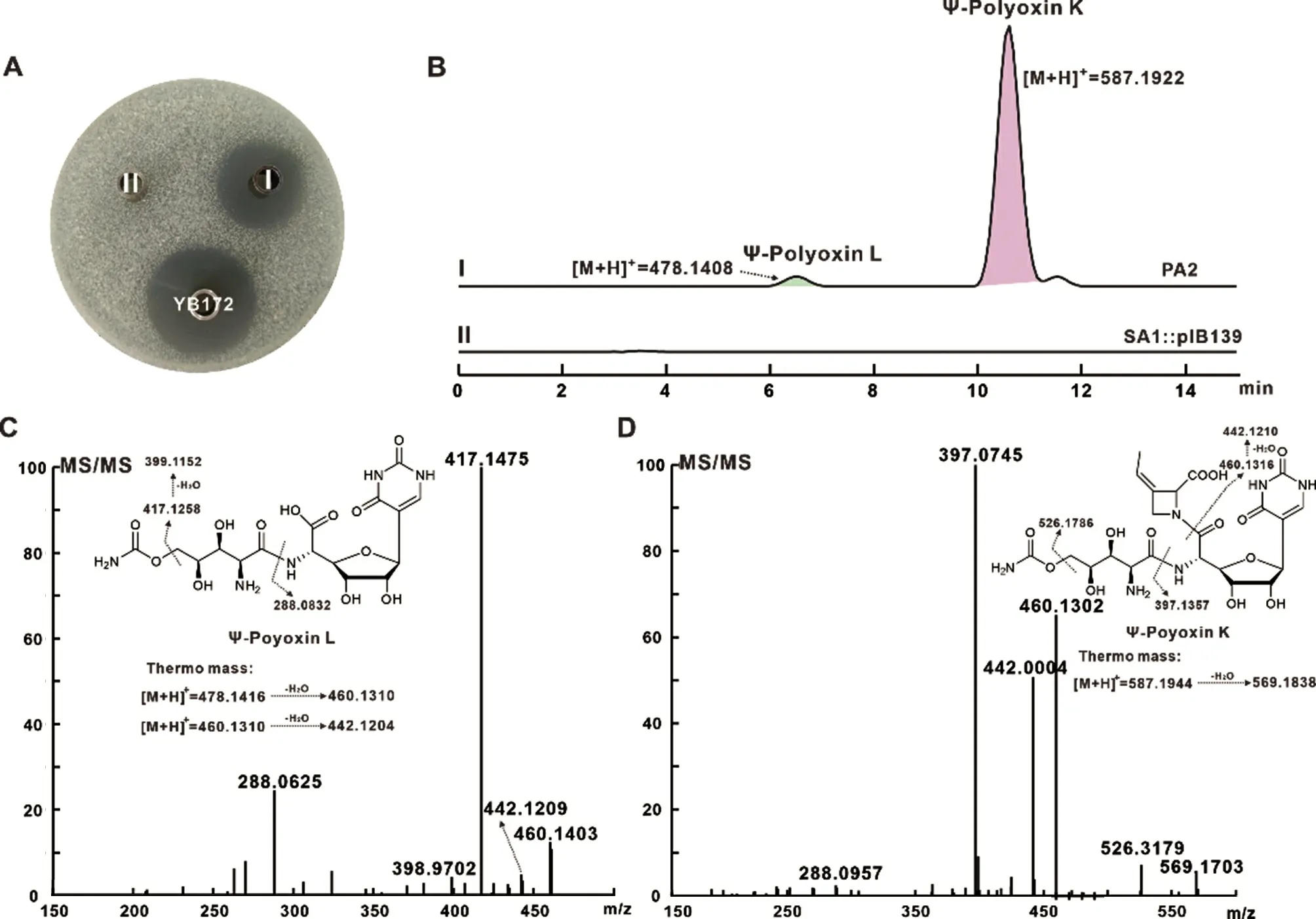
图4 重组菌PA2 代谢产物的生物活性测定与LC-HRMS 分析
2.4 在重组菌株PA2 中加倍多氧霉素基因簇提高杂合抗生素的产量
综合生物活性测定和高分辨质谱检测等手段的应用,确定了重组菌株PA1 能够产生Ψ-polyoxin L,重组菌株PA2 不仅能够产生Ψ-polyoxin K,还能产生少量的Ψ-polyoxin L,但二者产量都较低,限制了目标杂合抗生素的开发与应用。因此,本研究拟将多氧霉素相关结构单元生物合成途径的基因簇加倍,以期提高目标杂合抗生素的产量。在含有多氧霉素完整生物合成基因簇的质粒pJTU4774 的基础上,采用PCR-targeting 技术将polA 基因进行同框缺失得到质粒pJTU4774ΔpolA(图5A)。以polAidF/R 为鉴定引物进行PCR 验证,结果如图5B 所示:pJTU4774 的PCR 产物大小为1.44 kb,连上卡那霉素抗性基因neo 后的质粒PCR 产物大小为2.06 kb,而pJTU4774ΔpolA 的PCR 产物大小为0.62 kb,随后将PCR 验证正确的质粒pJTU4774ΔpolA 进行测序验证(图5C)。
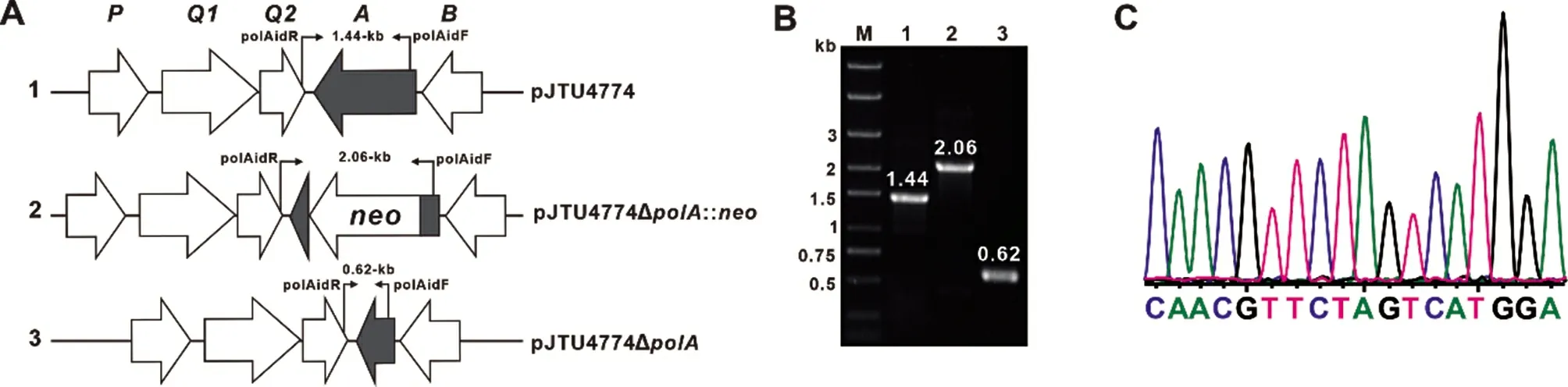
图5 质粒pJTU4774ΔpolA 的构建及验证
由于pIB139 与pJTU4774 具有相同的抗性,无法同时通过属间接合转移的方法整合到底盘菌染色体上。因此,本研究采用了分别带有ermEp*启动子的truD 和malO 基因,通过Gibson 重组的方式克隆至pJTU4774ΔpolA 的EcoRI 位点上,得到质粒pJTU4774ΔpolA/truD&malO。随后,通过属间接合转移的方法将其导入SA1 中,筛选得到重组菌株SA1::pJTU4774ΔpolA/truD&malO,命名为PA3(图6A)。将重组菌株PA3 和PA2 同批次发酵,取等量原始发酵液以T. cutaneum 为指示菌对其进行生物活性测定,检测结果如图6B 显示:重组菌株PA3 相较PA2 对T. cutaneum 的抑制作用更强,表明PA3发酵液中相应目标杂合抗生素的产量相较PA2 有了明显的提高。进一步的LC-HRMS 分析如图6C-D所示:PA3 中Ψ-polyoxin L 的产量提高到了原来的150%左右,而Ψ-polyoxin K 的相对产量较PA2 提高到了原来的330%左右。实验结果表明,通过加倍多氧霉素生物合成基因簇pJTU4774ΔpolA,能显著提高目标化合物Ψ-polyoxin L 和Ψ-polyoxin K 的产量。
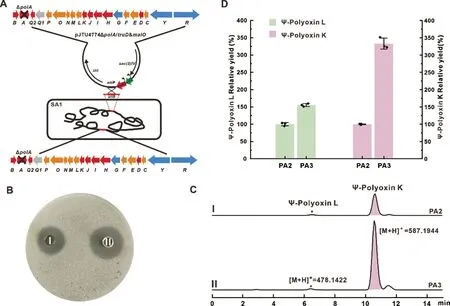
图6 多氧霉素基因簇加倍重组菌株PA3 构建示意图及相关杂合抗生素产量比较
3 讨论与结论
近年来,面对从天然产物中分离新抗生素越来越缓慢和抗生素耐药性增长等困境,在全面了解天然抗生素生物合成代谢途径的基础上,通过组合生物合成策略对其进行结构改造成为了破解抗生素开发困境的重要途径之一[19]。本研究中,依循组合生物合成策略的指导, 以多氧霉素工业菌S.aureochromogenes YB172 为底盘菌株,通过改造其生物合成途径并整合马来亚霉素中负责C-核苷结构的基因truD 和malO,首次定向产生了具有C-核苷结构的新型杂合抗生素Ψ-polyoxin L 和Ψpolyoxin K。此外,通过增加多氧霉素生物合成基因簇的拷贝数,显著提高了杂合抗生素Ψ-polyoxin L和Ψ-polyoxin K 的产量。这为今后基于多氧霉素和马来亚霉素等相关核苷类抗生素设计和合成多重杂合抗生素奠定了基础。
