叉角厉蝽对于不同虫龄茶谷蛾幼虫的捕食功能反应
2023-12-25龚雪娜罗梓文玉香甩王雪松龙丽雪梁晓静陈林波龙亚芹
龚雪娜,罗梓文,玉香甩,曲 浩,王雪松,龙丽雪,张 东,梁晓静,陈林波*,龙亚芹*
(1.云南省农业科学院茶叶研究所/云南省茶树种质资源创新与配套栽培技术工程研究中心/云南省茶学重点实验室,昆明 650201;2.北京云普茶业有限公司,北京 100025)
茶谷蛾AgriophararhombataMeyr.,又称茶灰木蛾,属鳞翅目Lepidoptera 谷蛾科Tineidae,是茶树重要的食叶性害虫之一,尤以幼虫咀食叶肉为害最为严重[1]。该虫喜食成叶和老叶,大发生时亦为害嫩叶,受害轻的植株叶片形成缺刻、干枯,受害重的茶树叶片被吃光,致使植株似火烧状,造成无茶可采,严重时整株枯死[2]。该虫曾是东北印度的主要害虫,后在广东、福建、厦门、四川等地普遍暴发,在浙江一年发生7 代,以海南种茶园受害较重[3]。但近年来开始在云南省普洱市、西双版纳傣族自治州部分茶园暴发性为害,已成为云南茶园的新发害虫。对云南茶园茶谷蛾生物学特性的调查研究显示:全年有4 个幼虫为害时期,以第1 代和第4 代密度较大;以2~3 龄幼虫在虫苞内取食成叶并过冬,翌年2 月下旬―3 月上旬化蛹;有逐年加重趋势,致使茶叶产量损失严重、树势恢复困难,给茶叶生产带来巨大损失[1,3]。
虫害防治一直是茶树植保及茶产业发展中的重大课题,作为应急防控措施,化学防治见效快,但往往伴随一系列农残、抗药性、不可持续、茶叶品质下降等问题。生物防治作为一种绿色安全、可持续发展的防控措施,已成为害虫防治的重要技术,对于减少化学农药使用、保护环境及形成良好的绿色防控措施有重要意义[4,5]。已有诸多研究[6-8]发现:保护和利用天敌昆虫来控制害虫种群数量、提高作物产量和品质能取得较好的防控效果。天敌昆虫在自然界中大量存在,对于某些害虫发生、成灾起着制约作用,对于维持生态平衡、保持物种多样性起着重要作用,主要包括寄生性昆虫和捕食性昆虫。研究发现,寄生性昆虫如螟黄赤眼蜂TrichogrammachilonisIshii、夜蛾黑卵蜂Telenomusremus(Nixon)[9]等能寄生草地贪夜蛾Spodopterafrugiperda(J.E.Smith);捕食性蝽如蠋蝽Armachinensis(Fallou)、叉角厉蝽Eocantheconafurcellate(Wolff)也有很好的控害效果[10],事实上对于草地贪夜蛾的捕食性天敌昆虫已经涉及蝽、瓢虫、草蛉、蠼螋等5 目12 科58 种[9];而对斜纹夜蛾Spodopteralitura天敌资源的研究也包括丽草蛉[4]、环斑猛猎蝽Sphedanolestesimpressicollis[11]等。但目前对茶谷蛾的天敌防控还鲜有报道。
叉角厉蝽属半翅目Hemiptera、蝽科Pentatomidae、益蝽亚科Asopinae、厉蝽属Eocanthecona[12]。一年可发生多代,据报道,该蝽具有发生早,时间长,捕食量大,捕食范围广,活动能力强等特点,可用来防治鳞翅目、膜翅目、鞘翅目、半翅目等40 种以上的幼虫,其中尤喜鳞翅目幼虫,故而又有“杀虫王”之称[12]。在绿额翠尺蛾ThalassodesproquadriaInouce[13]、黄野螟Heortiavitessoides幼虫[14]、亚洲玉米螟Ostriniafurnacalis幼虫[15]、草地贪夜蛾[16]、斜纹夜蛾[17,18]等的研究中均得到了较好的捕食控害效果。捕食功能反应是生物防治中研究捕食性天敌捕食能力的重要指标之一[19],为探索防治茶谷蛾的天敌昆虫资源,本文开展了叉角厉蝽对该虫的捕食作用、捕食功能反应、种内干扰作用及田间控害试验,以期为茶谷蛾的生物防治提供理论依据。
1 材料与方法
1.1 供试昆虫
叉角厉蝽:广州绿农夫农业服务有限公司提供,供试前禁食24 h。
茶谷蛾:各虫龄幼虫,于2021 年3 月采自云南省勐海县云南省农业科学院茶叶研究所基地内,并带回室内用新鲜茶树叶片饲养至化蛹,待蛹羽化后将成虫放入40 cm×40 cm×40 cm 的养虫笼内,喂以10%蜂蜜水,放入茶苗供其产卵、孵化,繁殖两代以上,选择健康幼虫进行试验。该研究的室内试验均在温度(26±1)℃、相对湿度65%±10%,光周期16L∶8D 的条件下完成,且均不人为干涉试验对象生活习性,因茶谷蛾吐丝结苞未受到叉角厉蝽攻击视为存活。
1.2 叉角厉蝽对茶谷蛾幼虫的捕食能力测定
叉角厉蝽选取4~5 龄若虫及成虫;茶谷蛾设1~2、3~4、5~6 龄3 个虫龄,共6 个处理。用足量的茶谷蛾供叉角厉蝽取食24 h 后再饥饿处理24 h,用于室内捕食能力测定。
在底部边长15 cm 的塑料透明养虫盒(15 cm×15 cm×20 cm)中放1 头叉角厉蝽及10 头茶谷蛾幼虫,每盒放置一个浸湿脱脂棉,每处理重复5 次,24 h 及48 h 后记录每盒茶谷蛾的死亡数。试验时放入足量茶树叶片供其取食防止茶谷蛾自相残杀,并计算单头叉角厉蝽的捕食量及捕食率。捕食率(%)=茶谷蛾幼虫死亡数/茶谷蛾幼虫总数×100%。
1.3 叉角厉蝽对茶谷蛾幼虫的捕食功能反应
叉角厉蝽选取4~5 龄若虫及成虫;茶谷蛾分为1~2、3~4、5~6 龄3 个虫龄阶段,密度设为1∶3、1∶6、1∶9、1∶12。每盒放1 头叉角厉蝽,其他处理及重复数同1.2,每处理设置对照(不放置叉角厉蝽),记录自然死亡率。24 h 后记录叉角厉蝽捕食量,并以自然死亡率校正。
Holling Ⅱ圆盘方程:Na=aNTt/(1+aThN);Na为茶谷蛾的被捕食量,N为茶谷蛾的密度;Tt为叉角厉蝽搜寻茶谷蛾的总时间(本试验为1 d),Th为处理时间(叉角厉蝽捕食1 头茶谷蛾所需的时间),a为叉角厉蝽对茶谷蛾的瞬时攻击率。通过倒数法将方程化简为:1/Na=1/aN+Th。模拟方程Y=A+BX,令1/Na=Y;Th=A;1/a=B;1/N=X[20]。当茶谷蛾密度N趋近于无穷时,1/N趋近于0,从而得到理论最大日捕食量,Namax=1/Th,以a/Th来反应叉角厉蝽对茶谷蛾的捕食效能。
1.4 叉角厉蝽对茶谷蛾幼虫的搜寻效应
搜寻效应方程为S=a/(1+aThN),其他参数同上[21]。
1.5 干扰反应对叉角厉蝽的捕食作用影响
叉角厉蝽设4~5 龄若蝽及成虫,与不同虫龄茶谷蛾按照1∶4 比例设置。每个养虫盒内放置4、8、12、16 头茶谷蛾,然后依次放入1、2、3、4 头4~5 龄叉角厉蝽若虫及成虫,每处理5 次重复。24 h 后统计被捕食的茶谷蛾幼虫数量。
干扰反应方程为:E=QP-m(E=Ne/NP),方程中E是捕食作用率,Q为搜寻系数,P为叉角厉蝽密度,m 为干扰常数,Ne为叉角厉蝽捕食茶谷蛾的数量,N为茶谷蛾密度[21,22]。
1.6 叉角厉蝽对茶谷蛾的田间控害试验
叉角厉蝽对茶谷蛾的田间控害试验在云南省农业科学院茶叶研究所科研基地内(E:100.44、N:21.95)进行,海拔1200 m,年均气温10 ℃,进行套袋试验时为5 月中旬湿度70%左右,气温26 ℃左右。试验设叉角厉蝽4~5 龄若虫及成虫两个虫态,茶谷蛾幼虫设1~2 龄、3~4 龄、5~6 龄3 个处理,共计6 个处理,每处理5 次重复。试验前将植株上其他生物清除,仅保留茶谷蛾幼虫并接种虫量至每株20 头,接种不同虫态叉角厉蝽1 头,使用立方体纱笼(60 目)将茶树整株罩住以隔绝其他生物,避免其他生物影响试验结果。对照茶树植株同样使用纱网(60 目)笼罩以避免其他昆虫影响试验,重复5 次。释放叉角厉蝽1、3、5、7 d 后调查茶谷蛾幼虫的剩余存活虫数。虫口减退率(%)=(茶谷蛾幼虫总数―剩余存活数)/茶谷蛾幼虫总数×100%;校正防效:A=(B―C)/(1―C)×100%。式中:A 为校正防效;B 为试验前后处理区虫口减退率;C 为试验前后对照区虫口减退率;A、B、C 均以百分数表示。本试验只探究茶谷蛾幼虫期,周期为7 d,不存在繁殖情况,且套袋处理不存在迁入迁出及其他昆虫影响,所以对照区数据以自然存活数记录。
1.7 数据统计与分析
所有数据均用Excel 2010 计算均值,再使用SPSS 19.0 进行单因素方差分析及Holling Ⅱ圆盘方程拟合,运用Duncan 氏新复极差法比较不同数据组间差异。
2 结果与分析
2.1 叉角厉蝽对茶谷蛾幼虫的捕食行为
叉角厉蝽通过触角上下左右摆动来搜索猎物,当接近茶谷蛾幼虫时,触角停止摆动,伸出口针,保持前伸或半前伸长状态,停顿在虫体附近,试探性接触虫体,伺机将口针刺入幼虫体内。口针通常从低龄幼虫体侧或腹部、尾部插入(偶尔从背部刺入),而捕食茶谷蛾高龄幼虫时,经常从其腹部末端插入,其次为头壳部位;或者松开口针更换刺吸部位,通过多次刺入最终使茶谷蛾幼虫逐渐静止后开始吸食虫体。
2.2 叉角厉蝽对茶谷蛾幼虫不同虫态的捕食能力测定
叉角厉蝽不同虫态对茶谷蛾不同虫龄幼虫的捕食量及捕食率见表1。叉角厉蝽同一虫态对不同虫龄茶谷蛾幼虫的捕食能力存在明显差异。总体表现为随着茶谷蛾幼虫虫龄增大,4~5 龄若蝽的捕食能力降低,而成蝽的捕食能力增强。其中4~5 龄若蝽对1~2 龄幼虫的捕食能力最大,日捕食量为7.40 头,捕食率达74.00%;对3~4 龄幼虫的日捕食量为3.2 头,捕食率为32.00%;对5~6 龄幼虫的捕食能力最小,日捕食量为1.60 头,捕食率为16.00%。成蝽则对1~2 龄幼虫的捕食能力最小,日捕食量为1.80 头,捕食率为18.00%;对3~4 龄幼虫的日捕食量为5.4 头,捕食率为54.00%;对5~6 龄幼虫的捕食能力最强,日捕食量为7.6 头,捕食率为76.00%。

表1 叉角厉蝽对茶谷蛾幼虫的捕食量(头)Table 1 Predation amount of E.furcellate on A.rhombata larvae
叉角厉蝽不同虫态对同一虫龄茶谷蛾幼虫的捕食结果显示,4~5 龄若蝽对1~2 龄幼虫的捕食量高于成虫,而成蝽对3~4 龄、5~6 龄幼虫的捕食量高于4~5 龄若蝽。
2.3 不同虫态叉角厉蝽对茶谷蛾幼虫的捕食功能反应
对不同虫态叉角厉蝽捕食不同虫龄茶谷蛾幼虫的数据进行拟合,1/N与1/Na之间的相关系数均大于r0.01(0.917),表明二者极相关。4~5 龄若蝽对不同虫龄幼虫的捕食功能反应符合 Holling II 方程(表2),对不同虫龄幼虫的捕食能力从大到小依次为1~2 龄幼虫、3~4 龄幼虫、5~6 龄幼虫,日最大捕食量分别为28.6588、8.2683、1.8658 头;瞬时攻击率依次为1.057、0.932、0.987。从捕食效能指标看,对1~2 龄幼虫的捕食能力(30.2886)大于3~4 龄(7.7098)及5~6 龄幼虫(1.8421)。由以上研究结果可知,4~5 龄若蝽对茶谷蛾低龄幼虫的捕食效果显著优于高龄幼虫。成蝽对不同虫龄幼虫的捕食功能反应亦符合Holling II 方程(表3),捕食能力从大到小依次为5~6 龄幼虫、3~4 龄幼虫、1~2 龄幼虫,日最大捕食量分别为11.5936、10.7190、9.8288 头;瞬时攻击率依次为1.283、0.930、0.835。从捕食效能指标看,对5~6 龄幼虫的捕食能力(14.8712)大于3~4 龄(9.9634)及1~2 龄幼虫(8.2109)。由以上研究结果可知,成蝽对茶谷蛾低龄幼虫的捕食效果显著低于高龄幼虫。

表2 4~5 龄叉角厉蝽对茶谷蛾幼虫的捕食功能反应Table 2 Predation functional response model of 4th―5th E.furcellate nymph to A.rhombata larvae

表3 叉角厉蝽成虫对茶谷蛾幼虫的捕食功能反应Table 3 Predation functional response model of adult E.furcellate to A.rhombata larvae
分别将叉角厉蝽对茶谷蛾各虫龄幼虫捕食功能反应的参数a和Th代入相应捕食功能反应方程中,计算得到理论捕食量进行方程拟合(图1、图2),并与实测值进行χ2检验,χ2值均小于相应自由度下的χ24,0.05,差异不显著(P>0.05),说明Holling Ⅱ圆盘方程拟合结果较为理想,模型能够较好地反应叉角厉蝽对茶谷蛾幼虫不同密度下的捕食量变化规律。

图1 4~5 龄若蝽对茶谷蛾幼虫的捕食功能反应Fig.1 Predation functional response of 4th―5th E.furcellate nymph to A.rhombata larvae
2.4 叉角厉蝽对茶谷蛾不同虫龄幼虫的搜寻效应
根据寻找效应方程和捕食功能反应的参数值,可以得到不同虫态叉角厉蝽对不同虫龄茶谷蛾幼虫的寻找效应(图3、图4)。当茶谷蛾幼虫的种群密度为3 头/皿、6 头/皿、9 头/皿、12 头/皿时,4~5 龄若蝽对1~2 龄幼虫的寻找效应为0.9516、0.8654、0.7935、0.7326;对3~4 龄幼虫的寻找效应为0.6967、0.5561、0.4628、0.3962;对5~6 龄幼虫的寻找效应为0.3816、0.2365、0.1713、0.1343。成蝽对1~2 龄幼虫的寻找效应为0.6657、0.5533、0.4733、0.4136;对3~4 龄幼虫的寻找效应为0.7376、0.6114、0.5220、0.4555;对5~6 龄幼虫的寻找效应为0.9630、0.7709、0.6427、0.5511。由图3、图4 可知,叉角厉蝽对茶谷蛾幼虫的寻找效应随其密度的增加而降低,4~5 龄若蝽对低龄幼虫的搜寻效应大于高龄幼虫,而成蝽对高龄幼虫的搜寻效应大于低龄幼虫。

图3 4~5 龄若蝽对茶谷蛾幼虫的寻找效应Fig.3 Searching efficiency of 4th-5th E.furcellate nymph to A.rhombata larvae

图4 叉角厉蝽成虫对茶谷蛾幼虫的寻找效应Fig.4 Searching efficiency of adult E.furcellate to A.rhombata larvae
2.5 干扰作用对茶谷蛾幼虫捕食作用率的影响
保持叉角厉蝽与茶谷蛾不同虫龄幼虫都以相同比例的密度成倍增长,叉角厉蝽的总捕食量随猎物密度的增加而增加,但其平均捕食量及捕食作用率逐渐下降,说明叉角厉蝽之间存在种内干扰作用,种内干扰作用会对捕食效应有一定的影响(表4~6)。按照Hassell 提出的符合这种变化规律的方程E=QP-m拟合表4~6 数据。当4~5 龄若蝽的密度分别为1、2、3、4 头/皿时,对1~2 龄幼虫的平均捕食量分别为3.80、3.30、2.86、2.60 头,捕食作用率分别为0.9500、0.4125、0.2389、0.1625,通过计算搜寻常数Q及干扰系数m,拟合干扰方程为E=0.9665P-1.1280;对3~4 龄幼虫的平均捕食量分别为2.60、2.70、2.0、2.15 头,捕食作用率分别为0.6500、0.3375、0.1667、0.1344,拟合干扰方程为E=0.6765P-1.181;对5~6 龄幼虫的平均捕食量分别为0.80、1.10、1.20、1.20 头,捕食作用率分别为0.200、0.1375、0.1000、0.0750,拟合干扰方程为E=0.2081P-0.6950。将各处理的捕食作用率实测值与理论值进行χ2检验,均为χ2<χ24,0.05,差异不显著(P>0.05),说明拟合曲线符合因干扰作用而引起的捕食效应变化规律。

表4 干扰作用对叉角厉蝽4~5 龄若虫捕食茶谷蛾幼虫的影响(1~2 龄虫)Table 4 Affection of interference effect on 4th―5th E.furcellate nymph’s predation to A.rhombata larvae (1st―2nd instar)

表5 干扰作用对叉角厉蝽4~5 龄若虫捕食茶谷蛾幼虫的影响(3~4 龄虫)Table 5 Affection of interference effect on 4th―5th E.furcellate nymph’s predation to A.rhombata larvae (3rd―4th instar)

表6 干扰作用对叉角厉蝽4~5 龄若虫捕食茶谷蛾幼虫的影响(5~6 龄虫)Table 6 Affection of interference effect on 4th―5th E.furcellate nymph’s predation to A.rhombata larvae (5th―6th instar)
按照Hassell[22]提出的符合这种变化规律的方程E=QP-m拟合叉角厉蝽成虫对茶谷蛾幼虫的捕食数据(表7~9)。在1、2、3、4 头/皿的密度下,成蝽对1~2 龄幼虫的平均捕食量分别为3.0、2.0、2.0、2.25头,捕食作用率分别为0.7500、0.2500、0.1667、0.1406,方程拟合结果为E=0.6857P-1.2330;对3~4 龄幼虫的平均捕食量分别为3.80、3.40、2.73、2.5 头,捕食作用率分别为0.9500、0.4250、0.2278、0.1563,干扰方程拟合为E=0.9807P-1.310;对5~6 龄幼虫的平均就捕食量分别为4.0、3.4、3.27、3.0 头,捕食作用率分别为1.0000、0.4250、0.2722、0.1875,干扰反应方程拟合结果E=0.9946P-1.199。将各处理的捕食作用率实测值与理论值进行χ2检验,均为χ2<χ24,0.05,差异不显著(P>0.05),说明拟合曲线符合因干扰作用而引起的捕食效应变化规律。
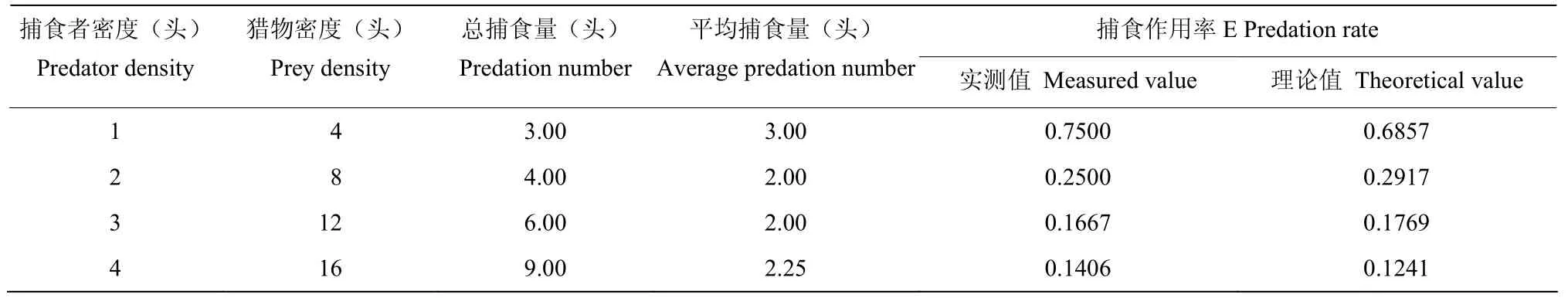
表7 干扰作用对叉角厉蝽成虫捕食茶谷蛾幼虫的影响(1~2 龄虫)Table 7 Affection of interference effect on E.furcellate adult’s predation to A.rhombata larvae (1st―2nd instar)

表8 干扰作用对叉角厉蝽成虫捕食茶谷蛾幼虫的影响(3~4 龄虫)Table 8 Affection of interference effect to E.furcellate adult’s predation on A.rhombata larvae (3rd―4th instar)
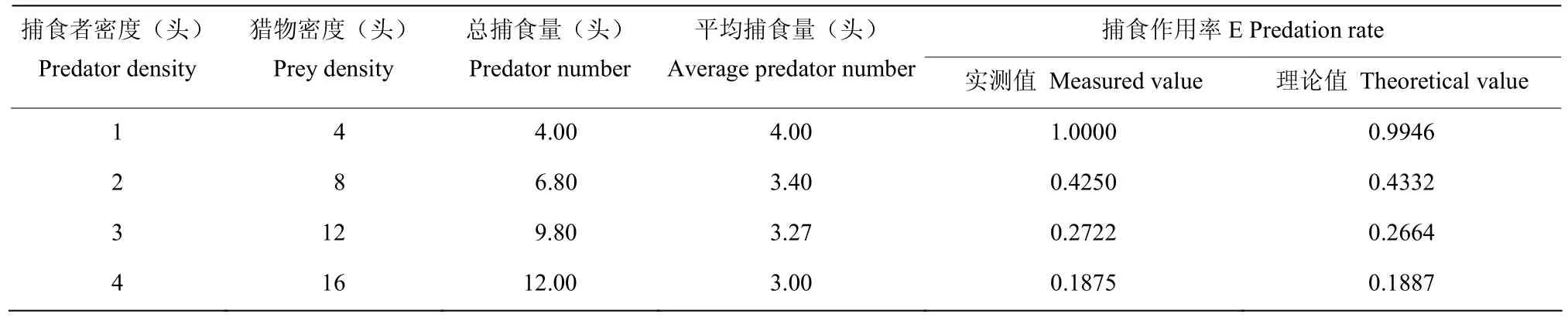
表9 干扰作用对叉角厉蝽成虫捕食茶谷蛾幼虫的影响(5~6 龄虫)Table 9 Affection of interference effect to E.furcellate adult’s predation on A.rhombata larvae (5th―6th instar)
2.6 叉角厉蝽对茶谷蛾幼虫的田间控害效果
4~5 龄若蝽对低龄幼虫的防效显著高于5~6 龄幼虫,并随时间延长而增加,在第7 d 时,对1~2 龄幼虫的防效达42%,对3~4 龄幼虫的防效达36%;成蝽在整个试验周期内对3~4 龄幼虫的防效较好,在第7 d 时达到47%,5~6 龄幼虫次之,第7 d 时为34%(表10)。
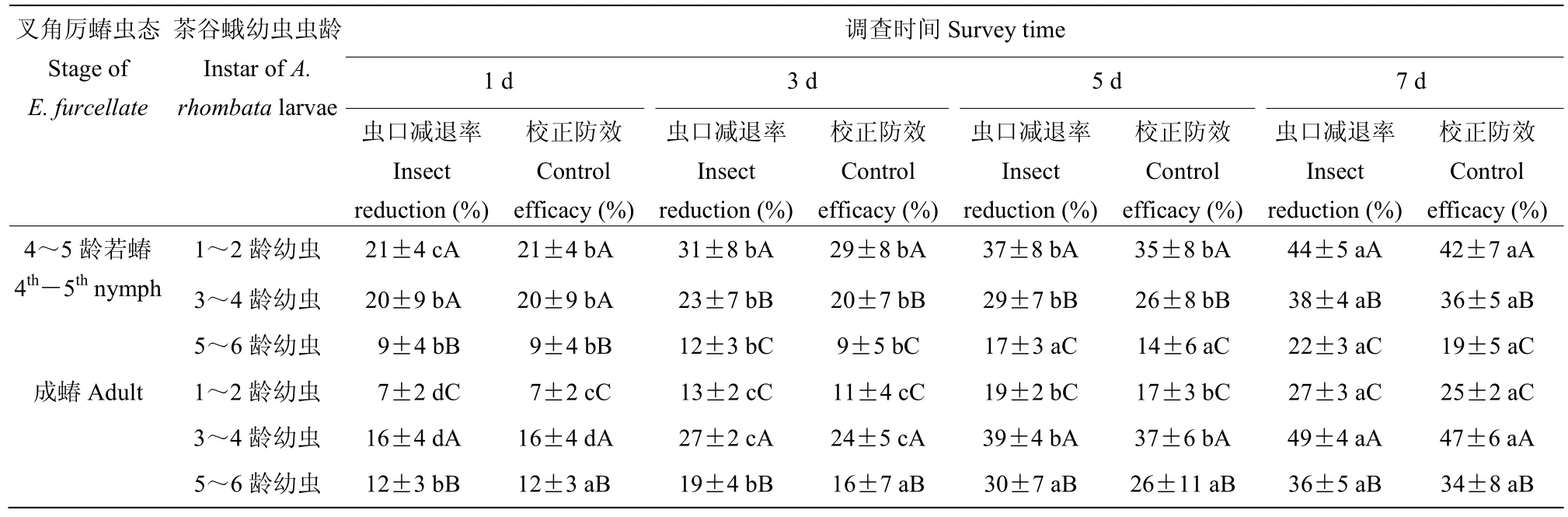
表10 叉角厉蝽对茶谷蛾的田间防效Table 10 Control effects of E.furcellate on A.rhombata
3 讨论
叉角厉蝽对茶谷蛾幼虫的捕食能力较强。茶谷蛾自2014 年在云南茶区暴发性成灾以来,已成为当地为害严重的新发害虫。叉角厉蝽作为诸多农林果蔬害虫的重要天敌资源,能捕食40 种以上的幼虫,其中尤喜鳞翅目幼虫[12],其对于同属鳞翅目害虫的茶谷蛾来说具有很大的捕食潜能。本试验通过对叉角厉蝽捕食茶谷蛾的行为观察及捕食能力测定,发现4~5 龄若蝽对茶谷蛾1~2 龄幼虫的日捕食量及捕食率最高,分别为7.4 头和74.00%;而成蝽则对5~6 龄幼虫有最大日捕食量及捕食率,分别为7.6 头及76.00%,不同虫态叉角厉蝽对不同虫龄茶谷蛾表现出较好的搜索与捕食能力。这表明叉角厉蝽对茶谷蛾有一定的控害效果,且具有虫龄选择性,但不排除由于在培养皿内进行试验,茶谷蛾利用叶片吐丝结苞的习性受限,使得自身更容易被叉角厉蝽搜索、攻击、取食。因此,叉角厉蝽对茶谷蛾的防治能力还应以田间试验为主。
叉角厉蝽的捕食能力与茶谷蛾虫龄及躯体大小有关。蠋蝽对斜纹夜蛾的捕食效能随害虫虫龄增大而降低[23];叉角厉蝽对黄野螟3 龄幼虫的捕食量也高于5 龄幼虫[14],说明天敌昆虫的捕食能力受限于猎物的自身情况。本研究中对于茶谷蛾1~2 龄幼虫来说,4~5 龄若蝽的捕食能力较强,随着茶谷蛾虫龄增大、躯体增大,4~5 龄若蝽的捕食效能降低。在捕食行为观察中亦发现,在若蝽与茶谷蛾的对抗中,高龄幼虫躯体较大,通过扭动躯体逃脱刺吸的概率大于低龄幼虫;相反,低龄幼虫一旦被若蝽搜索到则很难逃脱直至被吸食死亡;成蝽对高龄幼虫则更易搜索并攻击成功。说明不同虫态叉角厉蝽的捕食能力对茶谷蛾幼虫具有虫龄及大小选择性。
利用Holling Ⅱ功能反应模型可有效评价天敌昆虫对靶标猎物的捕食适合度[17]。本研究中叉角厉蝽对茶谷蛾幼虫的捕食能力符合Holling Ⅱ模型。4~5 龄若蝽对茶谷蛾幼虫的瞬时攻击率依次为1~2 龄>5~6 龄>3~4 龄,日最大捕食量及捕食效能依次为1~2 龄>3~4 龄>5~6 龄;成蝽则相反,瞬时攻击率、日最大捕食量及捕食效能均为5~6 龄>3~4 龄>1~2 龄。可能是因为高龄幼虫的体型较大,活动能力较弱,对于活动能力很强的成蝽更容易被搜索和攻击成功。由此可见,在不同的茶谷蛾幼虫期,释放不同虫态的叉角厉蝽更有利于发挥控害作用。
寻找效应是捕食者在捕食过程中对猎物的搜寻攻击行为效应,受到空间及自身竞争干扰,捕食者的捕食作用率会下降[24]。本研究中不同虫态叉角厉蝽对茶谷蛾幼虫的搜寻效应均随茶谷蛾幼虫密度的增大而下降,这一结果与叉角厉蝽对斜纹夜蛾的捕食功能反应结果相似[24]。其中,4~5 龄若蝽的搜寻效应随茶谷蛾虫龄增大而降低,成蝽的搜寻效应则随虫龄增大而增加,但整体的搜寻效应均大于4~5 龄若蝽。
捕食者对猎物的捕食作用与捕食者及猎物的密度相关,随着二者密度增加,捕食者及猎物的相互干扰作用明显[17]。本研究中当空间和猎物比例一定时,随着叉角厉蝽和茶谷蛾密度的增加,叉角厉蝽的捕食作用率均下降,表明二者之间存在干扰反应,这说明虽然叉角厉蝽的捕食量随捕食者及猎物密度的增加而增加,但其寻找效应会受到个体之间的觅食竞争及猎物密度的干扰,导致整体捕食效能下降;同一密度下,随着虫龄增大,4~5 龄若蝽的捕食作用率下降,成蝽则上升,这与建立的Holling Ⅱ功能反应模型一致,即随着茶谷蛾虫龄增大,成蝽的捕食效能增加,4~5 龄若蝽的降低。
在田间进行套袋控害试验发现,4~5 龄若蝽对1~2 龄幼虫的防效达42.00%,3~4 龄及5~6 龄幼虫次之,与室内测定结果一致,可能是相较于高龄幼虫,低龄幼虫活动能力较弱,体型较小,若蝽更容易攻击成功并吸食;不同于室内测定结果的是,成蝽对3~4 龄幼虫有更大防效,5~6 龄幼虫次之,可能是因为高龄幼虫活动能力较强,且田间有茶树叶片可作遮挡,有避让攻击能力。且茶谷蛾在田间可利用茶树叶片吐丝结苞,既可居于苞内取食,又可防御天敌搜索,因此相对于室内,田间捕食能力有所降低。
根据试验结果,叉角厉蝽对茶谷蛾具有一定的控害效能,可作为防治茶谷蛾的天敌昆虫之一,在茶谷蛾暴发的不同幼虫期,应选择不同虫态的叉角厉蝽才能达到较好的捕食控害效果。如在每年茶谷蛾暴发初期,虫龄较小时,可以释放一定数量的若蝽,将茶谷蛾的数量控制在较低的水平,从而预防茶谷蛾的大暴发;在茶谷蛾幼虫后期,释放一定数量的成蝽,减少茶谷蛾化蛹、羽化,抑制其继代繁殖。但在田间开展生物防治时,叉角厉蝽对茶谷蛾的捕食作用不仅会受到自身密度、猎物密度的影响,还受到自然界中温度、湿度、化学药剂等一些不稳定气候因子的影响[6,10,25]。因此应结合茶谷蛾的种群动态及天气情况,并充分考虑叉角厉蝽的释放密度,才能达到较好的防治效果。
