岷江源区两种优势针叶树当年生小枝性状与生物量分配随海拔的分异规律
2023-12-25王金牛王丽华田炳辉盖艾鸿
张 宁,王金牛,石 凝, 4,王丽华,朱 牛,田炳辉,张 林,盖艾鸿
1 甘肃农业大学资源与环境学院,兰州 730070
2 中国科学院成都生物研究所,成都 610041
3 西藏生态安全屏障生态监测网络芒康生物多样性与生态监测站,昌都 854000
4 中国科学院大学,北京 100049
5 阿坝师范学院,汶川 623002
6 中国科学院青藏高原研究所,北京 100101
植物性状是植物对不同生境的适应特征,受谱系或环境条件的影响[1],并对植物适应和生存有显著影响[2]。当年生小枝是植物分枝系统中活跃的部分,其上生长着叶片、花和果实,次生组织较少,内部营养物质和碳水化合物运输速率快,能较好地反映多年生植物的近期生长情况[3]。树冠上枝叶的数量、形态及分布直接影响乔木的生长及繁殖。叶数量与展叶效率的变化直接影响光拦截效率[4];叶面积通过光合与蒸腾作用影响植物对水热及养分的利用效率;高比叶面积、低叶干物质含量的物种对体内养分保有能力更强[1]。小枝茎构型的改变同时对叶片的空间排布和养分运输造成影响,优越的光获捕体系能最大程度地提高碳收益[5]。由此,枝叶构件的内在相关关系反映了植株个体内部的资源利用水平、构件间的分配及权衡关系。有效的资源配置会影响到物种的生存和繁殖,即植株体内资源的分配权衡关系是植物不同生长策略的重要特征。譬如,适宜生境中植物倾向于r-对策,将大量资源分配至繁殖结构;生长受限时植物倾向于K-对策,即提高茎叶生物量占比[6]。一般来说,小枝内生物量分配可以从茎和叶两个水平进行分析。最优分配理论认为,植物会将更多的资源分配至生长受限的构建单元以维持植物最优的生长速率[5]。异速生长关系可以预测植物器官生物量分配的内在规律并进行量化,揭示物种部分与整体或部分与部分间的差异和对比。将两种理论结合起来,可以很好的揭示植物对环境变化的响应,进一步明确其生态适应策略。
近年来,全球增温对高纬度和高海拔地区的森林生态系统造成深刻影响。青藏高原已成为当今世界全球气候变化下的敏感保护区,会在未来100年内由于气候的变化导致区域内植物地理分布发生剧烈改变[7]。亚高山森林同时也是气候变化的敏感指示因子,研究其对气候变化的响应对掌握全球气候变化具有深刻意义。高山地区具有复杂的环境异质性,小范围内空间垂直尺度上的海拔变化,会使得温度、降水和光照强度等非生物因素呈梯度性变化[8],进而造成植物性状发生相应改变,是研究植物性状-气候变化响应的天然实验室[9]。近年国内外有关当年生小枝性状研究主要集中在不同物种或群落在生长梯度上枝叶性状的变化[1]、小枝-叶生长权衡关系上,包括茎构型或茎生物量与叶数量[10]、叶面积大小[11]、叶生物量[12]及展叶效率[4]间的关系等、叶柄与叶片生物量分配关系[13]、地上地下部分生长限制[14]及比例研究[15]等。研究指出,随着海拔的上升,植物叶片变得小而厚,比叶面积降低,增加光合速率的同时减少蒸发面积[5]。受区域小气候和海拔影响,植物生物量分配沿海拔梯度变化明显。高海拔的植物为了资源获取倾向于小枝茎的投资,但与提高支撑能力的茎粗间存在一定权衡[10]。近年来,全球变暖造成森林生态系统的改变,鉴于青藏高原对气候变化的敏感性,在青藏高原亚高山森林生态系统内研究树木当年生小枝性状及生物量分配关系随海拔梯度的变化能有效地揭示不同生境下树木近期的生长情况,便于诠释全球气候变化背景下森林种群的分布与适应策略。
亚高山常绿针叶林是青藏高原东南部分布的主要森林类型,青藏高原东缘部分从东南到西北降水量逐渐减少,原以冷杉属为优势种的喜阴湿的森林逐渐被更耐干冷的云杉林取代[16]。紫果云杉(PiceaPurpurea)和岷江冷杉(Abiesfaxoniana)为我国本土特有的高山耐寒常绿针叶乔木。紫果云杉在中国分布范围为30°36′00″—36°18′00″N, 100°24′00″—105°12′00″E,海拔范围为2600—3800 m,岷江冷杉为29°48′00″—35°30′00″N, 100°24′00″—105°12′00″E,2600—3800 m,两者均呈现出分布的海拔高度同经纬度成反比的规律。在亚高山海拔跨度较大范围的阴坡、半阴坡广泛分布。森林作为最顶级的生态演替结果,与高原湿地和草原共同参与岷江上游区域生态稳定的维护[17]。全球气候变化背景下,冷云杉的生长发育及分布变迁会导致青藏高原原本生态稳定的失调,从而影响其生态服务功能的实现。当前有关青藏高原东缘地区紫果云杉和岷江冷杉的研究多见于种群结构[18],群落生物多样性[19],凋落物分解[20]、树线变化[21]等,但针对全球气候变化下两优势种当年生小枝的响应与适应策略缺乏研究。本文以岷江源区亚高山森林分布的优势种紫果云杉和岷江冷杉的当年生小枝为研究对象,通过比较不同海拔间植物小枝和叶片功能性状的相关关系与种间种内的变异,分析茎叶生物量分配及其生长关系的权衡并探讨小枝大小作为协变量所起的作用,对当年生小枝的大小所起到的作用进行较充分的探讨,拟揭示两种常绿针叶树对海拔变化的长期适应及生长策略,为全球变暖下的种群保护、维持及变迁提供基础数据及理论依据。
1 地区与方法
1.1 研究区概况
本研究样地位于川西高原岷江源区的卡卡山(32°59′03″N,103°40′12″E;海拔:3550—4000 m),青藏高原东南缘的该区域[21],属于岷江源国家湿地公园所辖地段,是成都市重要的水源涵养区。沿着九黄旅游线承载着关键生态系统功能与服务的多样性,也是当地社区与居民获取生计的主要来源[22]。该区域作为高山与高原的过渡地带,亦是森林到高寒草地的林草交错区域,受亚热带季风影响,该区域日温差距较明显,年均气温3—7 ℃。1月最低气温-8—15 ℃,7月最高气温为9—15 ℃。年均降水量稳定在700 mm左右,集中分布在夏秋季节,冬季降水少。在10月底开始有降雪事件发生,积雪期一般为5个月,厚度可达15 cm[23]。
研究区样地生境异质性较大,植被类型多样,具有较高的生物多样性。亚高山森林的原生建群乔木为岷江冷杉(Abiesfaxoniana)、紫果云杉(PiceaPurpurea),灌木主要包括高山柳(Salixcupularis)、窄叶鲜卑花(Sibiraealaevigata)、陇蜀杜鹃(Rhododendronprzewalskii)、小叶杜鹃(Rhododendroncapitatum)等[18]。岷江源区岷江冷杉与紫果云杉分布数据来自全球生物多样性数据库(GBIF),物种分布图如图1所示。为了分析卡卡山优势乔木紫果云杉和岷江冷杉当年生小枝性状随海拔的变化,沿着亚高山针叶林带(3500—3600 m)、乔灌混交林带(3650—3750 m)和亚高山灌丛带(3800—3850 m)内分别设置3条样带进行研究,研究样点图见图1。

图1 物种分布及研究样地图
1.2 实验设计与样品采样
在研究区域目标物种的自然分布地,沿海拔梯度分别在3500—3550 m、3650—3700 m、3800—3850 m处开展调查取样工作。在每个海拔针对每个物种随机选择生长状况、胸径相似的健康标准木10株,标准木间最少间距30 m,经计算,云杉三个海拔上选取标准株胸径平均值±标准误为(0.421±0.082)m、(0.317±0.050)m、(0.461±0.082)m,在海拔间不存在显著差异;冷杉胸径平均值为(0.375±0.071)m、(0.375±0.039)m、(0.673±0.152)m,在海拔间不存在显著差异。每株标准木选择东西南北四个方位三年生枝条各一枝,然后取下枝条上的所有当年生小枝,共获得三个海拔上(3500—3550 m、3650—3700 m和3800—3850 m)云杉当年生小枝132枝、129枝、136枝;冷杉当年生小枝117枝、118枝、127枝。当年生小枝为枝条最末端到第一个分叉处[24]。采集当年生小枝上所有叶片,将研究样品装入袋密闭保存,测量其相关性状。
1.3 性状与方法
使用精度为0.01mm的游标卡尺对所得样品的茎长与茎粗进行测量。茎粗测定位置为当年生小枝与二年生小枝连接端茎横截面的直径长。在70 ℃下烘干茎和叶片至恒重,记录小枝上茎生物量和总叶生物量。获得的指标为:茎长(SL);茎粗(SD);比茎长(SSL);茎生物量(SB);总叶生物量(LB)。其中比茎长(SSL)=茎长(SL)/茎生物量(SB)。
1.4 权衡值计算
不同环境会影响植物性状的不同表达,成对性状之间存在一定关系的权衡。为了直观的表示两者之间的关系,采取Bradford[25]等的方法计算并进行可视化展示。计算权衡值首先需要对性状的观测值进行标准化处理,减少量纲影响,用其标准化值Astd来代表性状的相对收益。计算方法如下公式所示:
其中AOBS、Amax和Amin分别代表性状A的观测值、最大值、最小值。权衡值则用使用成对性状标准值的均方根误差(RMSE)表示,在图上直观表达为成对性状标准值坐标点到对角线(1∶1)的距离(图2)。

图2 成对性状间权衡图
1.5 数据处理
对所获得性状数据进行正态分布检验,不满足的进行数据转换使其满足,其次对数据进行方差齐性检验,对满足的数据进行one-way ANOVA检验,不满足的进行非参数检验并进行两两对比检验。利用嵌套设计及其方差分析将小枝功能性状变异分解为海拔水平、个体水平及小枝水平三个层次。使用Pearson相关法判断小枝功能性状与海拔间关系。对不同海拔茎叶生物量分配比例进行多重比较。根据异速生长模型[26]y=βxα对不同海拔研究对象的成对生物量进行对比分析,该公式可以转化为log(y)=log(β)+αlog(x),x和y代表研究对象成对的生物量,β为标准化常数,α为相关生长系数。当α=1时,表示两者为等速生长关系,α≠1时为异速生长关系。采用标准化主轴分析[26](SMA)方法来确定参数,使用R语言内置SMATR包完成。其他数据分析用SPSS Statistics 25完成,作图使用Origin 2021完成,研究样地示意图使用ArcGIS 10.8完成。
2 结果与分析
2.1 不同海拔下云冷杉的当年生小枝功能性状种内种间变异
岷江冷杉茎长与生物量在各个海拔均高于紫果云杉,茎粗、比茎长则小于云杉(表1),相同海拔植物性状表达不同,说明植物性状受到植物本身和环境的共同作用。不同海拔间卡卡山地区优势乔木的茎性状存在显著差异(表1,P<0.05),具体表现为云杉总叶生物量、茎生物量在3500—3550 m、3650—3700 m、3800—3850 m海拔间显著下降,比茎长则在三个海拔间显著上升。冷杉茎生物量、茎长则在3800—3850m处表现出最大值,在3650—3700 m处总叶生物量、茎生物量、茎粗表现出最小值,比茎长表现为最大值。在整个研究海拔上,种间小枝性状变异表现为茎生物量>总叶生物量>比茎长>茎长>茎粗,云杉在各研究海拔上持有较高的生物量变异,且总叶生物量、茎生物量和茎粗变异均强于冷杉,这说明,云杉通过改变不同器官生物量的积累积极响应海拔的变化。

表1 紫果云杉与岷江冷杉小枝性状及分异特征表
嵌套分析及方差分解结果(图3)显示,云杉当年生小枝功能性状总变异来源的解释率为85%—97%,海拔水平及小枝水平上的变异占茎叶生物量变异的大部分;冷杉当年生小枝功能性状总变异解释率为83%—92%,海拔水平及个体水平上的变异占茎叶生物量变异的大部分,其余所有性状均为海拔水平的解释率大于个体及小枝水平解释率。

图3 海拔、个体与小枝水平对植物性状变异的解释
2.2 不同海拔下云冷杉的当年生小枝及叶片性状相关性
紫果云杉总叶生物量、茎生物量与海拔间存在显著负相关关系(表2,P<0.001、P<0.01),比茎长与海拔间存在显著正相关关系(P<0.001),这说明紫果云杉在低海拔处拥有较高的茎叶生物量。岷江冷杉总叶生物量与海拔间均存在显著负相关关系(P<0.01),茎性状与海拔间不存在相关关系(P>0.05)这说明岷江冷杉小枝的茎不随海拔变化而显著变化。紫果云杉的总叶生物量、茎生物量、茎粗、茎长与比茎长两两间存在显著的相关关系,比茎长与其他性状间呈显著的负相关关系,其他两两间则为正相关。与云杉不同的是,岷江冷杉比茎长与茎长间存在显著的正相关关系(P<0.01)。两个物种茎叶性状与海拔间存在相似的相关性,反映出植物对环境的趋同适应特征,这说明,木本植物生长过程中,可能形成了一定的策略维度。
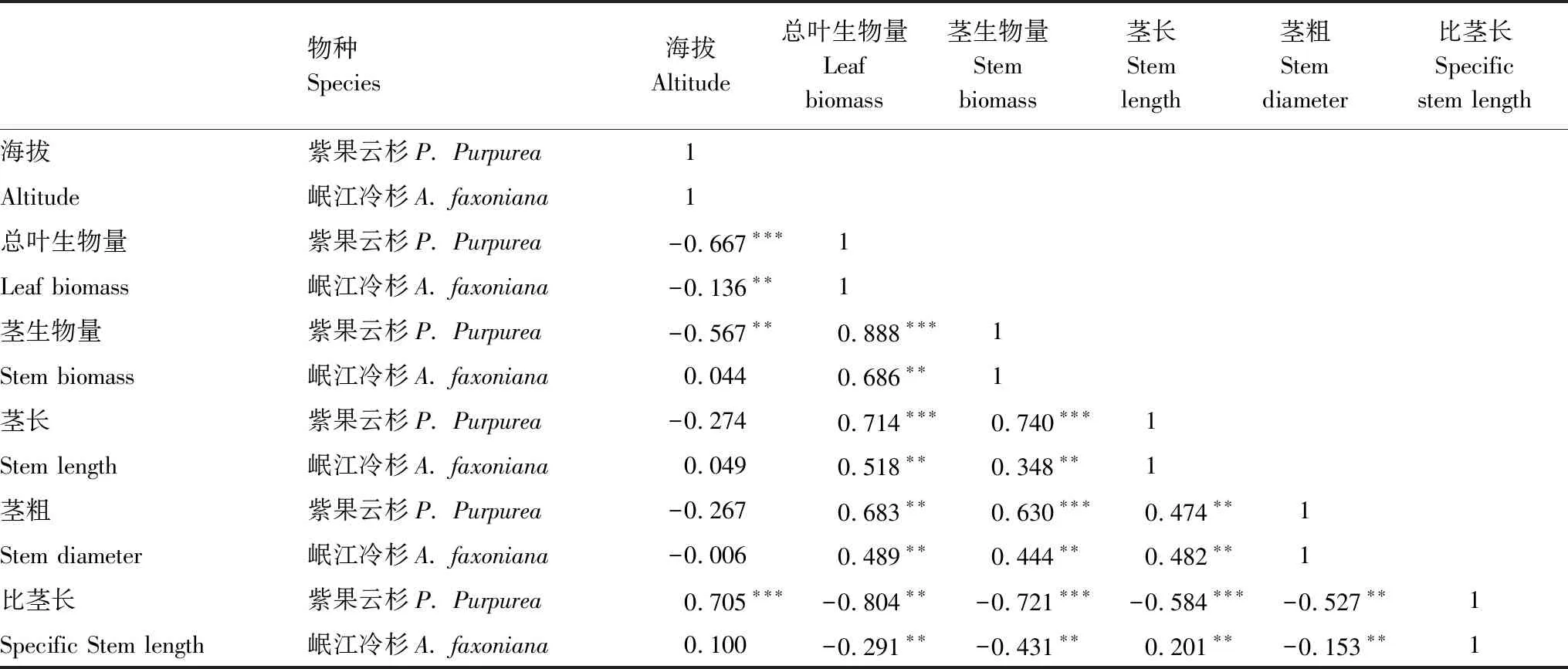
表2 紫果云杉和岷江冷杉小枝性状的相关系数和显著性
2.3 不同海拔下两物种的当年生小枝生物量分配及其生长关系的权衡
云杉在3500—3550 m海拔上升至3650—3700 m海拔时,由茎生物量倾向转变为总叶生物量倾向,茎生物量分配比例也由原本的33%显著降低至26%(图4,P<0.05);冷杉则无倾向变化,茎生物量分配比例变化不显著。在3650—3700 m海拔上升到3800—3850 m海拔时,云杉转变为茎质量偏好;冷杉则与前两个海拔叶质量偏好相反,倾向于茎质量,且茎生物量比例由原本的24%显著增加至28%(P<0.05)。这表明在3500—3550 m海拔处,云杉和冷杉偏好不同,但随着海拔的上升,两物种偏好相同,且茎、总叶生物量分配比例逐渐接近(27%∶73%; 28%∶72%)。

图4 不同海拔下紫果云杉与岷江冷杉茎叶部分生物量分配及其权衡关系
云杉茎生物量与总叶生物量在三个海拔处呈极显著的正相关关系(任一海拔内均是P<0.001,图5),3500—3550 m和3650—3700 m海拔上SMA斜率分别为1.436(95%置信区间为1.324,1.558)、1.113(95%置信区间为1.012,1.223),均显著高于1.0(P<0.001;P<0.05),表明茎生物量与总叶生物量呈现显著异速生长关系,且茎生物量积累大于总叶生物;3800—3850 m海拔上SMA斜率为1.133(95%置信区间为0.993,1.294),与1.0无显著差异(P>0.05),表明该海拔上二者呈现同速生长关系;茎叶生物量在中高海拔发生了截距漂移和共轴漂移,在中高海拔处存在共同斜率1.120(95%置信区间为1.037,1.209),共同斜率与1.0存在显著差异(P<0.05)。冷杉茎生物量与总叶生物量在三个海拔处同样呈极显著的正相关关系(任一海拔内均是P<0.001,图5),且SMA斜率分别为1.208(95%置信区间为1.090,1.338)、1.175(95%置信区间为1.060,1.302)、1.313(95%置信区间为1.181,1.461),均显著均显著高于1.0(任一海拔内均是P<0.001),表明冷杉三个海拔上茎生物量与总叶生物量呈现显著异速生长关系,且茎生物量积累大于总叶生物;三个海拔处存在共同斜率1.228(95%置信区间为1.157,1.305),共同斜率与1.0存在显著差异(P<0.05),低海拔与中海拔均和高海拔存在截距漂移,三个海拔间均存在共轴漂移(P<0.05)。

图5 紫果云杉与岷江冷杉相关生长关系图
云杉和冷杉茎长与茎粗间的生长关系表现出随茎叶生物量相关生长相同的规律,即云杉低海拔为异速生长,SMA斜率分别为1.441(95%置信区间为1.241,1.674)、1.195(95%置信区间为1.013,1.410),均显著高于1.0(P<0.001;P<0.05);高海拔为同速生长,SMA斜率为1.163(95%置信区间为0.992,1.364),与1.0无显著差异(P>0.05);三个海拔处存在共同斜率1.269(95%置信区间为1.158,1.391),共同斜率与1.0存在显著差异(P<0.05),三个海拔间均存在共轴漂移(P<0.05)。冷杉三个海拔处同样呈极显著的正相关关系(任一海拔内均是P<0.001,图4),且SMA斜率分别为1.301(95%置信区间为1.101,1.538)、1.187(95%置信区间为1.017,1.386)、1.289(95%置信区间为1.114,1.493),均显著均显著高于1.0(P<0.01,P<0.05,P<0.001),表明冷杉三个海拔上茎长与茎粗呈现显著异速生长关系,且茎长增加大于茎粗。三个海拔处存在共同斜率1.258(95%置信区间为1.150,1.375),共同斜率与1.0存在显著差异(P<0.05),高海拔与中海拔间发生共轴漂移(P<0.05)。
2.4 当年生小枝性状及生物量沿着海拔变化与小枝大小的关系
云杉、冷杉茎的性状随海拔梯度和植株大小的不同有显著差异(表3),具体表现为以茎长为协变量时,云杉茎粗、总叶生物量受海拔影响较小(PA=0.404,PA=0.524),主要受茎长度的影响(均为PL<0.001);冷杉总叶生物量受海拔影响较小(PA=0.322),主要受茎长的影响(PL<0.001)。以小枝生物量为协变量,云杉茎粗受海拔影响较小(PA=0.905),主要受小枝生物量的影响(PL<0.001);冷杉则表现出相同的规律,冷杉除茎粗外(PB<0.001=PA),其他性状均受到小枝生物量的显著影响(任一性状PB<0.001,PA>0.05)。整体而言,以不同小枝性状来代表小枝作为构件对整体的影响结果不同,海拔对茎叶性状影响随着构件指标不同而不同,但是不论海拔效应如何变化,植株大小对性状均存在显著影响(P<0.05)。
3 讨论
3.1 云冷杉当年生小枝功能性状的海拔效应讨论
植物性状可根据环境的异质性而改变,性状间存在一定关联性,在长期适应中形成了相互协调,共同进化的关系,由此,植物性状变异的程度可以反应植物对环境的适应[27]。本研究表明,海拔变化对云冷杉当年生小枝的总叶生物量、茎生物量、茎长、茎粗存在显著影响,表明当年生小枝对海拔变化积极响应,且除比茎长外,茎叶性状间存在极显著的正相关关系,这在前人研究中得到普遍证明。叶片是植物获取资源的重要场所,全叶生物量与茎生物量、茎长、茎粗间的正相关关系说明营养结构与支撑结构有较大的交互影响,枝条是支撑结构的重要部分,它不仅维持物质的运输还会通过其结构形成一个最优的光拦截体系,从而提高叶片生存能力[28]。优越的构型会提高养分获取的能力,获得的养分会在植物体内合理分配,从而促进茎构型的优化。营养结构与支撑结构的生长有着相互促进的正向关系。云杉低海拔处茎长可能是为了增加植物对光能的获取,而高海拔小枝变短可能与抵抗风力,减少低温、高光等恶劣环境带来的影响有关[29]。茎粗也在不同海拔上差异显著,最高海拔拥有最细的茎,可能与茎生物量分配较少有关。冷杉茎长与海拔变化无显著差异,茎粗呈现“V”型趋势,可能是高海拔处常绿植物为确保叶片水分运输的稳定,增加茎粗以减少冰冻所引起的栓塞。云杉茎叶生物量与海拔呈现极显著的负相关关系,说明随着海拔上升,云杉地上部分可利用资源获取减少。最高海拔处,云杉茎叶生物量与茎形态性状值均为最低,有可能是高海拔下云杉倾向于地下器官的投资,产生大量细根来获取养分[30]。冷杉总叶生物量与海拔间存在负相关关系,茎生物量与茎粗变化一致,在高海拔拥有较高茎生物量,说明冷杉通过提高茎生物量来抵御恶劣环境造成的影响。植物茎叶性状对气候变化的不同变异表征了植物不同的生长策略。
以茎长或小枝生物量作为为协变量,海拔作为固定因子,剩余性状作为因变量,用SPSS软件对性状指标进行分析;PL:植株大小(茎长)效应 Plant size (stem length) effect;PB:植株大小(小枝生物量)效应Plant size (twig biomass) effect
植物性状在低海拔处变异小于其他海拔处的变异,这可能是因为低海拔环境变异相对于高海拔也相对平缓[18]。种间性状的变异说明了面对相同环境不同植物采取适应策略的不同。种内云杉变异程度普遍大于冷杉,研究表明,高变异性状与植物存活、资源获取以及群落动态有重要作用[31],证实了岷江源区冷杉适应性较云杉强,这与Naudiyal等[22]对岷江源区亚高山森林研究得到的结论一致。
3.2 云冷杉当年生小枝生物量分配的海拔效应讨论
山体垂直带上,海拔变化综合影响着其他环境因子的组合及变异,相同海拔下植物茎叶分配的差异显示了不同功能型的响应策略。有研究表明,生物量分配与积累是植物净碳获取的重要驱动因素,生物量分配格局和不同器官性状生长关系权衡对植物个体生长具有重要意义[32]。叶片对光能和碳物质拦截有直接作用,为提高光合速率,植物会尽量提高叶物质的分配[33],但在高光和强风的环境下,植物为了避免恶劣环境下的机械损伤,增加支撑结构的物质分配[9]。本研究表明,云杉在低海拔处对茎的百分比分配显著高于较高海拔处,小枝水平上不同海拔茎生物量和总叶生物量低海拔呈现大于1的异速生长,在高海拔呈现等速生长关系。植物自身的生物分配量会在生长关系的约束与资源的限制下达到最优[34],即使植物在同速生长的情况下也会因为适应环境的改变而不断调整自身的生长方式,以维持自身功能的良好实现。这表明在不同海拔下云杉当年生小枝存在权衡关系,低海拔下茎生物量增加速率比叶生物量增加速率快,茎构型不断调整以维持养分获取能力,整体而言,茎对小枝内生物量分配具有重要影响。冷杉高海拔处的茎质量分配比高于低海拔,三个海拔上茎叶生物量呈现异速生长关系,印证了冷杉在高海拔通过增加茎生物量分配,积极调整茎构型来应对恶劣环境。
3.3 云冷杉不同适应策略的讨论
在本研究中,云杉小枝性状对海拔的变化响应策略与冷杉表现不同。方差分析结果表明,云杉在应对海拔梯度变化时除了本身小枝性状会发生变化,小枝大小也会随海拔变化而发生变化,从而影响小枝性状的表达。相较冷杉来说,无论是以茎长还是小枝生物量作为协变量来说明植株大小对小枝性状的影响,结果均表现出性状与植株大小间显著的相关性,说明冷杉主要通过改变当年生小枝的大小从而影响植物茎叶性状的表达。
异速生长分析发现,云杉低海拔处茎生物量积累速率大于总叶生物量,茎长增加率高于茎粗,高海拔为茎叶生物量增加及茎长茎粗转变为同速生长,说明云杉生长过程中存在生境胁迫,在低海拔处通过提高茎长来提高植物获得养分的能力,但是高海拔处生物量最低,代表胁迫仍未解除,为降低由于茎长而受到高海拔强风的机械损伤,茎长与茎粗间转为同速生长关系。云杉的分布格局主要受到气候的影响[35],前人研究紫果云杉对温度因子生态相对较窄[36],本研究样地不同海拔间环境因子数据显示(未发表)不同海拔空气温度、土壤养分及温度指标均对海拔变化有所响应,生长季高海拔空气温度最低,土壤温度中海拔与高海拔温度交替最低。通常寒冷地区温度的上升有助于打破植物的休眠,提高物质积累[16]。三个海拔云杉生长胁迫一直存在,且最高海拔胁迫可能最大,因此我们推断云杉胁迫因子为温度。随海拔的升高,冷杉茎叶生物量间和茎长、茎粗间生长关系保持异速生长,茎生物量累积和茎长生长较快,植物纵向的变化影响光能获取与种子扩散[36],茎长增大的同时需要径向生长以保持水分、养分的运输,同时增加机械支撑,保持稳定性[37],结合茎叶性状在三个海拔上的表现,说明冷杉主要通过调整茎构型与茎生物量来适应环境变化,茎性状在海拔上“ V ”形的趋势表明冷杉在高海拔生长策略的改变。冷杉属适应湿冷的环境,是松科原始的属,与云杉相比较,冷杉对水分要求更高,云杉比冷杉更耐干旱[35]。结合研究区环境因子的变化来看,冷杉可能在此海拔存在干旱胁迫[18]。
综合分析云杉与冷杉的生长策略及胁迫因子,判断在全球气候变化的情况下,温度的升高会促进紫果云杉的生长,13万年前,在间冰期后,随着温度的升高,高山地区种群向青藏高原东北部高海拔低温地区出现大规模的扩张现象,而当今气变环境下,会推动紫果云杉向低温地区进行扩张[37],这与李宁宁等[7]对青藏高原紫果云杉研究结果一致。冷杉胁迫因子为干旱[38],随着海拔的升高,干旱加剧。冷杉较云杉更易遭到生长季高温抑制作用[37],所以降水和温度同时主导岷江冷杉的分布。未来高海拔低温区可能会是冷杉的扩张目标,这与潘少安等[18]对四川省岷江冷杉研究结果、Liao等[39]对中国西南地区冷杉未来分布研究结论一致。
4 结论
云杉和冷杉当年生小枝性状在不同海拔间有显著的差异,不同植物相同性状表现出不同变化规律:云杉茎生物量,总叶生物量和比小枝长与海拔存在极显著相关关系,云杉性状的改变也受到小枝大小的影响;冷杉总叶生物量与海拔间存在显著负相关关系。此外,云杉与冷杉在不同海拔间的生物量分配及权衡关系也因海拔差异表现出不同的分配偏好;不同海拔下的相关生长分析说明典型高山乔木在不同海拔下呈现不同的生长策略,同时表明云杉与冷杉在不同异质环境下有较强可塑性,能够通过采用不同生长策略来缓解生长压力并实现资源最优配置。全球气候变化状态下,紫果云杉会向低温地区进行扩张,岷江冷杉则偏向于高海拔低温区。
