大陈岛礁海域甲壳类群落结构时空格局及其环境驱动
2023-12-25陈怡卉汪振华章守宇程晓鹏邹沁东
陈怡卉,汪振华,2,*,章守宇,2,程晓鹏,邹沁东,林 沅
1上海海洋大学海洋生态与环境学院,上海 201306
2上海海洋大学海洋牧场工程技术研究中心,上海 201306
近岸岛礁海域因其特殊的地理位置、极高的资源养护能力和独特的生物多样性维持功能,成为生产生活、科学研究和管理保护工作的重要区域[1—5]。该海域往往是连接淡水和外侧海洋环境的缓冲带,从生物圈层次上看,是从淡水向海洋过度的重要生态过度区。而从资源产出角度全面解构其系统状况,并揭示其功能特征和潜在的生态服务功能,为资源的合理利用和高效管理提供参考,是海洋科学研究的重要内容。因此,围绕岛礁海域资源格局的系统研究显得颇为重要。
环境与海洋动物群落分布之间的关系是生态学的一个基本问题。近年来,随着甲壳类群落生态学研究的不断深入,近岸、河口等浅水海域局域尺度的研究逐渐活跃,沿岸岛礁海域的相关研究也有一定积累[6—11]。尽管围绕岛礁海域甲壳类群落结构的研究越来越多,但岛礁系统这一特殊生境的多变水体环境、极高空间异质性是否造成群落结构的多元化这一问题认识不足。大陈岛礁海域复杂的流系、多样的底质和凹凸的地形,加之潮间带的影响以及镶嵌其中的各种养殖设施和海岸工程结构体,共同形成了空间异质性极高的栖息环境。研究认为,相比开阔海域,高异质性的岛礁海域其生物群落更易受栖息地及海底环境的动态变化所制约。为了验证上述科学问题,本文将从大陈岛礁海域甲壳类资源状况、群落组成的时空格局及其环境因子的集成驱动机制、群落稳态三方面进行分析,为海域渔业资源的保护和合理利用及海洋牧场的功能设计提供直接依据。
1 材料与方法
1.1 站点设置
围绕大陈岛上下主岛共设置21个站位(2020年9月为前20个站点),调查范围为28°22′—28°28′N,121°48′—122°00′E(图1)。研究海域主要位于10m等深线外侧,西侧水深小于10m,东、北、南侧水深大于10m,上、下大陈两主岛中间为水深10—30m的潮流槽。根据岛礁辐射范围,水平方向上将研究海域细划为岩礁区(2、3、8、9、13、21号)、近岩礁区(4、7、12、14、17、18号)、河口侧浅水区(1、10、11、16号)和外侧深水区(5、6、15、19、20号)。2020年9月(夏季)、11月(秋季)、2021年1月(冬季)、4月(春季)连续开展4个季度的渔业资源底拖网调查。

图1 大陈岛礁海域调查区域及采样布设
1.2 采样方法
甲壳类样本皆采用单船网板底拖网采集,网板类型为V型铸铁。租用当地拖网渔船浙椒渔15050协助采样。网具全长36m,其中网袖长度16m,目大6cm+7cm+16cm(袖口);网身长度18m,目大3.5cm+4.5cm+5.5cm(与网袖连接处),网囊长度2m,目大为2cm。拖曳时网口平均有效扩张高度和宽度分别约为1.5m和5m,曳纲在水深为25m以内时设置长度为100m,水深大于25m时长度设置成150m,平均拖速为3.0kt。2020年夏季渔获物较多时拖曳时间设置为15min,其余航次皆为20min。采样和实验环节皆依据《海洋调查规范—海洋生态调查指南》(GB17263.9, 2007)进行。各站位温盐深等水文环境因子采用温盐深仪监测同步测定(对比后采用合理数据进行分析)。对每站所得渔获物进行编号、记录和冰鲜保存,带回下大陈临时实验场地进行分类与生物学测量,重量精确到0.01g。甲壳类鉴定主要依据《浙江动物志(甲壳类)》和《东海经济虾蟹类渔业生物学》等工具资料,并对不同物种进行大、中、小型分类,分类依据为优势组体长/甲长和体重,具体范围详见表1。

表1 大中小型甲壳类的分类依据
1.3 数据处理
采用Pinkas[12]相对重要性指数(IRI)判定群落优势种:
IRI=(N+W)×F×104
式中,N为某物种的个数占总数的百分比,W为某物种的重量占总重量的百分比,F为某物种出现的站次数占调查总站次数的百分比。IRI>1000,物种为优势种;100—1000,重要种;IRI<100,少见种。
各测站拖网渔获物资源密度的估算采用扫海面积法[13],计算式为:
ρi=Ci/aiq
式中,ρi为第i站的资源密度(重量:kg/km2;尾数:尾/km2);Ci为第i站的每小时拖网渔获量(重量:kg/h;尾数:尾/h);ai为第i站的网具每小时扫海面积(km2/h)(网口水平扩张宽度(km)×拖曳距离(km),拖曳距离为拖网速度(km/h)和实际拖网时间(h)的乘积;q为网具捕获率(可捕系数,=1-逃逸率),取0.5。调查海域资源密度取各站点拖网渔获物密度的算术平均数。采用单因素方差分析(ANOVA)和最小显著差法(LSD)对季节间和区域间资源密度进行差异性检验,显著性水平为α=0.05。
通过置换多元方差分析(PERMANOVA)探索群落结构差异的潜在来源[14—15]。基于丰度数据构建Bray-Curtis相似性矩阵,数据使用之前进行标准化处理(皆统一为每小时的渔获数量)和四次方根转化[16]。采用非度量多维标度排序(NMDS)和等级聚类(Cluster),分析甲壳类群时空格局[17—18],同时结合丰度生物量(ABC)曲线及W统计量进行群落扰动和变化分析[18]。NMDS二维结果图的可用性用胁迫系数(Stress)来衡量,Stress<0.2时,表示其图形有一定解释意义[19]。相似性百分比(SIMPER)用来判别造成群落组间差异性和组内相似性贡献率较大的关键生物。相似性分析(ANOSIM)用于检验生物组合之间差异的统计学意义,置换次数为999。
冗余分析(RDA)用于分析生物群落与环境因子的相关性。对物种数据进行去趋势对应分析(DCA),根据每个轴梯度长度(LGA)选择最佳排序方法[20]。根据DCA分析结果,本研究选择冗余分析。原始环境数据进行标准化处理。采用R4.0.4的vegan包完成,并用rdacca.hp包进行层次分割获取单个解释变量的贡献[21]。
站点图、资源密度分布图使用ArcGIS10.8绘制,群落结构的多元分析基于PRIMER7.0实现,数据统计在SPSS26中完成。
2 结果与分析
2.1 环境特征
大陈岛水体环境季节变动明显。底层水温(BT)、底层溶氧(BDO)在季节间呈现极显著差异(P=0.000)。夏季的底层盐度(BS)远低于冬春季(P=0.000,P=0.003)。春季的底层海水叶绿素(BChl.a)显著高于夏季(P=0.03),极显著高于秋冬季(P=0.000)。夏季无机氮(DIN)极显著低于其他三季(P<0.01)(表2)。

表2 不同季节环境因子统计结果
水平方向上,外侧深水区与其他区域形成清晰划分,具有局域相对高盐、低氧特征。外侧深水区的盐度显著高于其他区域(P<0.05),底层溶氧极显著低于其他区域(P<0.01)。外侧深水区底温在夏季显著低于其他区(P<0.05),冬季极显著高于其他三区(P<0.01)。春季的叶绿素含量在近岩礁区显著高于其他区域(P<0.05),夏季外侧深水区显著低于其他区域(P<0.05)。营养盐方面,冬季的无机氮在岩礁区极显著低于其他区域,春季河口侧浅水区极显著高于其他区;冬季的活性磷酸盐在外侧深水区极显著高于最低区域近岩礁区(P=0.005),春季河口侧浅水区显著高于其他区域(P<0.05)。
水深主导的垂直方向上,底温、底盐以及底层溶氧以10m为深度梯度出现季节性差异。春夏季,随水深的增加底温下降,在20—30m与1—20m形成显著差异(P<0.05),秋冬季变化趋势相反,10—20m底温较高,但仅冬季有统计学差异(P冬<0.05);底盐则始终保持20—30m显著高于10—20m(P<0.05);底层溶氧随水深降低,但在春季无水深间显著差异(P夏秋冬<0.05);春季的最主要差异体现在叶绿素含量,20—30m初级生产力明显较高(P<0.01)。
2.2 种类组成及优势种更替
结果共采集甲壳类53种,隶属2目13科29属(表3)。夏季共捕获甲壳类26种,秋、冬、春季为43、41、31种。四季均出现物种19种,占全年总种类数的35.85%,包括重要经济种哈氏仿对虾、三疣梭子蟹、口虾蛄等。单一季节分布物种15种。区域间物种组成差异明显,岩礁区全年采集物种40种,近岩礁区、河口侧浅水区、外侧深水区各为47、31、38种,且秋冬春三季,近岩礁区物种数均为最丰富,河口侧浅水区种类数明显低于其他三区。

表3 大陈岛礁海域甲壳类物种名录
夏季优势种有6种,其中哈氏仿对虾和刀额仿对虾优势度不持续,为当季优势种。秋季优势种中三疣梭子蟹、断脊小口虾蛄、中华管鞭虾优势度不再持续,为暖水季优势种。冬季优势种减少为4种,双斑蟳优势度突出,与细巧仿对虾,日本蟳共同构成冷水季优势种。春季,日本鼓虾为当季优势种(表4)。夏春、夏冬、秋冬、秋春季间优势种种类更替率分别为0.90、0.89、0.71、0.75,夏秋、冬春季优势种较重叠,更替率分别为0.43和0.20。综上,大陈海域夏秋季节优势种以经济种为主,冬春季节小型个体取而代之。口虾蛄为全年优势种,优势度突出。

表4 大陈岛海域甲壳动物优势种组成
2.3 资源量的季节变动与空间分布
甲壳类资源密度呈现出秋季>夏季>冬季>春季节的季节变化特征。丰度的季节间差异极显著(F=5.111,P=0.003),秋季丰度约为冬季的1.3倍(P=0.006),春季的1.7倍(P=0.003);夏季丰度约为春季1.6倍(P=0.009),也较高于冬季(P=0.017)。生物量同样具有季节间极显著差异(F=6.731,P=0.000)。
水平方向上,甲壳类资源分布不均(图2、3)。夏季,近岩礁区资源密度高,外侧深水区低,但无显著差异。秋季,外侧深水区丰度最高,河口侧浅水区最低(P=0.028)。冬季,近岩礁区、外侧深水区丰度显著高于岩礁区、河口侧浅水区(P<0.05),外侧深水区生物量极显著高于岩礁区与河口侧浅水区(P=0.001),近岩礁区生物量显著高于岩礁区与河口侧浅水区(P=0.016,P=0.012)。春季,丰度在近岩礁区最高,生物量在外侧深水区最高,河口侧浅水区依然为资源量最低,且外侧深水区生物量远高于其他三区(P<0.01)。

图2 大陈岛甲壳类丰度的空间分布
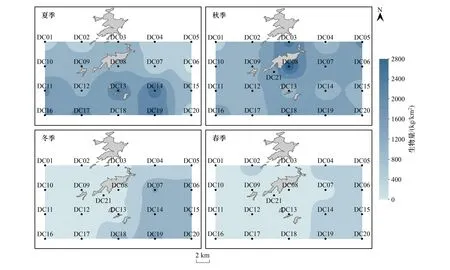
图3 大陈岛甲壳类生物量的空间分布
垂直方向上,甲壳类丰度和生物量在全年时间范围内均随水深的增加而增加,但各水深间无显著差异(P>0.05),Pearson分析显示,水深与丰度之间存在很弱的正相关(R=0.26,P=0.02);而以季节做时间范围,冬季的资源量以5m为水深梯度即呈现显著差异:25—30m的资源量显著高于15—20m、10—15m(P<0.05),20—25m显著高于10—15m(P<0.05);此时做Pearson分析,丰度与生物量均与水深存在中等程度正相关(R丰度=0.60,P=0.004;R生物量=0.65,P=0.002)。需要注意的是,夏季资源量与水深的增减关系逆转,资源量随水深增加而下降。
2.4 群落结构的时空差异
2.4.1群落差异潜在来源
PERMANOVA分析结果显示,“季节”和“区域”因子在0.01水平对群落结构产生影响,两因子的交互作用也在0.01水平发挥作用,而“水深”的影响在0.05水平(表5)。“季节”的方差解释度为38.65%,远大于其他因子。群落结构变动受“季节”因子的强烈影响,而“区域”对群落的作用具有“季节”依赖性。
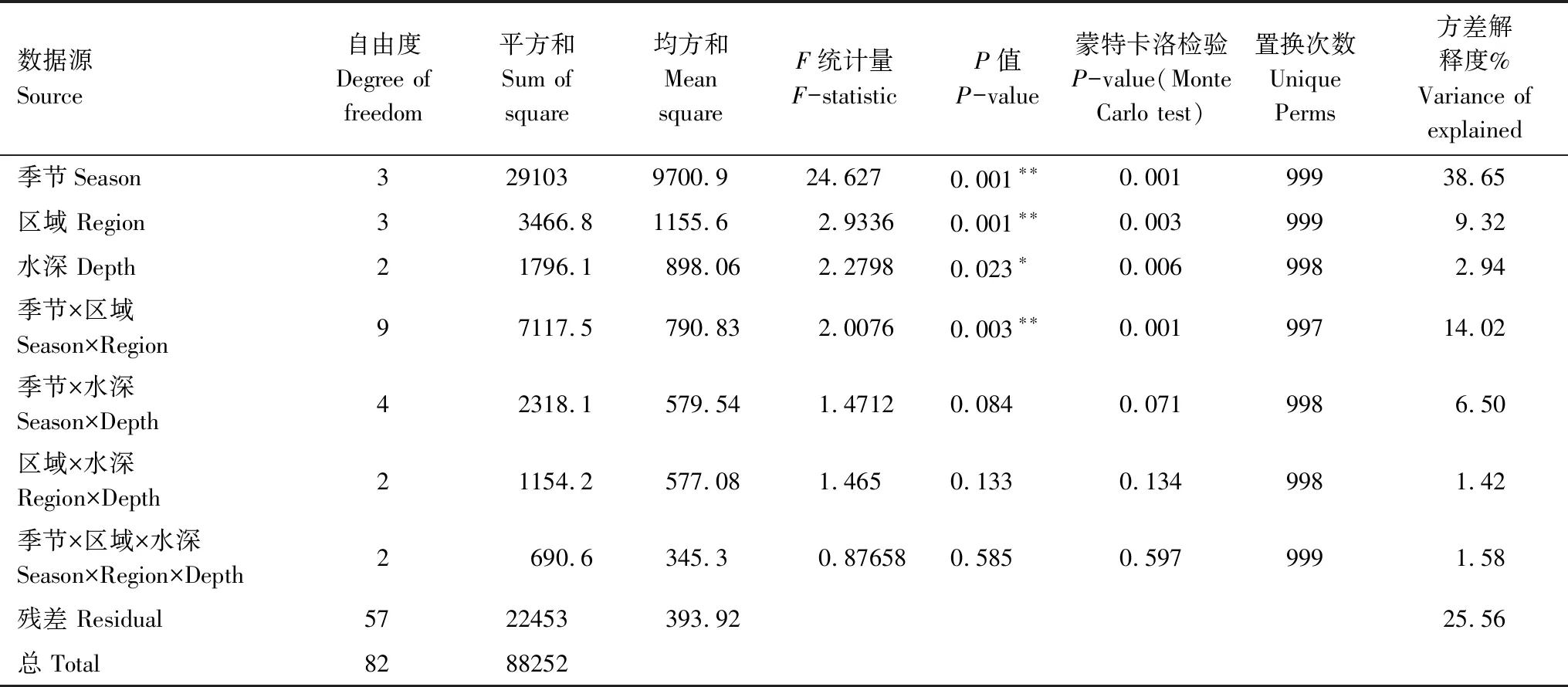
表5 整体丰度数据集的置换多元方差分析检验结果
2.4.2群落结构季节变化
全年丰度数据集聚类结果显示,60%相似性下夏季与秋季群落完全分离,群落结构特异性强,而冬季和春季群落在65%相似性附近部分分离(图4),存在明显交叉,群落结构具有相似性。NMDS在胁迫系数为0.15下显示相似结果,具有一定解释度(图5)。ANOSIM全局检验,季节间群落差异极显著(R=0.698,P=0.001)。

图4 大陈岛海域甲壳类群落全年聚类分析

图5 大陈岛海域甲壳类群落全年NMDS排序图
群落相似性分析结果显示(表6),各季群落的组内相似性范围为63.36%—70.96%,特征种季节变更明显,多为优势种。夏季特征种全部为优势种;秋季在优势种基础上新增5个特征种,其中,优势种口虾蛄、中华管鞭虾、断脊小口虾蛄为贡献率前三;冬季新增5种,优势种口虾蛄、细巧仿对虾以及日本鼓虾为贡献率前三;春季新增4种,贡献率前三均为春季优势种。口虾蛄是四个季节唯一的共同特征种。群落相异性分析结果显示(表6),夏冬两季间群落差异性最高,相异性系数为62.24%,其次为夏春。夏冬季主要分歧种为哈氏仿对虾、细巧仿对虾,刀额仿对虾等;夏春季主要分歧种为日本鼓虾、刀额仿对虾、哈氏仿对虾、双斑蟳和细巧仿对虾;秋冬季主要分歧种为断脊小口虾蛄、中华管鞭虾、双斑蟳、安氏白虾;夏秋季主要分歧种为细巧仿对虾、双斑蟳、刀额仿对虾等、三疣梭子蟹等;秋春两季主要分歧种为断脊小口虾蛄、中华管鞭虾、双斑蟳、日本鼓虾等;冬春季主要分歧种为安氏白虾、双斑蟳、中国毛虾等。

表6 大陈岛海域甲壳类各季节群落相似性与群落相异性贡献种(70%)
2.4.3群落结构空间差异
水平方向上,夏季群落在四个区域未发生完全分离(图6),但ANOSIM检验显示外侧深水区群落与岩礁区和近岩礁区具有显著差异(RD-R=0.336,P=0.048;RD-N=0.379,P=0.017),造成群落区域差异的典型分歧种为中华管鞭虾、中国毛虾、扁足异对虾、三疣梭子蟹等。秋季(图6),外侧深水区呈现单独聚集状态,群组Ⅱ则主要由河口侧浅水区站点组成,ANOSIM显示外侧深水区群落与河口侧浅水区、近岩礁区呈极显著差异(RD-S=0.9,P=0.008;RD-N=0.472,P=0.009),与岩礁区呈显著差异(RD-R=0.459,P=0.015),同时,河口侧浅水区群落与岩礁区和近岩礁区也呈现显著性差异(RS-R=0.611,P=0.01;RS-N=0.353,P=0.033);河口侧浅水区群落的典型分歧种为扁足异对虾、双斑蟳、日本鼓虾等,外侧深水区群落典型分歧种则为双斑蟳、葛氏长臂虾、红星梭子蟹等。冬季(图6),区域分化不清晰,但外侧深水区站点单独聚集,ANOSIM显示外侧深水区与其他区域群落分化极显著(RD-S=1,P=0.008;RD-N=0.675,P=0.004;RD-R=0.8,P=0.002),河口侧浅水区与近岩礁区分化显著(RS-N=0.456,P=0.029);外侧深水区群落的典型分歧种为双斑蟳、安氏白虾、日本蟳等,而河口侧浅水区与近岩礁区群落分化的典型分歧种为双斑蟳、脊尾白虾、中华管鞭虾等。春季(图6),河口侧浅水区站点主要集中在群组Ⅱ,ANOSIM显示河口侧浅水区群落与近岩礁区和外侧深水区具有显著性差异(RS-D=0.875,P=0.029;RS-N=0.373,P=0.038),典型分歧种为中国毛虾、隆线强蟹、三疣梭子蟹、断脊小口虾蛄等。
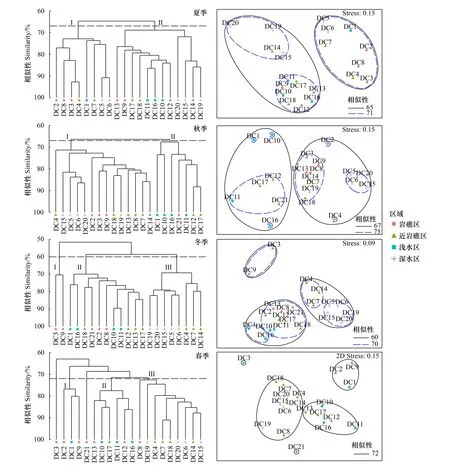
图6 大陈岛海域甲壳类群落季节聚类分析与排序
垂直方向上,除秋季外的群落,均以10m做水深划分,产生一定程度分化,ANOSIM检验显示,10—20m群落与20—30m群落差异显著(R夏季=0.312,P=0.009;R冬季=0.418,P=0.001;R春季=0.359,P=0.002)。夏季,两个水深群落的组间平均差异性为32.36%,SIMPER结果表明:三疣梭子蟹、红星梭子蟹和日本蟳在10—20m水层的平均丰度较高,而中国毛虾、中华管鞭虾、扁足异对虾和细巧仿对虾则在20—30m的平均丰度较高,断脊小口虾蛄在10—20m的平均丰度几乎是20—30m的两倍。冬季,两群落的组间平均差异性为40.66%,日本蟳和安氏白虾主要分布在10—20m,而口虾蛄集中分布在20—30m。春季,两群落的组间平均差异性为31.13%,中国毛虾、细螯虾和隆线强蟹在10—20m的平均丰度是20—30m的近乎3倍,断脊小口虾蛄则在20—30m的平均丰度是10—20m的几乎6倍。
2.5 环境因子对甲壳类群落的影响
去趋势对应分析结果表明,本研究适用基于线性模型的冗余分析。蒙特卡洛显著性检验结果显示,全年与夏秋冬三季的第一排序轴均达到极显著水平(P=0.001)。
全年丰度数据的RDA研究发现,环境因子矩阵共解释53.19%的生物群落季节变化,其中仅pH和活性磷酸盐对群落的变化没有显著性影响(P>0.05)(图7)。层次分割获取单个解释变量的贡献结果显示,底温和底层溶氧贡献远高于其他变量,分别为30.19%和30.25%。以轴1为界,夏秋季群落沿轴正向分布,冬春季群落则负向分布,而轴1与底温和底溶氧显著相关;同时,秋季主要沿轴2负向分布,其他季节仅外侧深水区沿轴2负向分布,轴2相关性较高的因子为水深、底盐等。夏秋季代表生物为中华管鞭虾、哈氏仿对虾、刀额仿对虾、锈斑蟳等,均与底温正相关,刀额仿对虾与哈氏仿对虾与底温的相关性尤为强烈。冬春季群落代表种类包括双斑蟳、细巧仿对虾、日本鼓虾、安氏白虾、中国毛虾等;双斑蟳与水深、底盐和无机氮正相关,且与其他物种相较与深度的相关性强烈,日本鼓虾、细巧仿对虾与底盐、无机氮和底层溶氧正相关。
夏季(图8),环境因子共解释72.29%的生物群落空间差异,仅底层溶氧和无机氮对群落结构的影响无统计学意义(P=0.496,P=0.094),单个解释量贡献值前三为活性磷酸盐(21.86%)、底盐(14.87%)、和底层叶绿素(12.55%)。外侧深水区分布在轴2的负方向,代表生物为中国毛虾、矛形梭子蟹、中华管鞭虾和扁足异对虾等,与水深和底盐正相关;其他区域主要分布在轴2正方向,代表生物中刀额仿对虾、口虾蛄、鲜明鼓虾等与活性磷酸盐、底温相关性强烈,三疣梭子蟹、日本蟳、红星梭子蟹等则与叶绿素和pH正相关。秋季(图8),群落空间差异的解释率为61.07%,底盐产生极显著影响(P=0.002)、水深和空间异质性具有显著影响(P=0.012,P=0.029),单个解释量贡献值分别为48.15%、20.72%、11.23%。外侧深水区沿轴1负向分布,代表生物为双斑蟳、日本鼓虾、刀额仿对虾、红星梭子蟹等,与底盐和水深正相关;河口侧浅水区沿轴1正向分布,代表生物为断脊小口虾蛄、周氏新对虾、葛氏长臂虾、日本蟳等,与空间异质性正相关。冬季(图8),群落空间差异的解释率为72.10%,底温、水深具有极显著性影响(P=0.003,P=0.001),无机氮和活性磷酸盐具有显著性影响(P=0.038,P=0.043),单个解释量贡献值分别为20.77%、14.73%、4.85%、3.10%。外侧深水区分布在轴1正方向,代表种为双斑蟳、周氏新对虾、中华管鞭虾等,与水深、底温正相关;河口侧浅水区沿轴1负方向分布,代表种为安氏白虾、脊尾白虾、矛形梭子蟹,主要与叶绿素正相关。春季(图8),共解释42.13%的群落空间差异,各环境因子的影响均不具有统计学意义。

图8 四个季节甲壳类群落与环境因子的冗余分析
2.6 群落稳态变化
四个季节的ABC曲线均呈现前段生物量曲线在上,丰度曲线在下,后段交叠的模式,W值均为正,秋、春季相较于夏、冬季,生物量曲线在更高位上变化(图9)。各季节丰度和生物量曲线累积优势度的前5位基本同时由当季优势种占据,而全年优势种口虾蛄在四季一直保持着第一生物量优势种的绝对地位,从而造就生物量曲线始终高位发展的态势(表7)。

表7 各区域不同季节丰度和生物量占比前5的种类

图9 甲壳类群落四个季节的丰度生物量曲线
不同区域的ABC曲线总体上依然表现为生物量曲线在上的模式,除去冬季的岩礁区,W值均为正(图10)。夏季,岩礁区和近岩礁区生物量曲线与丰度曲线相伴上升,差距不大,而河口侧浅水区和外侧深水区生物量在高位发展;秋季,ABC曲线的区域差异刚好与夏季相反;冬季,岩礁区的ABC曲线模式逆转为丰度在上,而其他三区依然为生物量在上;春季,近岩礁区和外侧深水区生物量曲线相对高位发展(表7)。
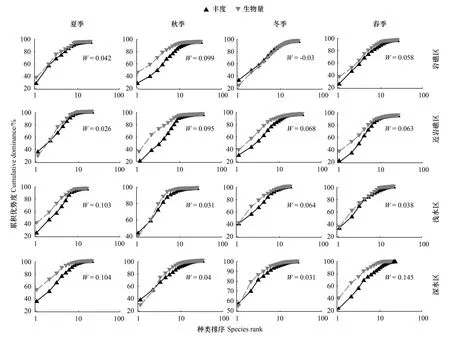
图10 甲壳类群落四个区域的丰度生物量曲线
3 讨论
3.1 大陈岛礁海域甲壳类组成与优势种更替
大陈岛礁海域采集的甲壳类以广温广盐种类为主。与长江口北支水域比较[10],共有物种18种,与南麂列岛附近海域共有种32种[22],种类组成基本属于东海沿岸广泛分布种[23]。从生态类型看,有72%为广温广盐类群。同步监测的环境数据显示,底层水温变化范围为10.83—26.81℃,底层盐度变化范围为28.38‰—31.34‰,年内变化范围较大,因此更适宜耐受性较强的广温广盐物种。周年及季节优势种均属广温广盐生态类群,因此,广温广盐种对大陈岛甲壳类群落变化起绝对主导。
本研究4个航次共采集甲壳类53种,夏季26种、秋季43种、冬季41种,春季31种。与朱玉丹等[9]在大陈洋产卵场开展的调查相比(38种),秋季物种数明显(26种)较多,与椒江口调查记录的26个物种(春季21种,秋季20种)相较[24],丰富性更为明显。上述调查海域与本研究有重叠,调查范围相对更广,而站点设置少,皆为影响捕获物种数的因素之一,但更值得关注的是岛礁系统生境类型多样化问题。大陈洋产卵场海域开阔,比较而言生境类型相对单一,椒江口靠近沿岸低盐水,温盐范围相对较窄,且采样站点图显示,多数站点远离岛礁。因此,对于营底栖生活的甲壳类而言,岛屿效应是造成本次调查物种繁多的重要原因。本研究中的岛礁生境调查结果与大陈岛开阔海域的现存记录形成鲜明对比,直观的体现出岛礁生境的空间高异质性是物种多样性的重要支撑。徐兆礼等[25]在瓯江口对十足目和口足目虾类的调查研究中指出,岛礁海域及其外部海域,往往是种类较多的水域,齐海明等[24]通过对比分析椒江口、瓯江口、东海大陆架甲壳类研究结果后强调了岛礁附近海域是甲壳动物资源保护的重点关注区域,这些观点与本调查结果不谋而合,因此,基于实际提出,岛礁海域可以支撑起更为丰富的甲壳类多样性。
大陈岛优势种组成与乐清湾、瓯江口共有种6种[26—27],属于浙江中南部海域优势种组成[28]。优势种季节更替明显,表现为夏秋季以哈氏仿对虾、三疣梭子蟹、中华管鞭虾等中大型个体的经济种为群落主导,冬春季则以双斑蟳、细巧仿对虾、日本鼓虾这类小型个体的饵料生物占主要地位。其优势种基本为洄游性物种,故洄游物种的的繁殖、索饵及迁移习性是造成该海域优势种季节性更替的重要原因。洄游性物种不同生命阶段对水温、盐度有严格要求,产卵和幼体发育阶段要求较低盐度。根据宋海棠等[23]的研究,哈氏仿对虾、刀额仿对虾在东海产卵季节为夏季(6—8月),幼虾密集分布在30m水深以浅的沿岸低盐海域索饵成长,并随个体长大逐渐向外侧海区移动,秋季在30—60m水深海域分布密度较高,冬季向东可分布到100m水深海域越冬。
调查海域水深范围在30m以浅,由此推测,春季水温回暖,哈氏仿对虾、刀额仿对虾产卵洄游进入该海域,于夏季在此聚集形成庞大的繁殖群体,产卵结束进入秋季,随水温下降索饵群体逐渐游离进入30m以深海域,而原繁殖群体由于能量消耗造成大量个体死亡,使得种群数量急速下降,故在秋季优势度大幅降低,进入常见种划分范畴,最终成为该海域夏季的特有优势种;比较而言,中华管鞭虾繁殖期相对较晚、较长,从夏季开始维持至秋季(7—10月),故其繁殖群体在夏秋两季于调查海域大量存在,形成两季优势种,同时,索饵群体的游离、繁殖群体中部分亲虾的死亡,使得秋季优势度低于夏季;调查海域也是三疣梭子蟹的重要产卵场,但与虾类不同的是,三疣梭子蟹可越过1到3个冬天,因此其优势度反而在秋季有所上升。冬季,水温转凉,洄游性物种离开该海域,生态位空出,由小型饵料物种及定居性物种接替补充,成为冷水季节优势种。外部人为干扰也是造成冬春季优势种个体小型化的重要因素。由于伏季休渔管控,主要捕捞季节在夏末至冬初,渔汛期间个体较大者被捕捞,从而加剧冬春季节优势种个体小型化。另外,口虾蛄在全年时间范围内保持着资源量的压倒性优势,较其他优势种的相对重要性指数高出不少。口虾蛄为多年生甲壳类,喜好穴居,游泳能力强,是凶猛的捕食肉食性动物,食性较广,以鱼虾贝类为食,耐氧能力强,这些生物学特征奠定其资源竞争能力[29]。
3.2 大陈岛礁海域甲壳类资源量的变化特征
大陈岛甲壳类资源量呈现夏秋季显著高于冬春季的状态。各季优势种对当季丰度贡献率的平均值为71.14%,生物量贡献率的平均值为81.86%,优势种的季节更替牵连资源量的季节变化。该海域是多数物种的产卵场与索饵场[23—24],春季物种多样性得以补充恢复,洄游性物种经过繁殖育幼使得种群数量增大,成为夏秋季节的主要捕捞量;同时,渔业生产的相关因素也是导致群落结构和数量变化的重要因素。物种的生活习性也会造成资源量空间格局上的差异。夏季,外侧深水区是资源量的低洼,此时,优势种由繁殖群体和幼体构成,由于对低盐环境的依赖,主要分布在沿岸河口侧浅水区和岩礁附近。秋季开始,索饵群体逐渐向外侧海域迁移进行越冬洄游,调查海域资源量高值区逐渐向外侧深水区移动,河口侧浅水区成为秋冬春三季的资源低洼。垂向的资源分布印证了相同规律,夏季资源量在盐度较低的10—20m较高,结合SIMPER分析结果可知,在此水深范围平均丰度较高的物种是三疣梭子蟹、红星梭子蟹等洄游性蟹类,而夏季正是其产卵季节,繁殖群体在此水深范围内更加聚集。夏季结束,资源量向20—30m倾斜,也正是由于部分繁殖群体的消亡和索饵群体的移动。
3.3 大陈岛礁海域甲壳类群落结构时空格局及其环境因子的集成驱动机制
大陈岛甲壳类群落随水体环境的季节变化既呈现波动特征,又表现出稳定状况。聚类和排序结果显示,夏季、秋季群落各自单独聚集,特异性强,冬春两季群落呈现交叉现象,具有一定相似性。时间格局下的群落结构差异由群落内部生物效应和外部环境非生物效应共同影响。研究认为,优势种繁殖和索饵习性的差异,是导致群落季节异质特征的主要生物因素[30—31]。SIMPER进一步指出,各季节优势种同时扮演着造成季节群落分离的主要分歧种。造成夏秋季群落分离的主要分歧种为细巧仿对虾、双斑蟳和刀额仿对虾等,刀额仿对虾是夏季特有优势种,而双斑蟳在夏季仅为常见种。冬春季共有优势种四种,故两季群落结构相似,季节分界模糊。优势种作为群落中的关键因子,对于维持群落结构稳定性具有重要意义。另一方面,物种组成结构差异与水文环境的季节变化具有十分重要的联系[32—35]。RDA显示底温、底盐、底溶氧等多个环境参数能够解释53.19%的群落季节变化,底温和底溶氧的单个解释量远高于其他参数。夏秋季底温较高,成为影响生物群落的关键因子,三疣梭子蟹、中华管鞭虾、哈氏仿对虾等优势种对水温的需求较高,这是因为该时期的甲壳类群体以幼体为主,对低温的耐受程度低[23,],因而依赖于较高水温,同时需要低盐环境。冬春季底层溶氧上升,生物群体对溶氧有更高的要求,同时,相较于夏秋季,盐度和叶绿素的影响逐渐增大,重要优势种双斑蟳对盐度有绝对的需求,日本鼓虾和细巧仿对虾对盐度和溶氧都具有较高要求。此外,海底流场也是生物行为的潜在影响因素,其具体机制需要进一步深入探讨。
岛礁生境因其多样的底质和复杂的地形更具有空间异质性,PERMANOVA结果表明,区域对群落结构有显著影响,甲壳类群落也表现出不同区域的结构差异。夏季,外侧深水区群落结构差异突出,中国毛虾、扁足异对虾、细巧仿对虾、矛形梭子蟹等小型个体的分歧种在外侧深水区的平均丰度明显较高,RDA指出,这些物种更倾向于水深相对较深、盐度相对较高的生境;第一分歧种中华管鞭虾也有此生境选择倾向;而重要经济种三疣梭子蟹则表现出近岩礁区平均丰度最高特征,似乎更依恋于空间异质性较高的礁石区。秋季之后,双斑蟳的优势度突出,成为典型优势种,此后造成外侧深水区群落与其他区域群落异化的第一贡献种持续为双斑蟳,其平均丰度在外侧深水区明显高于其他区域,这与该物种对生境环境的物理需求有关,RDA显示双斑蟳强烈依恋于较高的盐度和水深,而双斑蟳同时作为河口侧浅水区群落的分歧种,在河口侧浅水区的平均丰度明显较低,这一现象与RDA揭示的信息相吻合。双斑蟳对生境的定向选择,是秋、冬两季外侧深水区、河口侧浅水区群落同时显著异化的重要原因。另外,安氏白虾、脊尾白虾等河口种在外侧深水区的平均丰度为0,进一步加剧外侧深水区与河口侧浅水区的群落空间分化。春季,河口侧浅水区群落的单一分化,与中国毛虾、隆线强蟹、细螯虾、巨指长臂虾等饵料种在河口侧浅水区的高平均丰度有关。
水深与生境类型的转换相关联,生物对生境的选择自然会在垂直方向表现出变化。在研究的局地尺度下,水深依然能引起群落的细微差异,例如夏季三疣梭子蟹、红星梭子蟹和日本蟳在10—20m有更多分布,而中华管鞭虾、细巧仿对虾相反;虾蟹类分布水深的差异和其主要习性密切相关,蟹类喜欢硬相岩礁生境,以便躲藏或抵抗强流,虾类喜欢软相泥地,“虾米吃稀泥”,小粒径的软泥方便其滤食。岛礁海域的生境复杂性,使得甲壳类群落更易受栖息地及海底环境的动态变化所制约,从而展现出时空交互的动态格局。
3.4 大陈岛礁海域甲壳类群落稳态特征
尽管不同季节的ABC曲线特征不尽相同,但W统计值均为正值,说明大陈岛礁海域甲壳类群落抵抗捕捞干扰、污染和栖息地破坏的能力处于较高水平,群落较稳定。
ABC曲线的不同特征主要由不同物种的生物量和丰度的比例引起,且优势种的体型大小也对丰度曲线和生物量曲线的位置有一定影响,ABC曲线特征可在一定程度上反映群落中物种体型的相对大小[36]。研究海域四个季度的生物量优势度曲线均位于丰度优势度曲线之上,说明甲壳类群落以大中型种类为主导,且口虾蛄在全年具有绝对的生物量优势。但从口虾蛄在冬季岩礁区的生物量占比下降导致其ABC曲线特征逆转,W值出现负值这一现象分析,本研究中ABC曲线在各季节各区域基本呈现生物量占优势这一信息,似乎并不能完全代表该海域的甲壳类群落状态,猜测口虾蛄的绝对主导作用可能掩盖了甲壳类群落的真实情况,从前期阐述的优势种的季节更替来看,十足目虾蟹类确实存在夏秋季以中型个体的经济种为主,向冬春季以小型的饵料种转变的事实。因此,姑且只能认为口虾蛄在该岛礁海域的资源量充足且稳定。
在阐述群落组成格局的基础上,结合ABC曲线的区域差异,可以进一步揭示河口侧浅水区和外侧深水区在物种组成比例上有明显差异。中华管鞭虾是外侧深水区的第一丰度贡献种,生物量的2位,中国毛虾是丰度第3位,而在其他区域的丰度占比非常低,蟹类中双斑蟳在外侧深水区的丰度和生物量都最大,说明这些物种对外侧深水区环境确实存在选择倾向。同样,进入冬季,季节性洄游种退出生态位竞争后,安氏白虾、脊尾白虾等河口性种在河口侧浅水区的聚集就显现出来。
4 结论
本文通过对大陈岛礁海域甲壳类群落结构的研究,得出以下结论:
(1)岛礁生态系统可以支撑丰富的甲壳类物种,优势种全年更替,资源量充沛。
(2)大陈岛礁海域优势种多为洄游性种,夏秋季群落以哈氏仿对虾、中华管鞭虾、三疣梭子蟹等中大型经济种主导,冬春季则转变为小型饵料种占优势,同时口虾蛄周年兴盛不衰。
(3)大陈岛礁海域甲壳类群落结构具有明显季节变化,夏季与秋季群落异质性高,冬春两季群落相似,底温、底盐和底层溶氧是造成群落季节波动的关键环境因子,优势种繁殖索饵习性发挥着重要生物效应。
(4)生境的空间异质性和环境因子的区域差异导致岛礁系统内甲壳类群落产生区域分化,河口侧浅水区和外侧深水区的群落结构差异尤为突出,中华管鞭虾、双斑蟳对盐度较高的外侧深水区生境具有明显选择倾向,而安氏白虾、脊尾白虾等河口种是河口侧浅水区低盐群落的特征种。
(5)大陈岛礁海域甲壳类群落全年较稳定,具有较高的抗干扰能力。

附录1 物种编号
