T淋巴细胞在原发性硬化性胆管炎中的作用
2023-12-18李子怡张万洁王富春毛小荣2a李俊峰2a2b
李子怡, 张万洁, 王富春, 毛小荣,2a, 李俊峰,2a,2b
1 兰州大学第一临床医学院, 兰州 730000; 2 兰州大学第一医院 a. 感染科, b. 感染科与传染病研究室,兰州 730000
原发性硬化性胆管炎(PSC)是一种免疫介导的慢性胆汁淤积性肝病,其特征是肝内外胆管炎症及纤维化导致多灶性胆管狭窄、慢性胆汁淤积,可进展为肝硬化及终末期肝病,且患胆管癌的风险增加[1]。目前认为,PSC 的发病可能与遗传、环境、免疫、肠道微生态及胆汁酸代谢等因素相关,但具体发病机制尚不明确。在遗传学研究上,PSC 与人类白细胞抗原的显著关联表明,其与适应性免疫反应密切相关。且研究[2]发现,在PSC 患者的肝活检中,汇管区炎症浸润的主要细胞类型是T 淋巴细胞。因此,T 淋巴细胞介导的免疫应答在PSC发病中起着重要作用。
T 淋巴细胞来源于骨髓中的多能干细胞,在胸腺中发育为成熟T 淋巴细胞,再随血液进入外周淋巴器官,接受抗原刺激,发生免疫应答。根据其抗原识别受体(T cell receptor,TCR)的多样性,T 淋巴细胞可被分为αβ T 淋巴细胞和γδ T 淋巴细胞。除了基于TCR的分类,T 淋巴细胞还可根据细胞表面共受体分为CD4 和CD8 两种亚群。这些共受体可通过增强TCR与主要组织相容性复合体的结合来促进T淋巴细胞的活化。除了表达TCR αβ的常规T淋巴细胞外,γδ T淋巴细胞、自然杀伤T淋巴细胞(nature killer T cell,NKT)和黏膜相关不变T 淋巴细胞(mucosal-associated invariant T cell,MAIT)被称为非常规T 淋巴细胞。其中,NKT 细胞表达NK 细胞和T 淋巴细胞表面标志物,MAIT 细胞具有恒定TCR α 链,它们分别识别由主要组织相容性复合体Ⅰ类相关分子CD1d呈递的脂质抗原 和MHC-Ⅰ类 相 关 分 子1(MHC class-I-related molecule 1,MR1)呈递的细菌来源的小分子抗原,均可参与天然免疫和适应性免疫反应[3](图1)。本文对T 淋巴细胞亚群在PSC 中的作用机制作一阐述,以期有助于PSC的诊断及治疗。
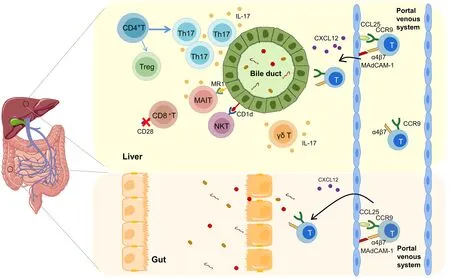
图1 原发性硬化性胆管炎中的T淋巴细胞及肝脏异常T淋巴细胞归巢Figure 1 T lymphocytes in primary sclerosing cholangitis and aberrant homing in the liver
1 CD4+ T淋巴细胞
CD4+T 淋巴细胞通过分泌细胞因子和诱导免疫细胞聚集参与自身免疫性肝病,是PSC 发病的主要免疫细胞之一。在PSC中,肝内CD4+T淋巴细胞数量增加,聚集在胆管附近,在病原体及细胞因子的刺激下,分化为不同亚型的T 淋巴细胞,如辅助性T 淋巴细胞17(Th17)和调节性T 淋巴细胞(Treg)等,介导促炎反应与抗炎反应的平衡[2]。
早期观点认为,记忆T 淋巴细胞主要驻留在不同组织中。与记忆T 淋巴细胞相比,幼稚T 淋巴细胞主要在血液和外周淋巴器官之间循环,很少驻留在肝脏等非淋巴组织中。然而近年来研究发现,多种肝病患者肝脏内均存在一群肝内幼稚样CD4+T淋巴细胞,如丙型肝炎、酒精性肝炎和非酒精性脂肪性肝炎等,但在PSC 中尤为明显。并且这些幼稚样CD4+T 淋巴细胞倾向分化为Th17,进而分泌白细胞介素(IL)-17 和IL-22等,促进炎症发生[2]。
1.1 Th17 Th17是CD4+T淋巴细胞在病原体及转化生长因子-β(TGF-β)和IL-6 等细胞因子共同诱导分化后的表型,视黄酸相关孤儿核受体γt(retinoic acidrelated orphan nuclear receptor γt,RORγt)是控制Th17分化的关键。Th17 主要通过产生特异性细胞因子IL-17介导促炎反应,参与PSC的进程。IL-17是肝脏炎症和纤维化的关键驱动因子,并与自身免疫性肝病的发展有关。IL-17可以激活肝肌成纤维细胞和肝星状细胞,增加炎症介质(如IL-6、IL-1β、TNF-α等)和促纤维化介质(如骨膜蛋白、TGF-β等)的产生,进而导致胆管炎症及纤维化[4]。
近年来研究[5]发现,与健康对照组和原发性胆汁性胆管炎相比,PSC 患者外周血和肝脏中Th17数量明显增加,伴随着IL-17 分泌增加。在微生物的刺激下,PSC 患者的单核细胞产生更多的IL-1β 和IL-6,驱动Th17 分化。IL-1β 有助于激活胆管上皮细胞,当胆管上皮细胞被激活后,除了释放大量促炎因子IL-6 和IL-23,还分泌高水平的单核细胞及Th17趋化因子C-C基序配体(C-C motif ligand,CCL)2和CCL20,进一步促进单核细胞和Th17 的分化及招募[5-6]。因此,单核细胞、胆管上皮细胞和Th17 的相互作用形成了一个正反馈循环,使大量Th17聚集在胆管周围,分泌IL-17等发挥促炎作用。此外,用RORγt反向激动剂来选择性地抑制Th17 分化后,虽然不能防止胆管炎症,但改善了胆管炎模型小鼠的肝胆损伤和纤维化[7],这也强调了Th17 在肝脏炎症中的致病作用。然而,用IL-17A抗体不能阻断胆管炎模型小鼠的疾病进展[7],可能是由于Th17 中残留其他起作用的细胞因子,如IL-17F和粒细胞-巨噬细胞集落刺激因子等,这提示了Th17参与PSC发病机制的复杂性。
1.2 Treg Treg与致炎性Th17作用相反,它在抑制免疫及调节免疫稳态中发挥了关键作用。Treg表达高亲和力的IL-2受体(IL-2 receptor,IL-2R),可以竞争性地与IL-2结合,进而促进Treg的生成及功能维持。Treg主要通过抑制致病性T淋巴细胞(如Th17细胞和CD8+T淋巴细胞)的表达和分泌抑制性细胞因子IL-10 和IL-35等来发挥免疫抑制作用,其数量和功能的失调均会打破免疫耐受,导致PSC进展[8]。
与健康对照组或其他自身免疫性肝病相比,PSC患者外周血及肝内Treg数量显著降低[8]。用IL-2R抗体消耗胆管炎模型小鼠肝内Treg 后,会增加肝内CD8+T 淋巴细胞数量,进而使TNF-α 和骨桥蛋白等细胞因子增多,加重胆道损伤及纤维化程度。相反,用低剂量IL-2 诱导肝内Treg 表达,则增加了CD39+Treg细胞的比例,导致CD8+T 淋巴细胞数量减少。CD39在炎症条件下可以稳定Treg 的抑制表型,增强其抑制活性[9]。这表明Treg 数量与PSC 疾病进展有关,Treg数量下降引起的CD8+T 淋巴细胞的增多可能是导致PSC发病的原因之一。全基因组关联研究在PSC患者中发现了IL-2 和IL-2RA 的风险等位基因[9]。因此Treg 数量的降低可能与IL-2RA 基因位点的多态性有关,但还需要进一步的研究证实。
PSC 患者肝内Treg 数量显著降低的同时也存在着功能障碍。通过对胆管炎和结肠炎小鼠模型注射IL-2/IL-2Ab 免疫复合物治疗,实现了Treg 在体内的扩增,结果发现小鼠的结肠炎得到改善,但胆管炎的严重程度并没有降低。这提示,肝内Treg 的抑制功能选择性地降低。进一步探究其机制后发现,肝脏中Treg的FoxP3 表达降低,且IL-12Rβ2 表达上调[10]。FoxP3是Treg的核心转录因子,能够上调IL-2R的表达,增强Treg 对IL-2 的敏感性,进而增加Treg 的数量并维持正常功能[8,11]。IL-12 是FoxP3 的调节因子,可通过结合表达上调的IL-12Rβ2,降低FoxP3 的表达,减少IL-2对Treg 的激活,最终导致肝内Treg 抑制能力降低[10]。此外,一项临床研究[12]也发现,通过自体体外培养Treg并单次回输来治疗1例溃疡性结肠炎伴PSC 的患者,结肠炎得到了长期有效缓解,PSC 短暂缓解后肝功能回复。这表明单次Treg 回输治疗对PSC 短暂有效,Treg 功能的丧失可能与转移的Treg 在肝脏中生存能力降低有关,若想达到PSC 长期缓解,可能需多次的Treg回输。
综上,PSC 中Th17 数量升高,伴随着Treg 减少,Th17/Treg 的平衡在PSC 的发病中具有重要意义。FoxP3 位点的甲基化状态决定了Treg 群体的稳定性,并微调了Th17/Treg平衡。然而,仅有研究[13]发现,在原发性胆汁性胆管炎患者中,FoxP3 启动子表现出高度甲基化状态,并导致Th17/Treg分化轴向Th17倾斜。对于FoxP3 启动子在PSC 中有无异常,还需要进一步的研究证实。
2 CD8+ T淋巴细胞
CD8+T 淋巴细胞是PSC 胆管中主要的组织驻留淋巴细胞,通过其介导的细胞毒作用,招募炎症细胞并分泌促炎因子来诱导炎症发生[14-15]。通过对肝脏免疫细胞进行定位后发现,PSC 患者的肝脏中存在着大量CD8+T淋巴细胞,并且主要集中在胆管上皮细胞周围,是胆道中主要的免疫细胞群。胆管中驻留的CD8+T 淋巴细胞和胆管上皮细胞共同分泌中性粒细胞趋化因子CXCL8,招募中性粒细胞聚集在胆管周围,介导炎症反应[14]。此外,CD8+T 淋巴细胞还可以分泌IFN-γ 和TNF-α 来加重胆道损伤[15]。在PSC 患者及胆管炎小鼠模型中,检测到的大量CD8+T淋巴细胞主要为缺乏CD28表型。CD28是T淋巴细胞的共刺激因子,与CD28+T 淋巴细胞相比,CD8+CD28-T 淋巴细胞内含有颗粒酶B 和穿孔素,并且在受到刺激后,IFN-γ 和TNF-α 分泌增加,因而CD8+CD28-T 淋巴细胞毒作用更强。并且CD8+CD28-T 淋巴细胞似乎是一种慢性激活的免疫细胞,不易受到Treg 的调节。因此,组织驻留的CD8+CD28-T 淋巴细胞可能是PSC 患者持续性慢性炎症的重要驱动因素[16-17]。
3 γδ T淋巴细胞
γδ T 淋巴细胞作为非常规免疫细胞,具有先天免疫细胞和适应性免疫细胞的特征。一方面,它可以通过TCR 识别病原体和宿主细胞上的保守结构,包括非肽代谢物和热休克蛋白;另一方面,它们是双阴性的,即不表达CD4 和CD8 共受体,因此,它不需要主要组织相容性复合物分子的抗原呈递,能比αβ T淋巴细胞更快地产生免疫反应。此外,γδ T 淋巴细胞在肝脏中富集,占血液中T 淋巴细胞总数的2%~10%,而占肝脏T 淋巴细胞总数的15%~25%。在PSC 患者的外周血和肝脏中,γδ T 淋巴细胞的数量较正常相比升高,且与胆管炎症的程度有关。研究[18]发现,从PSC患者肝脏中提取出来的γδ T淋巴细胞在受到毒素刺激后,倾向分化为能产生IL-17 的γδ T 淋巴细胞亚群,进而生成大量IL-17。并且相较于Th17,γδ T 淋巴细胞可能是肝脏中IL-17的主要提供者。
4 NKT
NKT 在肝脏中含量丰富。研究[19]发现,肝细胞、肝星状细胞和胆管上皮细胞等均可以作为抗原呈递细胞向NKT 呈递脂类抗原。且在PSC 患者胆汁中,含有可以激活CD1d 的脂类抗原,当脂质抗原被胆管细胞吸收并由CD1d 呈递后,可能激活周围组织中的NKT。它们可以通过快速产生促炎或抗炎细胞因子,招募并激活其他免疫细胞来指导免疫反应。此外,与健康对照组相比,PSC 的胆管上皮细胞中CD1d 表达下调,这提示CD1d 依赖的抗原呈递受到影响,表明NKT 可能在PSC 发病机制中发挥作用,但NKT 具体如何参与PSC的发病仍有待研究[20]。
5 MAIT
MAIT 在PSC 发病中的具体作用目前尚不清楚。研究[21]表明,PSC 患者的胆汁中存在着能够激活MAIT 的抗原,胆管细胞将胆道抗原呈递给MAIT,进而激活MAIT,这可能是PSC 发病机制中连接免疫系统和微生物组的病理生理途径之一。MAIT 在PSC 患者中存在着数量和功能的变化。与健康对照组相比,PSC 的外周血和肝组织中的MAIT 明显减少,但在胆道中富集,胆道中的MAIT数量是对应的外周血中的4倍[22]。并且这些MAIT 表现出激活慢和功能衰竭的特征,IFN-γ 和TNF-α 等产生受损,抗菌能力下降,这可能是由于长期暴露于促炎性细胞因子IL-12和IL-18引起的。此外,尽管MAIT 功能下降,但MAIT 在IL-12的重复刺激下,能够上调RORγt 的表达,产生大量的IL-17A,进而激活肝星状细胞,介导炎症及纤维化的进展[23]。
6 T淋巴细胞归巢
淋巴细胞归巢是指外周循环中的淋巴细胞被选择性地招募到次级淋巴器官和外周组织,介导免疫反应的过程。在炎症性肠病中,肠道相关淋巴组织中的树突状细胞识别病原体后,激活幼稚T 细胞表达“归巢受体”,即趋化因子受体CCR9 和整合素α4β7,它们分别与血管内皮细胞表面的“地址素”、趋化因子配体CCL25 和内皮黏附分子MAdCAM-1 识别并结合,进而引导T 淋巴细胞进入固有层,完成对T 淋巴细胞的招募[24]。在生理条件下,CCL25 和MAdCAM-1 的表达主要局限于肠道的血管内皮细胞。因75%的PSC患者常患有炎症性肠病(inflammatory bowel disease,IBD)[25],近年来研究[24]发现PSC 与IBD 中存在一个共同的肠-肝免疫轴。在PSC 患者的肝窦内皮细胞上,也可表达肠道中的CCL25 及MAdCAM-1,招募肠道来源的T 淋巴细胞,随后在CXCL12 的驱化作用下,将其募集到胆管周围,介导PSC的进展(图1)。
然而,最近的研究对这一发现提出了挑战。一项多中心研究[26]通过应用维多利珠单抗(α4β7整合素抗体)阻断α4β7介导的CD4+T淋巴细胞向肠道和肝脏的募集,来治疗IBD-PSC 的成年患者,结果发现,维多利珠单抗治疗可改善炎症性肠病的症状,但碱性磷酸酶等肝脏生化指标和梅奥风险评分等均未出现显著变化。这表明T淋巴细胞向肝脏的运输并不主要依赖于α4β7。另有研究[25]报道,在75%的慢性肝病患者肝脏中,均发现MAdCAM-1免疫反应阳性。与正常肝脏相比,所有慢性肝病患者的CCL25 和MAdCAM-1 的mRNA 表达均增加,并诱导了表达α4β7 和CCR9 的CD4+T 淋巴细胞的组织浸润。这些研究[25,27]表明T淋巴细胞归巢可能并不是PSC 特有的发病机制,而是慢性肝病的一种特征。
7 问题与展望
尽管目前对PSC 的发病机制有了一定的了解,且越来越多研究证实T淋巴细胞参与了PSC的发生与发展,可能有助于PSC 的治疗,但T 淋巴细胞数量繁多、种类复杂,对于其发挥的促炎与抗炎作用仍存在疑问:首先,Th17/Treg 在PSC 发病过程中涉及信号通路及分子层面上的具体机制尚未明确,异常淋巴细胞归巢是否是PSC 发病的关键机制还存在争议;其次,针对T 淋巴细胞靶向治疗的药物在胆管炎动物模型上取得了一定的进展,但能否应用于临床还需要多中心、大样本的研究。因此,未来仍需更进一步的研究来明确PSC 发病的关键机制,阐明T 淋巴细胞在PSC中的生物学功能及潜在意义,并为寻找合适的治疗靶点提供新的思路。
利益冲突声明:本文不存在任何利益冲突。
作者贡献声明:李子怡、毛小荣、李俊峰对文章的思路和设计有关键贡献,并参与修改文章关键内容;李子怡、张万洁、王富春均参与了文献检索及起草文章初稿。
