贵州野核桃天然林主要树种生态位和种间关系研究
2023-12-14吴华丽余德会杨焱冰杨小红黎大顺
吴华丽 余德会 杨焱冰 杨小红 黎大顺



摘 要:选取分布集中、种群较大且原生性强的贵州野核桃天然林群落,采用生态位分析、方差比率法、χ2检验、AC指数、Jaccard指数、Ochiai指数、Dice指数分析的方法,探明贵州野核桃群落20个主要树种生态位和种间联结特征及其相互关系。结果表明:(1)生态位宽度B(SW)i值比B(L)j值大,测度结果基本一致,群落中野核桃生态位宽度值最大,白檀生态位宽度值最小;重要值和生态位宽度值依次排前三的是野核桃、灯台树、漆树,他们构成了所处群落优势种或建群种;(2)贵州野核桃天然林主要树种利用资源的相似性程度和生态位重叠大多数种对都较小,种间竞争相对较弱,但青榨槭与主要树种资源利用相似性和重叠值都较大,其种间竞争最激烈;(3)生态位相似性比例值和生态位重叠值的对数分布不均,且小值的对数中数值最小(Cih=0,αih=0),大值的对数中数值较大(Cih=1,αih=0.89);(4)贵州野核桃天然林群落中主要树种总体关联性不显著,群落处于演替中期。贵州野核桃天然林群落中优势种群对环境资源的利用较强,种群伴生种少部分竞争相当激烈,大部分竞争相对较弱,野核桃幼树的更新状况较差,群落结构可能具有一定的不稳定性,本研究可为贵州野核桃的科学保护与合理开发利用提供理论依据。
关键词:野核桃;主要树种;生态位特征;种间联结;贵州
中图分类号:Q948
文献标志码:A
生态位研究和种间联结性的研究是种群生态学和群落生态学的重要内容[1-2],与此相关产生了诸多关于生态位的研究理论,GRINNELL、ELTON和HUTCHINSON等分别提出的空间生态位、营养生态位和多维超体积生态位理论是目前被广泛接受的理论[3-5]。后有研究人员评析各生态位概念的内涵要点,并提出可将生态位视为一个宏观概念,在具体应用中才需采用细分的生态位定义和内涵[6]。生态位理论作为生态学的一个主要内容,已经在群落结构[7-8]、群落种间关系[9]、演替[10]及生物多样性[11]等研究中得到广泛应用,并取得了较大进展[12]。森林群落中植物树种之间的联结性是群落的重要数量和结构指标,是群落结构形成和演化的重要基础[13],静态描述了群落中各种群在一定时期内的相互关系[14],是种间相互共存、竞争或独立等状态的体现。鉴此,开展天然林群落主要树种生态位及种间联结研究,认识群落动态和种群生态适应性以及树种组成和种间关联的影响状况,对野核桃植物群落科学保护与培育利用具有重要意义。
野核桃Juglans cathayensis为胡桃科Julandaceae胡桃属Juglans落叶乔木,是我国特有的多功能经济生态树种[15-16]。野核桃材质坚实,种子油可作润滑油,可制肥皂,亦可食用;树皮和果皮含鞣质且皮厚;果实内含有亚麻酸、亚油酸及多种微量元素,极具药用价值和经济价值[17-19]。该树种在贵州以绥阳、石阡、雷山、桐梓、盘县、普安、道真和印江等地为主要分布区域[11]。目前,对野核桃的研究主要集中在病虫害防治、群落特征、种群结构、遗传多样性、保护生物学、濒危机制、化学成份等[20-22]。其中,有学者对山西太岳山野核桃林开展了优势种群生态位研究,预测野核桃能成为群落建群种,但生态位重叠值突出,如果环境资源不足将会发生种间竞争,影响野核桃群落结构及其种群发展[17]。贵州自然地理位置和环境独特,是野核桃主要分布区之一,其种群在天然林群落中的地位、资源的利用状况以及主要树种间相互关系的研究尚未见报道。本文选取贵州境内宽阔水、佛顶山和雷公山3个国家级自然保护区分布密集、集中且保存原生性强的野核桃天然林,对其生态位和种间联结特征开展耦合研究,可有效揭示贵州分布的野核桃天然林主要树种的生态适应性以及共存机制,旨在为野核桃保护与开发利用提供科学数据。
1 材料与方法
1.1 研究区概况
选取贵州宽阔水、佛顶山和雷公山3个国家级自然保护区为主要研究区,地处东经107°02′—108°22′,北纬26°15′—28°19′。研究区土壤以黄壤和黄棕壤为主,海拔650~2 178.8 m,年平均气温11.7~15.2 ℃,年降水量1 100~1 450 mm,年平均相对湿度超过80 %。研究区属亚热带湿润季风气候,地形地貌复杂多变,冬无严寒,夏无酷暑,云雾多,日照少,主要植被类型为山地常绿落叶阔叶混交林和山地落叶阔叶林、山顶杜鹃矮林和苔藓矮林,低海拔区域主要为地带性常绿阔叶林,植树种类丰富。
1.2 样地设置与调查
在宽阔水、佛顶山和雷公山3个国家级自然保护区分别设置20 m×30 m的6个典型样地,共18个,样地合计面积10 800 m2。采用相邻格子法(基本单元格设为10 m×10 m)将样地划分为6个小样方。对样地内所有乔木(胸径≥5.0 cm)树种进行每木调查,记为树种名、胸径(cm)、树高(m)、冠幅(m)等因子,生境调查样地岩石裸露率、土壤类型、坡度、坡向、海拔、总盖度等特征(表1)。
1.3 数据处理及计算方法
1.3.1 主要树种的确定
通過计算研究区野核桃天然林18个群落样地树种的重要值,筛选至少在2个以上样地出现的树种,或仅在单个样地里面重要值大于30%的植树种类记为群落的主要树种[11]。
重要值计算公式:重要值=(相对密度+相对频度+相对显著度)/3
1.3.2 生态位宽度
(1)Shannon-Weinner(1948)[23]多样性指标的生态位宽度
B(sw)i=-1logS∑rj=1PijlogPij
式中,Pij是树种i利用第j资源占它利用全部资源位的比例;S为种群数;r为资源位数。
上式中,Pij=NijYij,Yij=∑rj=1Nij,其中,Nij为树种i在第j资源位的重要值,Yij为树种i利用全部资源位的重要值之和。生态位宽度具有域值[0,1],即树种利用一个资源位,其B(sw)为0,而利用了全部资源位,其值为1。
(2)Levins(1968)[24]的生態位宽度指数
B(l)i = 1∑rj = 1P2ij
式中,r、Pij意义同上;生态位宽度B(l)具域值[1/r,1]。
1.3.3 生态位相似性比例
Cih=1-12∑rj=1Pij-Phj
式中,Cih表示树种i与树种h的相似程度,且有Cih=Chi,具有域值[0,1];Pij和Chi分别为树种i和树种h在资源位j上的重要值百分率。
1.3.4 生态位重叠
Pianka指数[25]
αhi=∑rj=1PijPhj∑rj=1(Pij)2∑rj=1(Phj)2
式中,αih=αhi,αih为树种i重叠树种h的生态位重叠值,αhi为树种h重叠树种i的生态位重叠值,αih和αhi具有值域[0,1]。
1.3.5 联结性分析
采用方差比率(Vr)法判断植物群落的总体关联性,以统计量W检验联结性的显著度[26],若Vr>1,表示种间呈净的正关联;若Vr<1,表示种间呈净的负关联;若Vr=1,表示所有种间无关联,采用统计量(W)检验关联程度,若W<χ20.95(N)或W>χ20.05(N),种间总体联结显著;若χ20.95(N)<W<χ20.05(N),则种间总体联结不显著,其中,N为样方数。
再采用Yates校正的χ2检验定性判定种间联结性[27]。若χ2<3.841,表示种间联结性不显著(P>0.05);若3.841<χ2<6.635,表示种间联结性显著(0.01<P<0.05);若χ2>6.635,则表示种间联结性极显著(P<0.01)。χ2值不能定量区分连结强度,因此结合联结系数(association coefficient,Ac)、Jaccard指数(Ja)、Ochiai指数(Oi)、Dice指数(Di)进一步分析[28]。
2 结果与分析
2.1 主要树种重要值
重要值是树种综合数量的重要性指标,表示树种在群落中的地位和作用,反映树种在群落中的优势程度[29]。本研究中,重要值结果(表2)表明,主要树种有20种。野核桃的重要值之和最大(17.45),占主要树种重要值总值的45.53 %,其次是灯台树Cornus controversa(3.59)和漆树Toxicodendron vernicifluum(2.05),分别占主要树种重要值总值的9.38 %和5.35 %。野核桃在18个资源位中的重要值排序在第一位达15个资源位,有2个资源位排在第二,分别是样地FDS-5和FDS-6,有1个资源位排在第三,是LGS-6。可见野核桃种群在宽阔水、佛顶山和雷公山3个国家级自然保护区分布的天然林中处于优势地位,且在资源位中为建群种。
2.2 主要树种生态位宽度
生态位宽度是度量植树种群对环境资源利用状况的尺度,种群生态位宽度越大,则它对环境的适应能力越强[25]。由表3可知,对比各资源位中任一树种B(SW)i的值都要比B(L)j的值大,且两种生态位宽度公式的测度结果基本一致。生态位宽度值降序依次为野核桃、灯台树、漆树、石木姜子Litsea elongata var. faberi等。上述4个树种均占较大生态位宽度,其值合计分别占B(SW)i和B(L)j总值和的31.84 %和41.00 %,其中,野核桃树种的生态位宽度最大(0.947、0.905),而白檀Symplocos tanakana生态位宽度值最小,B(SW)i和B(L)j分别为0.092和0.065,生态位宽度较窄,仅在2个资源位中出现。
2.3 主要树种生态位相似性比例
生态位相似性比例是指两个种群之间利用资源的相似程度。由表4可知,生态位相似性比例值为零的树种对有71对,占总对数的37.37 %;0<Cih≤0.4的有87对,占总对数的45.79 %;0.4<Cih≤0.8的有21对,占总对数的11.05 %;0.8<Cih≤1.0的有11对,占总对数的5.79 %。其中,Cih值域(0.8,1.0]有11对,且都是青榨槭Acer davidii与群落中主要树种形成的对数,甚至有Cih=1.0达4对,分别是青榨槭与杉木Cunninghamia lanceolata、四照花Cornus kousa subsp.chinensis、白檀和多脉青冈Quercus multinervis,而野核桃与群落中其他主要树种生态位相似性比例值都在0.5以下。
2.4 主要树种生态位重叠
较高的生态位重叠意味着种群之间对环境资源利用的能力越相似,因而可能存在着激烈的竞争[30]。如表4所示,αih介于0~0.887之间,αih平均值为0.179,αih≥0.500的有22个种对,占总种对数的11.58 %,最大的是山樱桃Prunus serrulata与白辛树Pterostyrax psilophyllus,为0.887;处于(0.200,0.500)的有42个种对,占总种对数的22.11 %;介于(0,0.200)的有126个种对,占总种对数的66.32 %。灯台树与香果树Emmenopterys henryi等80个种对不发生生态位重叠(αih=0),占总种对数的42.11 %。山樱桃与白辛树、灯台树与青榨槭具有较大的生态位重叠值(0.887、0.852);野核桃与其它树种间生态位重叠平均值为0.388,且各对生态位重叠值都大于总的平均值(0.179),大于0.5的有4对。
2.5 主要树种总体联结性
通过计算(表5),Vr=0.720<1,表明20个主要树种总体上表现为负联结,利用统计量W检测Vr取值偏离1的显著程度,W=12.880,χ20.95(18)=9.395,χ20.05(18)=28.869,9.395<W<28.869,可知野核桃天然林群落中主要树种总体关联性不显著。
2.6 种间联结性分析
贵州野核桃天然林群落主要树种种对的χ2统计量检验结果如表6所示。在190个种对中,有66个种对(占总种对34.74 %)呈正相关,102个种对(占总种对53.68 %)呈负相关,22个种对(占总种对11.58 %)表现为不相关。其中,仅云贵鹅耳枥-青榨槭、云贵鹅耳枥-大花野茉莉2个种对表现为极显著正相关(χ2>6.635),漆树-四照花、漆树-多脉青冈、青榨槭-大花野茉莉、杉木-四照花、四照花-多脉青冈、山樱桃-白檀6个种对呈显著正相关(3.841 >χ2 >6.635)。野核桃与其他主要树种之间均表现为不相关。190个种对中有95.79 %的种对相关性表现为不显著。
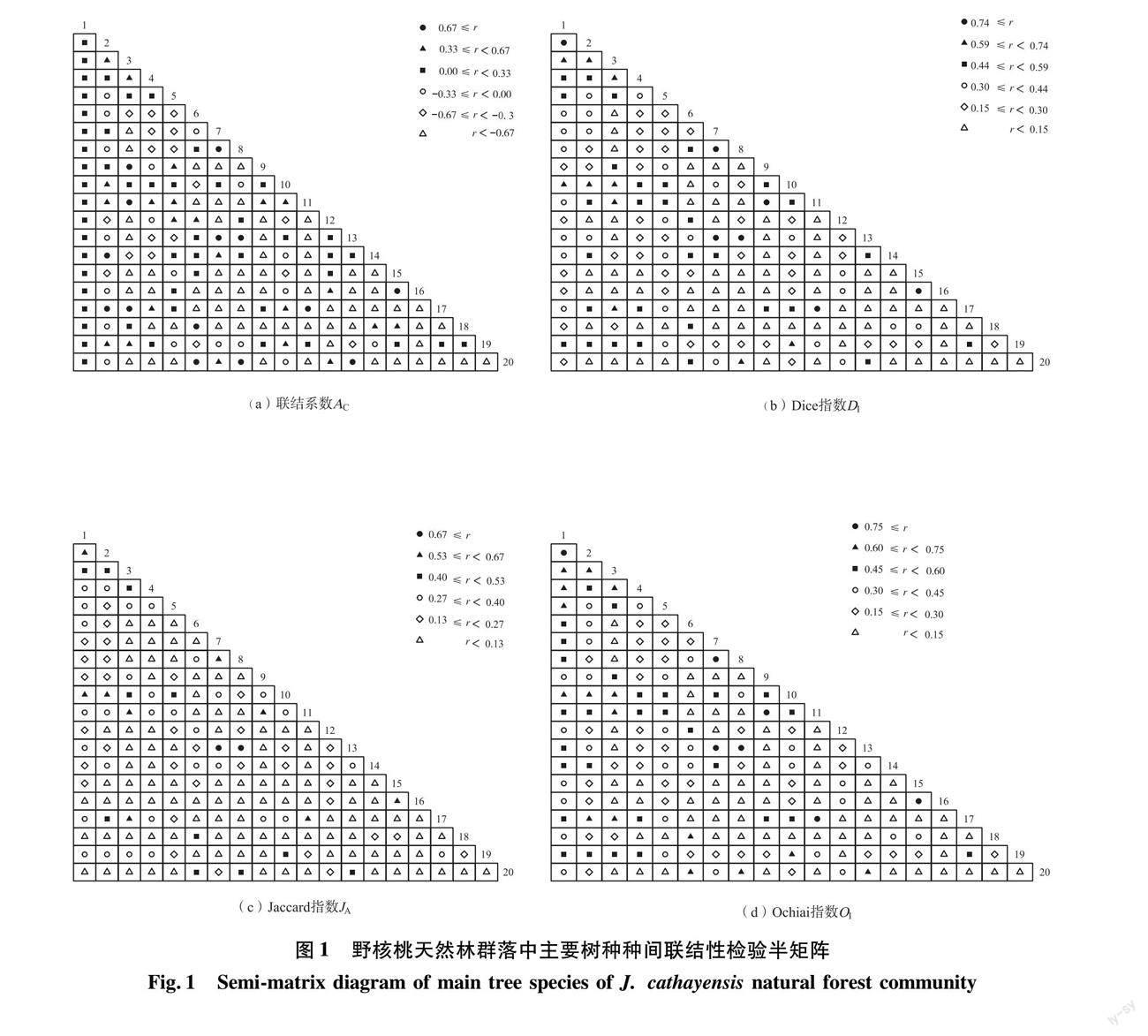
从图1(a)可以看出,Ac(r) ≥0.67的种对有青榨槭-大花野茉莉等13个种对;0.33≤ Ac(r)< 0.67的种对有漆树-灯台树等22个种对;0.00 ≤Ac(r) <0.33的种对有贵州野核桃-川桂等51个种对;此外,漆树-青榨槭等65个种对的Ac(r)<-0.67。从图1来看,云贵鹅耳枥-青榨槭、云贵鹅耳枥-大花野茉莉2个种对表现出极强的联结性(Di(r)、Ja(r)、Oi(r)值均大于等于0.67);四照花-多脉青冈、野核桃-灯台树、山樱桃-白檀、杉木-四照花4个种对也表现出非常强的联结性(Di(r) ≥0.74,Oi(r) ≥0.75,0.53≤Ja(r) <0.67);野核桃-石木姜子、灯台树-石木姜子、漆树-四照花、漆树-多脉青冈4个种对的联结性均较紧密(0.59≤ Di(r) <0.74,0.60 ≤Oi(r) <0.75,0.53 ≤Ja(r) <0.67);野核桃-漆树、灯台树-漆树、半月边-漆树、石木姜子-漆树、石木姜子-川桂、大花野茉莉-栲这6个种对在Di、Oi上表现较紧密(0.59 ≤Di(r) <0.74,0.60 ≤Oi(r) <0.75);有68个种对表现出极低的联结性(Di(r)、Oi(r) <0.15,Ja(r) <0.13)。
3 结论与讨论
在贵州分布的野核桃天然林群落20个主要树种中,野核桃的生态位宽度值最大,主要是因为野核桃是落叶乔木阳性树种,且种群整体呈集群分布[16],适应能力强,在多个群落中皆能处于优势地位,因此,往往形成建群种。灯台树、漆树、石木姜子、半边月Weigela japonica var. sinica、枫香树Liquidambar formosana、四照花、云贵鹅耳枥和多脉青冈等生态位宽度值大于0.5,表明它们数量多,分布范围广,适合该区的环境生长,在群落中占优势地位,说明这些树种具有较大的发展潜力,主导群落结构和生态系统稳定,对贵州野核桃群落发展起决定作用[31]。野核桃、灯台树和漆树重要值和生态位宽度都是前三位,构成了贵州野核桃天然林的建群种或优势种。研究区属于山地温凉气候,主要植被类型为山地落叶阔叶林或以落叶树种为主的常绿落叶阔叶混交林,野核桃生长较为速生,在该气候条件和山地环境下竞争能力强。而白檀、香果树和栲Castanopsis fargesii生态位宽度值小,在资源竞争中处于劣势,对当地环境的生态适应能力较弱,是野核桃等主要优势种的伴生种,在群落中不起主要作用。
本研究发现,野核桃天然林群落中主要树种出现生态位相似性比例值和生态位重叠值的对数都偏小,且小值的对数中数值最小(Cih=0,αih=0),大值的对数中数值非常大(Cih=1,αih=0.887)。表明野核桃天然林群落中主要树种间多数存在对不同的资源利用较多,且能在多种环境条件下共存的特点,在对相同的资源利用时竞争较激烈。因此,野核桃在研究区内具有较强的适应能力和竞争能力。研究区内贵州野核桃与其他主要树种的生态位相似性比例值和生态位重叠值越大,其他主要树种生态位宽度越大,反之亦然。说明野核桃对资源的利用相似程度较低(Cih<0.5),对资源需求产生的竞争较小,有利于群落稳定。
本研究中,野核桃天然林群落主要树种间的总體联结为负联结,但不显著,方差比率(Vr)<1,表明群落目前还处于不稳定阶段,受到外界干扰时数量和种类易发生波动。χ2检验结果表明:野核桃天然林群落主要树种间呈不显著联结和无联结的种对共182个,占95.79%,联结系数Ac用来进一步检验由χ2所测出来的结果及说明种间联结程度[32],综合Ac结果来看,20个主要树种之间大部分呈现无显著相关,树种间相对独立,群落中野核桃种群与其他主要树种均不存在显著联结性,这与薛卫星等[33]的研究结果相似,可能由于野核桃是阳性树种,且为高大乔木,对环境中的各种资源的利用具有优势。在Ac的数值上,数值为-1的有66对,树种间呈极显著负联结,这可能是因为两个树种没有出现在同一个样地,导致Ac值为-1,结果中极显著负联结的种对较多,可能是树种多度较低导致[34]。
Ja、Oi、Di均能较准确地反映种对间正联结程度并避免Ac值偏高或Ac值偏低的影响[35],本研究中,Oi和Di数值所反映的树种联结性结果较为一致,Ja数值有一定偏差,但是总体趋势一致。云贵鹅耳枥-青榨槭、云贵鹅耳枥-大花野茉莉的χ2、Ac、Ja、Oi、Di都表现为极强的正相关性,可能是因为这3个种都为小乔木,生态习性相似,出现在相同生境的可能性很大,从而产生紧密的相关性。四照花-多脉青冈、野核桃-灯台树、山樱桃-白檀、杉木-四照花4个种对的Ja、Oi、Di表现出非常强的联结性,可能因为野核桃、灯台树、四照花、多脉青冈生态位宽度值在主要20个树种中排在前列,在群落中广泛分布,所以彼此之间同时出现的概率较高。有68个种对的Ja、Oi、Di表现出极低的联结性,说明这些种对彼此之间的依懒性不强,种对间较为独立。野核桃与漆树、半边月、野茉莉、枫香树这些树种共同出现的频率较高,但是这些树种与野核桃之间的相关性不显著,这与前人的研究结果相一致[33],可能是因为树种间在各群落中占据了不同的生态位,以及林分中垂直分布格局不同,导致这些树种能够共存。因此,贵州野核桃天然林分布的群落中主要树种间竞争相对较弱,群落处于不稳定阶段,受到外界干扰时数量和种类易发生波动。本研究有效揭示贵州野核桃天然林主要树种的生态适应性以及共存机制,为野核桃种质资源的科学保护与合理利用提供可靠的科学依据。
参考文献:
刘润红, 陈乐, 涂洪润, 等. 桂林岩溶石山青冈群落灌木层主要树种生态位与种间联结[J]. 生态学报, 2020, 40(6): 2057-2071.
[2] 包庆德, 夏承伯. 生态位: 概念内涵的完善与外延辐射的拓展:纪念“生态位”提出100周年[J]. 自然辩证法研究, 2010, 26(11): 43-48.
[3] GRINNELL J. The niche-relationship of the california thrasher[J]. The Auk, 1917, 34(4):427-433.
[4] ELTON C S. Animal ecology[M]. Chicago: University of Chicago Press, 1927.
[5] HUTCHINSON G E. Concluding remarks. populations studies: animal ecology and demography[J]. Cold Spring Harbor Symposium of Quantitative Biology, 1957, 22: 415-427.
[6] 彭文俊, 王晓鸣. 生态位概念和内涵的发展及其在生态学中的定位[J]. 应用生态学报, 2016, 27(1): 327-334.
[7] 王洁. 吉林蛟河红松阔叶林群落结构特征及其生态位研究[D]. 吉林: 北华大学, 2022.
[8] 龙翠玲.茂兰喀斯特森林林隙更新生态位的研究[J]. 山地农业生物学报, 2006, 25(4): 302-306.
[9] 徐高兴, 赵鹏, 陈思航, 等.民勤绿洲荒漠过渡带沙拐枣群落种间关联及生态位研究[J]. 西北林学院学报, 2023, 38(1): 25-33.
[10]习靓靓. 河南白云山温带落叶阔叶林土壤微生物多样性及其分布机制[D]. 郑州: 河南农业大学, 2022.
[11]党柯柯, 侯金凤, 刘洪, 等. 生姜根系不同生态位细菌群落多样性特征、组成及结构差异[J]. 生态学报, 2023, 43(4): 1691-1702.
[12]余德会, 袁丛军, 安明态, 等. 赤水河流域清香木天然群落主要树种生态位研究[J]. 西部林业科学, 2014, 43(6): 91-96.
[13]LAUREN F H, THOMAS D L. Temporal patterns of vascular plant diversity in southeastern New Hampshire forests[J]. Forest Ecology and Management, 2003, 185(1): 5-20.
[14]SU S J, LIU J F, HE Z S, et al. Ecological species groups and interspecific association of dominant tree species in Daiyun mountain National Nature Reserve[J]. Journal of Mountain Science, 2015, 12(3): 637-646.
[15]娄丽, 胡彬, 张亚洲, 等. 贵州野核桃砧木育苗关键技术及苗木质量评价[J]. 种子, 2018, 37(8): 138-140.
[16]张维, 焦子伟, 尚天翠, 等. 新疆西天山峡谷海拔梯度上野核桃种群统计与谱分析[J]. 应用生态学报, 2015, 26(4): 1091-1098.
[17]程永生. 山西太岳山野核桃林優势种群生态位研究[J]. 山西师范大学学报(自然科学版), 2017, 31(4): 71-75.
[18]吴明, 李锋, 王茹琳, 等. 四川省野核桃生境适应性及适生区划研究[J]. 四川林业科技, 2022, 43(1): 82-86.
[19]陈超, 胡钰, 孙家祥, 等. 野核桃叶化学成分研究[J]. 中草药, 2011, 42(11): 2177-2180.
[20]王博, 巴雪瑞, 黎一阳, 等. 坡向与龄级对新疆野核桃自然保护区野核桃病害的影响[J].应用生态学报, 2023, 34(1): 39-46.
[21]王东升. 山东省紫椴和野核桃AFLP遗传多样性研究[D]. 泰安: 山东农业大学, 2013.
[22]张维. 新疆天山峡谷渐危植物野核桃保护生物学基础研究[D]. 长春: 东北师范大学, 2016.
[23]SHANNON, C E. A mathematical theory of communication[J]. The Bell System Technical Journal, 1948, 27: 379-656.
[24]LEVINS, R. Evolution in changing environments: some theoretical explorations[M]. Princeton: Princeton University Press, 1968.
[25]袁丛军, 喻理飞, 安明态, 等. 岩生红豆天然林主要树种生态位研究[J].生态科学, 2013, 32(4): 487-493, 528.
[26]DOLPH S. A variance test for detecting species associations, with some example applications[J]. Ecology, 1984, 65(3): 998-1005.
[27]張金屯. 数量生态学[M]. 1版. 北京: 科学出版社, 2004.
[28]江焕, 张辉, 龙文兴, 等. 金钟藤入侵群落的种间联结及生态位特征[J]. 生物多样性, 2019, 27(4): 388-399.
[29]王立龙, 王广林, 黄永杰, 等. 黄山濒危植物小花木兰生态位与年龄结构研究[J]. 生态学报, 2006, 26(6): 1862-1871.
[30]张芝萍, 李得禄. 河西走廊黑果枸杞群落植物生态位特征研究[J]. 安徽农业科学, 2022, 50(17): 62-65, 97.
[31]杨持. 生态学[M]. 2版. 北京: 高等教育出版社, 2008.
[32]郭志华, 卓正大, 陈洁, 等. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J].植物生态学报, 1997, 21(5): 33-41.
[33]薛卫星, 李春辉, 艾训儒, 等. 鹅掌楸天然林优势树种生态位与种间联结性[J]. 森林与环境学报, 2023, 43(1): 26-34.
[34]许金石, 陈煜, 王国勋, 等. 陕北桥山林区主要木本植物群落种间联结性[J]. 西北植物学报, 2014, 34(7): 1467-1475.
[35]刘翔宇, 何东, 田文斌, 等. 浙江普陀山岛森林木本植物的种间关联格局[J]. 植物生态学报, 2017, 41(12): 1219-1227.
(责任编辑:于慧梅)
Ecological Niche and Interspecific Relationship of Main Tree Species in
Natural Forest of Juglans cathayensis in Guizhou
WU Huali1,2, YU Dehui*3, YANG Yanbing4, YANG Xiaohong2, LI Dashun5
(1.Guizhou Forestry Information and Publicity Center, Guiyang 550001, China; 2.Walnut Research Institute of
Guizhou Province, Guiyang 550005, China; 3.Guizhou Leigongshan National Nature Reserve Administration,
Leishang 557199, China; 4.Forestry Academy of Guizhou Province, Guiyang 550005, China; 5.College of Forestry, Guizhou University, Guiyang 550025, China)
Abstract:
The natural forest community of Juglans cathayensis in Guizhou Province with concentrated distribution, large population and strong original nature was selected, and ecological niche analysis, variance ratio method χ2 Test, AC index, Jaccard index , Ochiai index, and Dice index analysis methods were used to explore the ecological niche and interspecific association characteristics of 20 major tree species in the J. cathayensis community in Guizhou and their relationships. The results showed that: (1) The ecological niche width value of B(SW)i was larger than the value of B(L)j, and the measurement results were basically the same. In the community, the ecological niche width value of J. cathayensis were the largest, and the ecological niche width value of Symplocos paniculata was the smallest; The top three in terms of important value and ecological niche width value were J. cathayensis,Cornus contorsa,and Toxicodendron vernicifluum, which constitute the dominant species or constructive species of the community; (2) The similarity and niche overlap of the utilization resources of the main tree species were mostly small, and the interspecific competition was relatively weak. However, the similarity and overlap of the utilization resources of Acer davidii and the main tree species were large, and the interspecific competition was the most intense; (3) The logarithmic distribution of ecological niche similarity ratio value and ecological niche overlap value was uneven, the logarithmic value of small value was the smallest (Cih=0, αih=0), and the logarithm of the larger value had a larger median value (Cih=1, αih=0.89); (4) The overall correlation of the main tree species in the natural forest community of J. cathayensis in Guizhou was not significant, and the community was in the middle stage of succession. The dominant population in the natural forest community of J. cathayensis in Guizhou had a strong utilization of environmental resources. The competition among a small number of accompanying species was quite fierce, while the competition among most was relatively weak. The regeneration status of young trees of J. cathayensis was poor, and the community structure might have some instability. This study can provide a theoretical basis for the scientific protection and rational development and utilization of J. cathayensis in Guizhou.
Key words:
Juglans cathayensis; main tree species; ecological niche characteristics; interspecific association; Guizhou
