草甸草原植物中微量元素吸收特点及其对硫添加的响应
2023-12-13张玉革刘梦霖高金影李天鹏
张玉革, 刘梦霖,, 高金影,, 冯 雪, 李天鹏, 姜 勇,3
(1. 沈阳大学 区域污染环境生态修复教育部重点实验室, 辽宁 沈阳 110044;2. 中国科学院沈阳应用生态研究所 额尔古纳森林草原过渡带生态系统研究站, 辽宁 沈阳 110016;3. 河北大学 生命科学学院, 河北 保定 071002)
土壤酸化是一个全球性环境问题[1],可导致土壤肥力下降、出现重金属迁移增加等土壤退化现象。随着工业化和城市化进程的加快,化石燃料燃烧及氮肥使用增加等会排放出大量的二氧化硫和氮氧化物[2],加速了对土壤圈和生物圈的酸化,给自然环境和生物健康造成巨大威胁。由于减排策略实施和新能源的开发利用,全球范围内大气硫沉降逐渐减少,但中国、印度等国家仍维持在较高水平[3]。过去20年,我国受硫沉降影响的区域由南部和东部向北部和西部扩散,内蒙古自治区东北部草甸草原区硫沉降量增加了1倍左右,未来可能在较长时间范围内持续增加[4]。土壤酸化影响草原植物群落结构及植物养分吸收,而草原植物叶片和根系中微量元素吸收如何响应硫沉降及导致的土壤酸化问题等相关研究相对较少。
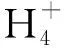
在英国开展的长达15年的野外试验[14]表明,元素S添加导致的土壤酸化对土壤和植物的影响是长期存在的,土壤酸化显著降低了土壤有效Ca浓度,进而又降低了植物中的Ca浓度,而对植物中的Fe、Mn浓度无显著影响。草地生态系统研究[15]发现,针茅和冷蒿2个优势物种叶片Mn浓度均与土壤有效Mn浓度呈显著正相关,而植物Fe吸收不仅受土壤有效Fe浓度影响,还受植物Fe、Mn元素拮抗作用调控。与植物叶片不同,根系对水分和养分的竞争强度远远高于地上部分对资源的竞争[16]。有研究[17]发现,樱桃番茄随着土壤酸化程度增加会降低根系Mg和Fe浓度,而增加根系Mn浓度。而在植物群落上,探讨草原植物根系金属养分吸收对土壤酸化响应的相关研究还较少。
本研究通过呼伦贝尔草甸草原野外单质S添加模拟土壤酸化控制实验,探究S沉降及土壤酸化背景下植物群落叶片及根系中微量元素的差异性吸收及其与土壤理化性质和有效养分之间的关系,研究结果有助于评估和预测半干旱草原土壤-植物养分循环过程对土壤酸化的响应。提出3个科学假设:
假设① 不同草原植物物种有不同的养分吸收偏好,因此不同物种叶片中量及微量元素浓度可能会存在显著差异;
假设② S添加导致的土壤酸化可加速土壤盐基离子淋失,进而抑制不同物种叶片和植物群落叶根对Ca、Mg元素吸收;
假设③ S添加导致的土壤酸化可促进土壤微量元素活化,进而增加物种叶片和植物群落叶根对Mn元素的奢侈吸收,但由于植物Fe、Mn吸收的拮抗作用,植物对Fe的吸收可能受到抑制植物Mn吸收的抑制。
1 研究区域与研究方法
1.1 研究区域概况
研究地点位于内蒙古自治区呼伦贝尔市的中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统研究站(50°12′19″ N,119°30′28″E)。该区域为温带寒温带大陆性气候,海拔523 m。具有鲜明的季节变化特点,春季降水少,温度回升快;夏季短暂燥热,雨量充沛,雨热同期;秋季降温快,初霜早;冬季寒冷漫长降水少。年平均降水量约363 mm,其中约70%的降水发生在5月至9月之间,年平均气温-2.45 ℃。植被以羊草(Leymuschinensis)、贝加尔针茅(Stipabaicalensis)、寸草苔(Carexduriuscula)、糙隐子草(Cleistogenessquarrosa)等禾本科植物为优势物种,以柴胡(Bupleurumscorzonerifolium)、白头翁(Pulsatillaturczaninovii)、达乌里芯芭(Cymbariadahurica)、冷蒿(Artemisiafrigida)等物种为常见种。该研究区的草原类型为草甸草原,土壤类型为黑钙土,土壤有机质质量分数为50~60 g·kg-1,土壤砂、粉、黏粒浓度分别占39%、37%、24%,容重约为1.21 g·cm-3。
1.2 试验设计
本试验依托额尔古纳硫添加试验平台。硫添加试验开始于2017年5月,为随机区组试验,包括8个硫添加处理,分别为0、1、2、5、10、15、20、50 g·m-2·a-1。每个处理5次重复,每个小区为6 m×6 m,相邻小区之间有2 m缓冲带,共计40个小区。选取元素硫粉末(>99.9%)来模拟硫输入及其导致的土壤酸化效应。自2017年起,每年5月中旬将不同剂量硫磺粉末与200 g处理过的土壤混合后均匀撒施于相应小区。元素硫添加试验2年后(2019年8月),土壤pH值随硫添加量增加显著下降,各处理0~10 cm土层土壤pH值分别为6.95、6.80、6.69、6.70、6.77、6.47、6.17、5.19。
1.3 样品采集与测定
1) 样品采集:在试验处理的第3年(2019年),于8月中旬采集土壤样品(0~10 cm)。利用直径5 cm土钻在每个小区随机选取5点取土并混合为1份土壤样品,共采集土壤样品40份。所取土壤样品过2 mm筛,去除植物残体和砾石,在实验室自然风干,用于测定土壤理化性质。植物样品同样采集于2019年8月中旬,在每个小区随机放置1个1 m×1 m样方框,分物种剪下框内所有植物并装入写好编号的信封内。所有植物带回实验室先在烘箱105 ℃杀青30 min,后在65 ℃下烘48 h至恒重。选取针茅、糙隐子草、白头翁、达乌里芯芭4个物种叶片进行前处理,用Retsch M400型球磨仪(德国莱驰公司生产)粉碎过100目筛(孔径为0.149 mm)以备用于测定叶片元素质量分数。之后进行植物群落根系样品采集,在每个小区用直径为5 cm的根钻采集3钻0~10 cm土块,筛分出根系,冲洗干净,在65 ℃下烘干,球磨仪粉碎过100目筛,备用。
2) 土壤及植物指标测定:土壤pH值采用S210型酸度计(德国梅特勒-托利多公司生产)测定(土水为比1∶5)。土壤交换性酸采用氯化钾(KCl)浸提、氢氧化钠(NaOH)中和滴定法测定。土壤交换性钙镁(Ca2+、Mg2+)采用1 mol·L-1的乙酸铵(NH4OAc)浸提,土壤有效态微量元素(Fe2+、Mn2+)采用二乙三胺五乙酸(DTPA)法浸提、AA6800型原子吸收分光光度计(日本岛津公司生产)测定。植物叶片及根系元素质量分数采用H2SO4-HClO4消煮后用5100ICP-OES型电感耦合等离子体发射光谱仪(美国珀金埃尔默公司生产)测定。
1.4 数据处理
植物群落根系养分质量分数为混合根系养分质量分数的测定值,群落叶片养分质量分数采用式(1)计算:
CWM= ∑piti。
(1)
式中:CWM为群落叶片养分质量分数,mg·kg-1或g·kg-1;pi为物种i的地上生物量占地上总生物量的百分比,%;ti为物种i的养分质量分数,mg·kg-1或g·kg-1。
采用SPSS统计软件进行邓肯多重比较法检验植物中微量元素质量分数在硫添加处理下的差异显著性(P<0.05),并用Peason相关分析,分析土壤指标和植物元素质量分数之间的相关关系。利用Origin 8.5软件作图,图中数据为平均值±标准误。
2 结果与分析
2.1 物种水平叶片中微量元素质量分数对硫添加响应
硫添加下草甸草原不同植物物种叶片对中微量元素Ca、 Mg、 Fe、Mn的吸收具有显著差异,如图1所示,杂类草(白头翁和达乌里芯芭)叶片Ca和Mg质量分数显著高于禾草(针茅和糙隐子草),而杂类草叶片Fe、Mn质量分数则显著低于禾草。硫添加处理下,植物物种的Ca质量分数较为稳定,但针茅和白头翁叶片Ca质量分数在硫添加量为50 g·m-2·a-1时表现为显著下降(图1(a));4个物种叶片Mg质量分数对硫添加的响应均不显著(图1(b));硫添加显著降低白头翁和达乌里芯芭叶片Fe质量分数,而加硫对针茅和糙隐子草叶片Fe质量分数有正向影响,但不同硫添加量间差异不显著(图1(c));随硫添加量的增加,4个物种叶片Mn质量分数均显著增加(图1(d))。
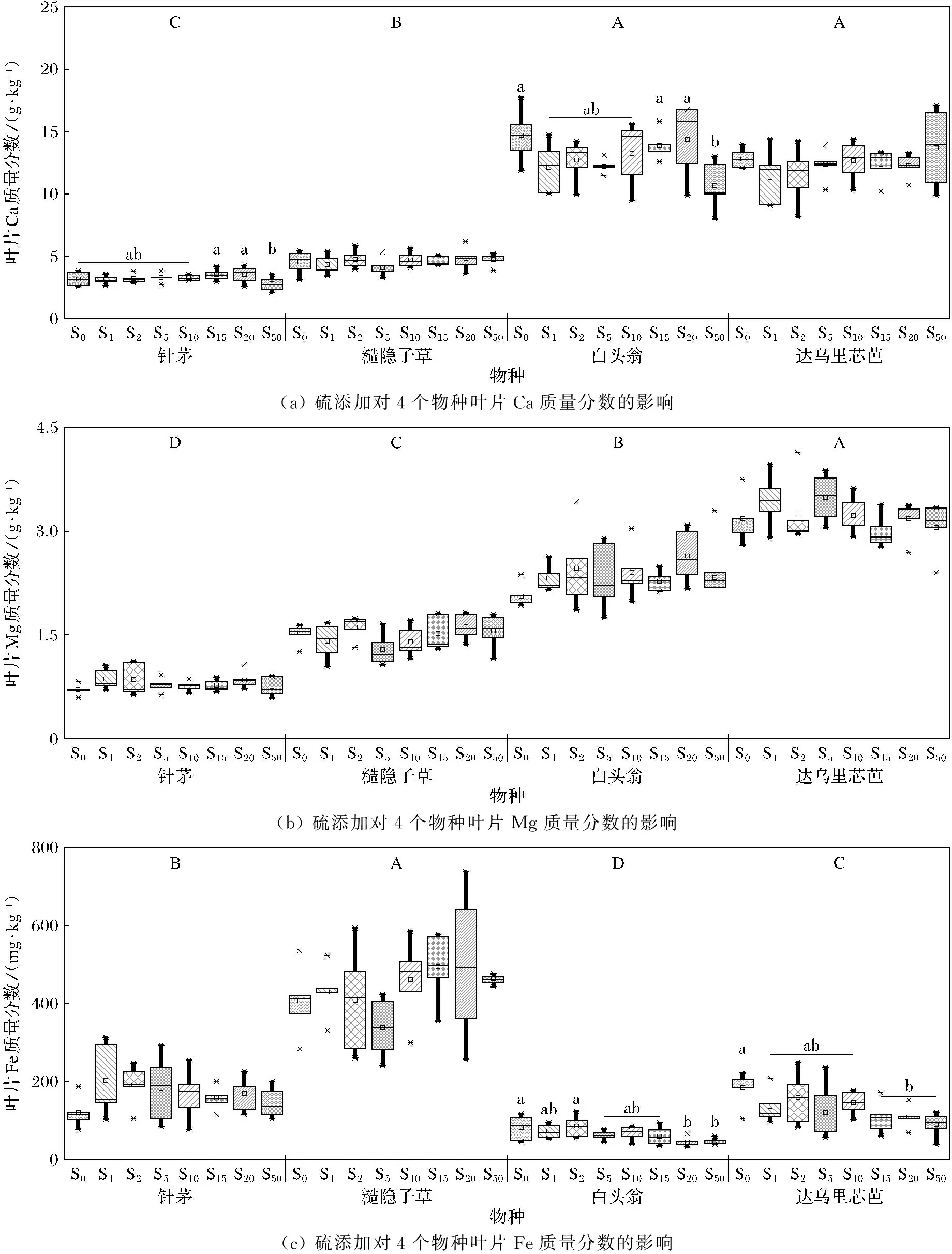
(a) 硫添加对4个物种叶片Ca质量分数的影响(b) 硫添加对4个物种叶片Mg质量分数的影响(c) 硫添加对4个物种叶片Fe质量分数的影响
2.2 群落水平叶片和根系中微量元素质量分数对硫添加响应
由图2可知,植物群落水平根系Ca、Fe、Mn质量分数高于叶片,其中根系Fe质量分数是叶片的几倍至数十倍;而根系和叶片Mg质量分数比较接近。群落水平叶片Ca质量分数对硫处理的响应不显著,而根系Ca质量分数在硫添加量为50 g·m-2·a-1时显著下降(图2(a));群落水平叶片和根系Mg质量分数对硫处理的响应均不显著(图2(b))。群落水平叶片Fe质量分数在中量硫添加量(10 g·m-2·a-1)下最高,低量和高量硫添加下相对较低,根系Fe质量分数对硫添加无显著响应(图2(c));硫添加显著增加群落叶片和根系Mn质量分数(图2(d))。
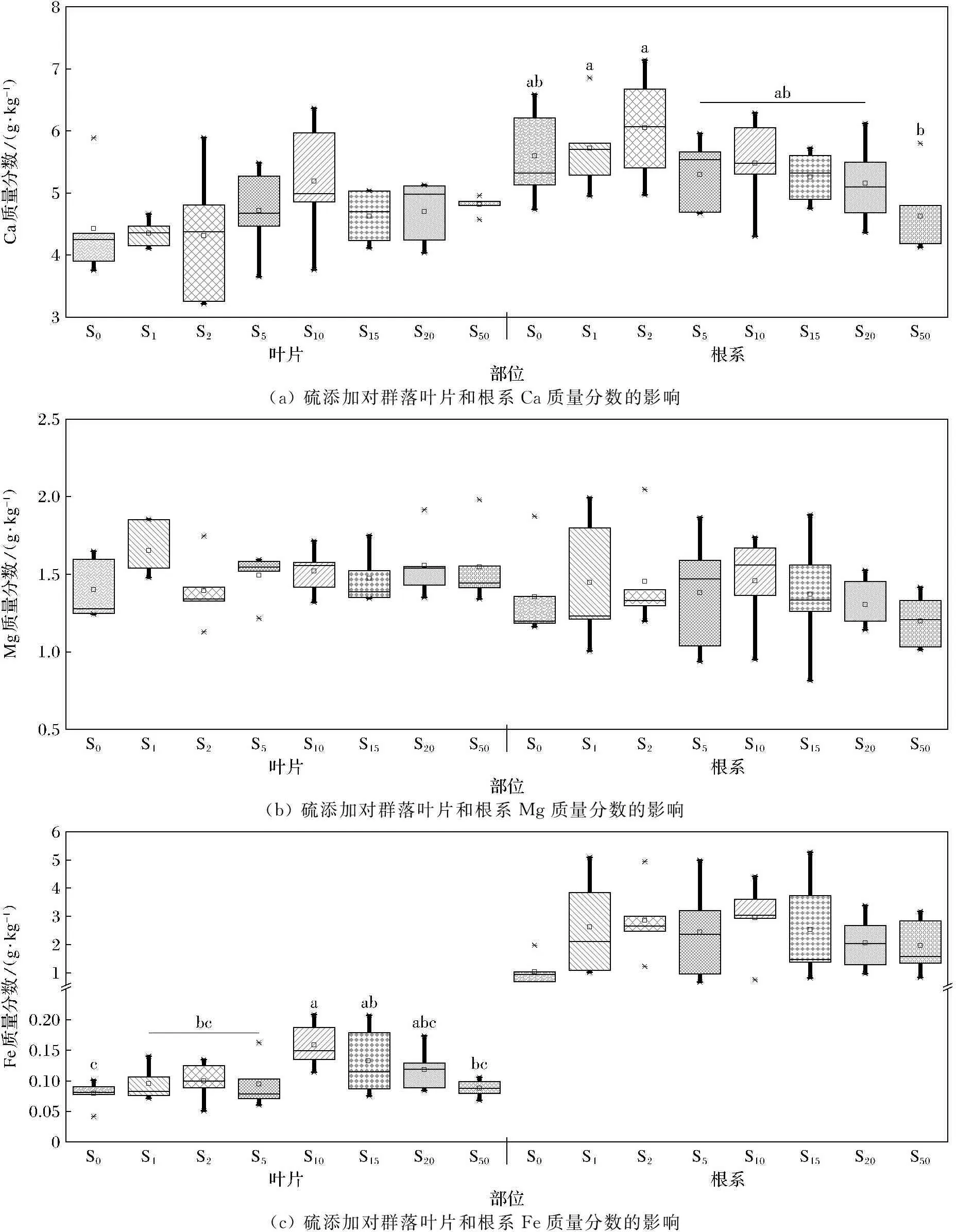
(a) 硫添加对群落叶片和根系Ca质量分数的影响(b) 硫添加对群落叶片和根系Mg质量分数的影响(c) 硫添加对群落叶片和根系Fe质量分数的影响
2.3 植物群落叶片和根系中微量元素质量分数与土壤性质的关系
从表1可知,植物群落叶片Ca、Fe质量分数与土壤pH值、交换性Al、交换性Ca、Mg、DTPA-Fe、DTPA-Mn无显著相关性;叶片Mg质量分数与土壤交换性Mg显著正相关;叶片Mn质量分数与土壤pH值、交换性Ca显著负相关,与交换性Al、DTPA-Fe、DTPA-Mn显著正相关。植物群落根系Ca质量分数与土壤pH值显著正相关,与土壤交换性Al和DTPA-Mn显著负相关;根系Mg、Fe质量分数与土壤性质间无显著相关性;根系Mn质量分数与土壤pH值显著负相关,与交换性Al、DTPA-Fe、DTPA-Mn显著正相关。
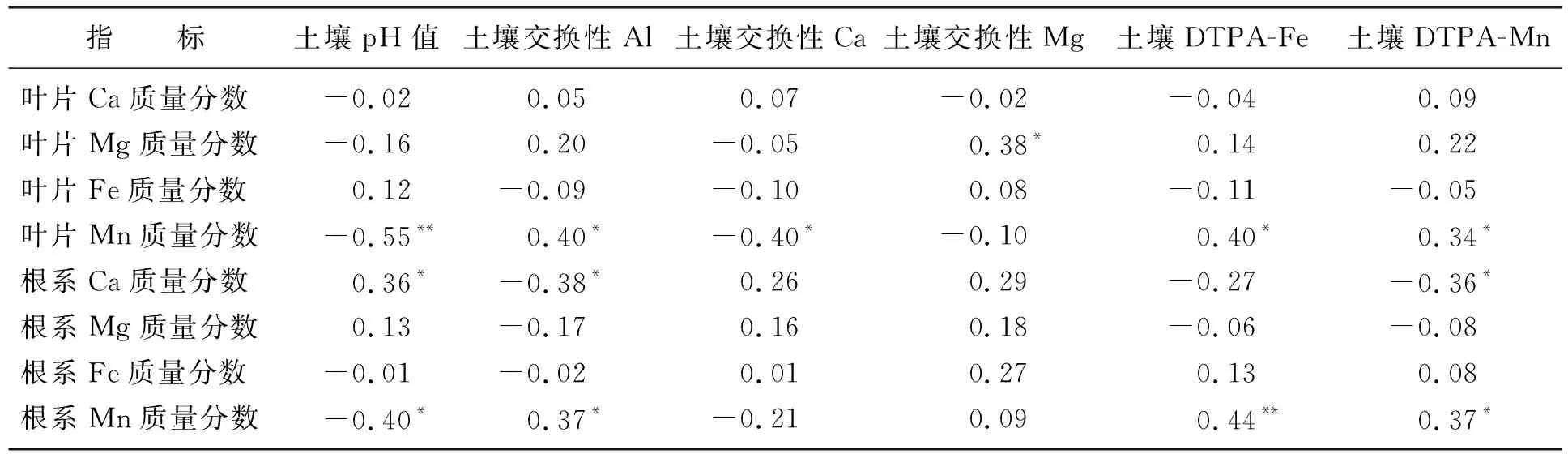
表1 植物群落水平叶片和根系中微量元素质量分数与土壤性质间的Pearson相关系数
3 讨 论
3.1 硫添加对草甸草原4个物种叶片中微量元素质量分数的影响
本研究中,不同植物物种叶片Ca、Mg、Fe、Mn质量分数存在显著差异,假设①成立。根据高等植物营养学原理,不同功能群植物物种对中微量元素的吸收存在差异,主要是植物发育过程中代谢途径以及养分在植物器官中的分配存在差异所致[18]。4个物种的叶片中微量元素质量分数对硫添加的响应具有显著的物种特异性。针茅和白头翁叶片Ca质量分数在硫添加量低于20 g·m-2·a-1处理时无显著变化,而在硫添加量为50 g·m-2·a-1处理下才显著降低,说明只有当土壤离子变化超过植物耐受范围时才会引起植物的响应,这与假设②一致。亚热带森林的酸添加试验表明,中低水平的酸化能够显著增加杉木Ca和Mg质量分数,但在高水平酸化处理下出现下降趋势[19]。在高硫处理下针茅和白头翁叶片Ca质量分数显著降低主要原因是土壤酸化增加了钙镁离子的淋失[20],这一结果与中国北方半干旱草原和欧洲草原的研究结果一致[21-22]。半干旱草原的研究发现,猪毛蒿和风毛菊2个杂类草叶片Ca质量分数同时受到土壤酸化的抑制[21],这种Ca吸收的降低可能减少植物抵抗环境胁迫的能力[23-24]。不同物种对土壤酸化的差异性响应,可能归因于植物根系形态特征和根系深度分布模式的不同导致植物对土壤中量元素变化的响应不同[21]。硫添加对4个物种的叶片Mg质量分数均无显著影响,与西北黄土高原的一项研究结果一致,土壤酸化下多年生C4禾草的地上组织Mg质量分数也表现出较好的稳定性[25]。这一结果与假设②不一致,由于草甸草原黑钙土富含中量元素,短期的硫添加对植物Mg吸收的影响不大。
本研究发现,两个杂类草物种白头翁和达乌里芯芭叶片Fe质量分数在硫添加下均显著降低,而禾本科物种针茅和糙隐子草Fe质量分数无显著变化(图1(c)),4个物种叶片Mn质量分数随硫添加量的增加均显著增加(图1(d))。这种叶片Fe吸收的差异主要由于杂类草Mn、Fe元素吸收表现出显著的拮抗作用,Mn与Fe竞争相同的吸收位点,较高的Mn积累抑制杂类草对Fe的吸收[26]。杂类草的Fe的吸收主要由还原反应介导,即根细胞内的铁还原酶将Fe3+还原成Fe2+,然后在铁运输蛋白的作用下吸收Fe2+进入根内并伴随着Mn2+离子的运输[18,27]。土壤酸化引起的土壤有效Mn质量分数增加导致Mn2+直接与Fe2+竞争,造成杂类草叶片Mn积累抑制Fe吸收[15]。禾本科植物可同时吸收Fe3+和Fe2+,进而这种拮抗作用对禾草叶片地上部Fe吸收影响较杂类草小。本研究表明,不同功能群物种Fe和Mn吸收存在显著差异性,由于杂类草Mn吸收显著抑制Fe吸收,加之Mn的奢侈吸收,可能会影响杂类草的生理生化过程进而导致一些杂类草物种因土壤酸化而在植物群落中的率先丢失。假设3得到实验验证。
3.2 硫添加对草甸草原植物群落叶片和根系中微量元素质量分数的影响
本研究是硫添加实验处理的第2年,尽管针茅和白头翁叶片Ca质量分数对高量硫酸添加(50 g·m-2·a-1)表现出显著响应,但群落水平上叶片Ca质量分数对硫添加并未有显著响应,这可能是由于实验处理时间较短,植物群落尚能抵抗土壤酸化胁迫[28]。根据质量比假说,生态系统过程主要由优势物种的功能特征决定[29],因此本研究中草地植物群落叶片Ca质量分数可能主要受优势物种(羊草和寸草苔生物量占比在60%以上)的影响。与之相反,根系Ca质量分数受到硫添加的抑制,这种叶片和根系的差异性响应,表明植物可能会通过调节不同器官的养分吸收策略进而保持体内的养分相对平衡[30]。另一种可能的原因是,根系与叶片所处的生长环境不同,其对土壤性质变化的敏感性要高于叶片[31]。本研究中,植物群落根系Ca质量分数与土壤pH值显著正相关,与土壤交换性Al显著负相关(表1)。由于Ca离子与Al离子会竞争交换位点,土壤中高量的交换性Ca会帮助植物抵御土壤Al的毒害[32],而硫添加导致的土壤钙淋失可能导致植物根系潜在遭受Al毒害[33]。与Ca相比,植物中Mg质量分数较为稳定,群落叶片和根系Mg质量分数对硫添加均无显著响应,且仅有群落叶片Mg与土壤交换性Mg呈显著正相关(图2(b),表1)。主要原因是短期硫添加并未显著改变土壤交换性Mg质量分数(未发表数据),因此不同物种Mg质量分数未受到硫添加处理的显著影响。在群落水平上,假设②不成立。
Ca和Mg是植物结构组织和基本代谢的基础元素,是有机化合物的组成成分,往往受到更严格的生物调控[34]。而微量元素Fe、Mn的环境敏感性高,生物控制弱,对环境变化的响应更快[30]。本研究中,硫添加可增加植物群落叶片Fe质量分数(图2c)且与土壤理化和DTPA-Fe的相关性很弱,符合土壤和植物元素质量分数之间存在微弱耦合关系的观点[35]。有研究发现,在硫充足的情况下,禾本科植物根系Fe质量分数更易受土壤硫有效性的影响[36],主要是因为Fe和S营养之间存在共同的稳态调节机制[37]。S供应充足时,植物会增加对S的同化和蛋氨酸的合成,同时增加Fe载体的分泌,因此施S可导致植物叶片Fe吸收增加[36]。本研究所选取的草甸草原主要以禾草为优势物种,植物群落叶片Fe质量分数对硫添加的正向响应与禾草的响应(图1(c))趋势一致。群落植物不同器官对Mn元素的吸收受到土壤pH值的调控,与土壤pH值显著负相关,而与土壤交换性Al、DTPA-Mn显著正相关,植物群落地上、地下器官的元素质量分数具有相似的变异模式(图2(d),表1)。多项研究表明,土壤酸化可显著促进草原植物对Mn的奢侈吸收,而酸化导致的植物Mn毒害也是草原植物多样性降低的一个重要机制[11,15,28]。在物种水平和群落水平上,假设③均成立,即土壤酸化促进植物Mn吸收,植物Mn与Fe吸收在一定程度上存在拮抗作用。
4 结 论
草甸草原不同物种对中量元素Ca、Mg和微量元素Fe、Mn的吸收存在显著的差异性,禾草叶片对微量元素Fe、Mn具有较高的富集能力,而杂类草则含有较高量的中量元素Ca、Mg。石灰性草甸草原交换性Ca、Mg质量分数相对较高,不同物种叶片Ca、Mg对短期的硫添加的响应较弱;硫添加显著降低土壤pH值、增加交换性Al质量分数,致使土壤酸化并活化土壤Mn和Fe,因此所有物种叶片Mn质量分数均随硫添加量的增加而显著增加;杂类草叶片Fe质量分数随硫添加量而显著降低,存在显著的锰铁拮抗效应。在群落水平上,根系Ca、Fe、Mn质量分数显著高于叶片,表明草原植物不同器官对中微量元素的富集能力具有显著差异。短期硫添加诱导的土壤酸化对植物群落叶片和根系的Mn质量分数、根系Ca质量分数具有显著影响,对叶片和根系的Mg、Fe质量分数影响相对较小。研究结果有助于评估和预测半干旱草原土壤-植物系统养分循环过程对土壤酸化的响应,为草原生态系统养分管理及可持续利用提供数据支撑。
