UV-B辐射对不同品种(品系)辣椒幼苗光合特性及UVR8表达的影响
2023-12-13陈艳邓昌蓉侯全刚文军琴刘敏邵登魁
陈艳 邓昌蓉 侯全刚 文军琴 刘敏 邵登魁






摘要: 為了解紫外线B(UV-B)辐射对辣椒(Capsicum annuum L.)光合特性及UV-B应答基因UVR8表达量的影响,以青海省主栽的9个辣椒品种(品系)为试验材料,紫外线处理剂量为28.56 kJ/(m2·d),设空白对照(无照射),测定幼苗叶片光合特性、形态指标等14个指标,同时利用实时荧光定量PCR技术检测UVR8基因在各试验材料中的相对表达量。结果表明,与对照相比,处理组辣椒的叶绿素、类胡萝卜素含量总体降低,光合指标总体下降,从而抑制叶片光合作用,总体上UV-B处理对华美105大多数光合指标的影响较大,对乐都长辣椒的影响较小。荧光定量PCR检测结果显示,UV-B处理后,UVR8在华美105中的相对表达量上升幅度最大且与对照间的差异达显著水平。综合分析可知,华美105的大多数指标显著受到抑制,光能利用率降低,因此华美105为UV-B敏感型品种,且UVR8在该品种中的表达水平较高,说明UVR8能够在不耐紫外辐射辣椒品种(品系)中积极响应UV-B胁迫。
关键词: 辣椒;UV-B辐射;光合特性;UVR8
中图分类号: S624.4+3 文献标识码: A 文章编号: 1000-4440(2023)07-1449-11
Effects of UV-B radiation on photosynthetic characteristics and UVR8 expression in different pepper seedlings
CHEN Yan1,2, DENG Chang-rong1,2,3, HOU Quan-gang1,2,3, WEN Jun-qin1,2,3, LIU Min1,2, SHAO Deng-kui1,2,3
(1.Qinghai Academy of Agriculture and Forestry Sciences, Xining 810016, China;2.Qinghai Key Laboratory of Vegetable Genetics and Physiology, Xining 810016, China;3.Laboratory for Research and Utilization of Germplasm Resources in Qinghai Tibet Plateau, Xining 810016, China)
Abstract: In order to understand the effects of ultraviolet B (UV-B) radiation on photosynthetic characteristics and UV-B response gene UVR8 expression of capsicum (Capsicum annuum L.), nine varieties (lines) of capsicum in Qinghai province were selected as test materials, the treatment dose was 28.56 kJ/(m2·d), and blank control was set. A total of 14 indices such as photosynthetic characteristics and morphological indices of seedling leaves were determined. Meanwhile, the expression level of UVR8 was detected by real-time fluorescence quantitative PCR. The results showed that compared with the control, the chlorophyll and carotenoid contents in the treatment group were generally reduced, and the photosynthetic characteristics generally decreased, which inhibited the photosynthesis of leaves. Overall, UV-B treatment had a greater effect on most photosynthetic indices of Huamei 105, and a smaller effect on Ledu long pepper. The results of fluorescence quantitative PCR showed that the relative expression of UVR8 in Huamei 105 increased the most after UV-B treatment and was significantly higher than that in the control group. In conclusion, most indices of Huamai 105 were significantly inhibited, and the utilization rate of light energy was decreased. Therefore, Huamei 105 was a UV-B sensitive variety, and UVR8 was expressed at a high level in this variety, indicating that UVR8 could actively respond to UV-B stress in the varieties that were not resistant to ultraviolet radiation.
Key words: chili;UV-B radiation;photosynthetic characteristics;UVR8
辣椒(Capsicum annuum L.)是茄科辣椒属一年或多年生草本植物,是全球广泛栽培的蔬菜作物,也是中国第一大果菜类园艺作物[1]。辣椒果实中富含维生素、矿物质和生物碱等多种营养物质,在食品、医药等行业有着广泛的用途[2]。辣椒作为青海省第一大设施蔬菜作物,常年栽培面积约为7 000 hm2,主栽品种有乐都长辣椒和陇椒等[3]。
青藏高原地区海拔高,紫外线辐射强。植物叶片是光的靶器官,受到自然光中紫外线B(UV-B)的辐射后,由于UV-B的能量较高,因此不能完全穿透叶片,使得叶片内部受到损伤[4]。有研究发现,由于UV-B促使叶绿素快速分解,使其光能的吸收率、转化率下降[5-7],促使光合作用减弱,而光合作用是植物獲取能量的直接来源。众多研究结果表明,UV-B增强会使多数作物的光合作用减弱,表现为光同化量下降、生长缓慢[8],使得光合作用受到抑制。1995年以来,随着全球气候变化和工业排放量的增多,臭氧层遭到破坏,使得地球受到更强的紫外线辐射,尽管90%UV-B被臭氧层吸收,但仅10%UV-B到达地球表面也会对植物生长产生很大影响[9],并对高原地区的农作物生产造成较大影响。因此,在高原地区开展抗UV-B辐射劣变及其主效基因研究具有比较重要的科学意义。本研究拟通过对不同耐UV-B辐射辣椒材料幼苗期叶片的光合指标进行检测,并借助统计学分析与关键基因表达特性分析,初步筛选具有不同抗紫外线能力的材料,以期为辣椒相关抗性研究体系的建立与资源评价提供参考。
1 材料与方法
1.1 试验材料与试验设计
试验地点为青海大学农林科学院园艺创新基地,供试辣椒材料为QB2016大皱、Yxj2013-16、航椒2号、B1401、青线椒2号、航椒6号、乐都长辣椒、华美105、YTS2013-13,均由青海大学农林科学院园艺研究所提供。
2022年3月初播种,采用72孔穴盘育苗,9个辣椒品种(品系)各育苗72株。待幼苗长至6叶1心期,取长势一致的壮苗移入有UV-B灯的温室中进行处理,设空白对照(无照射)。2022年5月27日开始进行UV-B辐射处理;2022年6月1日,待各辣椒品种(品系)叶片表观出现锈斑时,试验结束,共处理6 d。处理方法:在辣椒植株上方水平挂架上悬挂UV-B灯(26 W,光谱为275~380 nm),用乙酸纤维素膜滤去短波辐射,每日UV-B辐射时间为8 h(9:00-17:00),紫外线强度为99.01 μW/cm2;在UV-B照射期间调整UV-B灯挂架与植株生长面之间的高度使其恒定,对照小区无辐射。用紫外辐照传感器(杭州虹谱光色科技有限公司产品)测定UV-B的辐射强度。
1.2 测定项目及方法
1.2.1 形态及生长指标的测定 随机取各辣椒品种(品系)其中1株于处理第3 d对其叶片进行拍照,且在处理末期第6 d再进行拍照,对各辣椒品种(品系)叶片的表观形态进行观察;各辣椒品种(品系)选整体长势一致的3株植株进行株高、茎粗的测定。
1.2.2 光合参数的测定 选择各辣椒品种(品系)整体长势一致的3株植株,并选择完全展开且能充分补光的叶片,采用Li-6400光合系统测定仪测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等。叶室温度为(25±1) ℃,光量子通量密度(PFD)为100 μmol/(m2·s),CO2浓度为(750±10) μmol/(m2·s),相对湿度(RH)为60%~70%。测定相同叶片的荧光参数,使用荧光叶室测定如下叶绿素荧光参数:初始荧光(F0)、最大荧光(Fm)、暗适应30 min下PSⅡ最大光化学效率(Fv/Fm)、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)。
1.2.3 生理指标的测定 叶绿素、类胡萝卜素含量用95%乙醇提取并测定[10],均为鲜质量含量。
1.2.4 RNA提取和实时荧光定量PCR 参照边海燕等[11]的方法提取RNA,将提取的RNA和合成的cDNA于-20 ℃保存备用。本研究以辣椒UBI3为内参基因[12],根据从美国国家生物技术信息中心(NCBI)网站(https://www.ncbi.nlm.nih.gov)上获得的番茄SlUVR8基因编码序列(CDS)设计引物,引物用Primer 5.0软件设计,由北京奥科鼎盛生物科技有限公司合成,本研究所用的相对定量PCR引物序列见表1,基因相对表达量的计算用2-△△Ct法[13-14]。
1.3 数据处理与分析
试验数据的整理及制图用Excel 2010、Origin 2019,数据统计分析用SPSS 21。
2 结果与分析
2.1 UV-B辐射对不同品种(品系)辣椒幼苗期形态特征的影响
2.1.1 UV-B辐射对辣椒叶片的影响 由图1可以看出,UV-B辐射[28.56 kJ/(m2·d)]对幼苗期辣椒叶片造成了不同程度的损伤。经观察发现,处理第3 d时,华美105叶片表面出现紫色锈斑,处理第6 d时,华美105叶片出现卷曲枯萎的现象。9个辣椒品种(品系)叶片表面在处理末期(第6 d)均表现出不同程度损伤,其中华美105的损伤最严重,乐都长辣椒无明显变化。上述结果表明,用28.56 kJ/(m2·d) UV-B对辣椒进行辐射处理,会对不同辣椒品种(品系)幼苗期的叶片造成不同程度的影响。
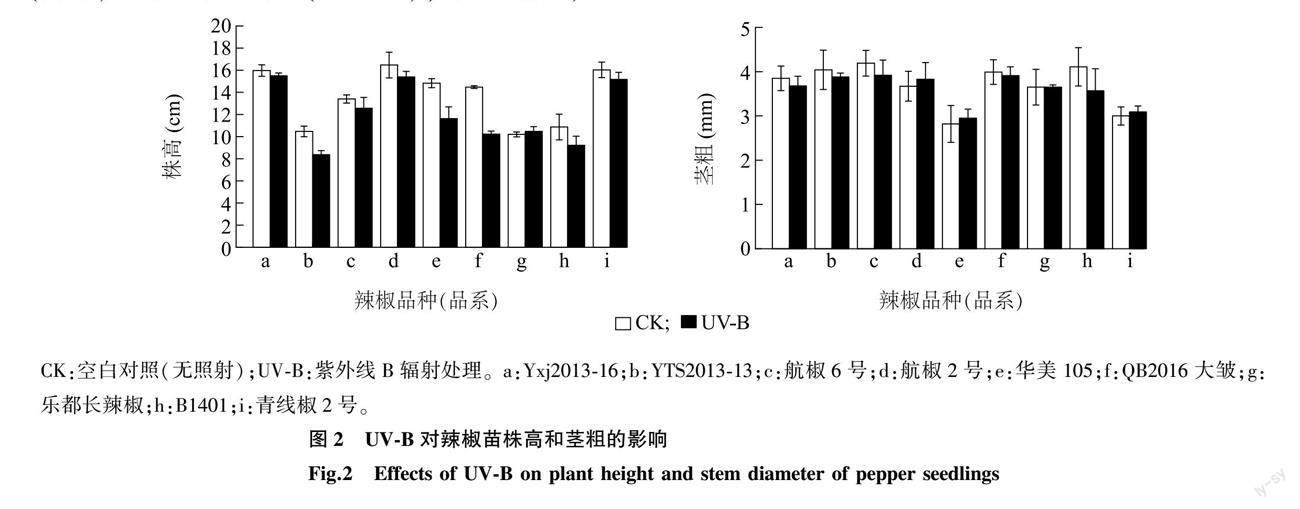
2.1.2 UV-B辐射对辣椒生长的影响 由图2可以看出,补增UV-B辐射对幼苗期辣椒株高整体上表现出不同程度的影响,除乐都长辣椒外,其余辣椒品种(品系)的株高与CK相比均出现受抑制现象,其中QB2016大皱的被抑制现象最明显,其株高显著降低了29.30%。在对辣椒茎粗的影响方面,除了乐都长辣椒、航椒2号、华美105和青线椒2号外,其余辣椒品种(品系)的茎粗与CK相比均表现出变小的特征。表2结果显示,与CK相比,UV-B处理显著减少了YTS2013-13、华美105、QB2016大皱的株高(P<0.05),但对其他辣椒品种(品系)无显著影响。与CK相比,UV-B处理对各辣椒品种(品系)的茎粗无显著影响。方差分析结果表明,经UV-B处理后各辣椒品种(品系)的株高、茎粗相较于CK的变化量间存在显著差异。
2.2 UV-B辐射对不同品种(品系)辣椒幼苗光合特性的影响
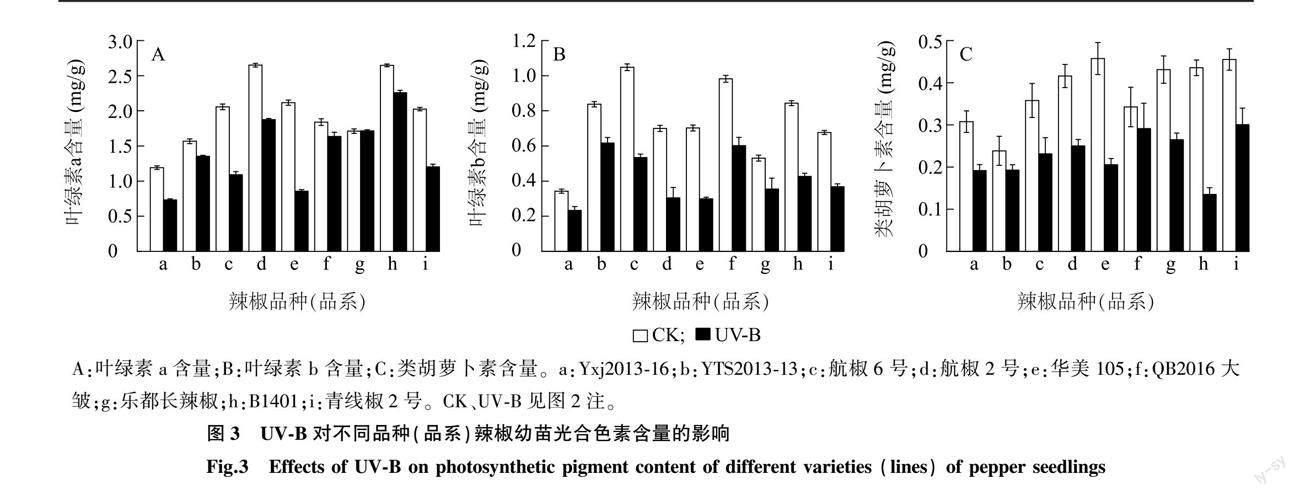
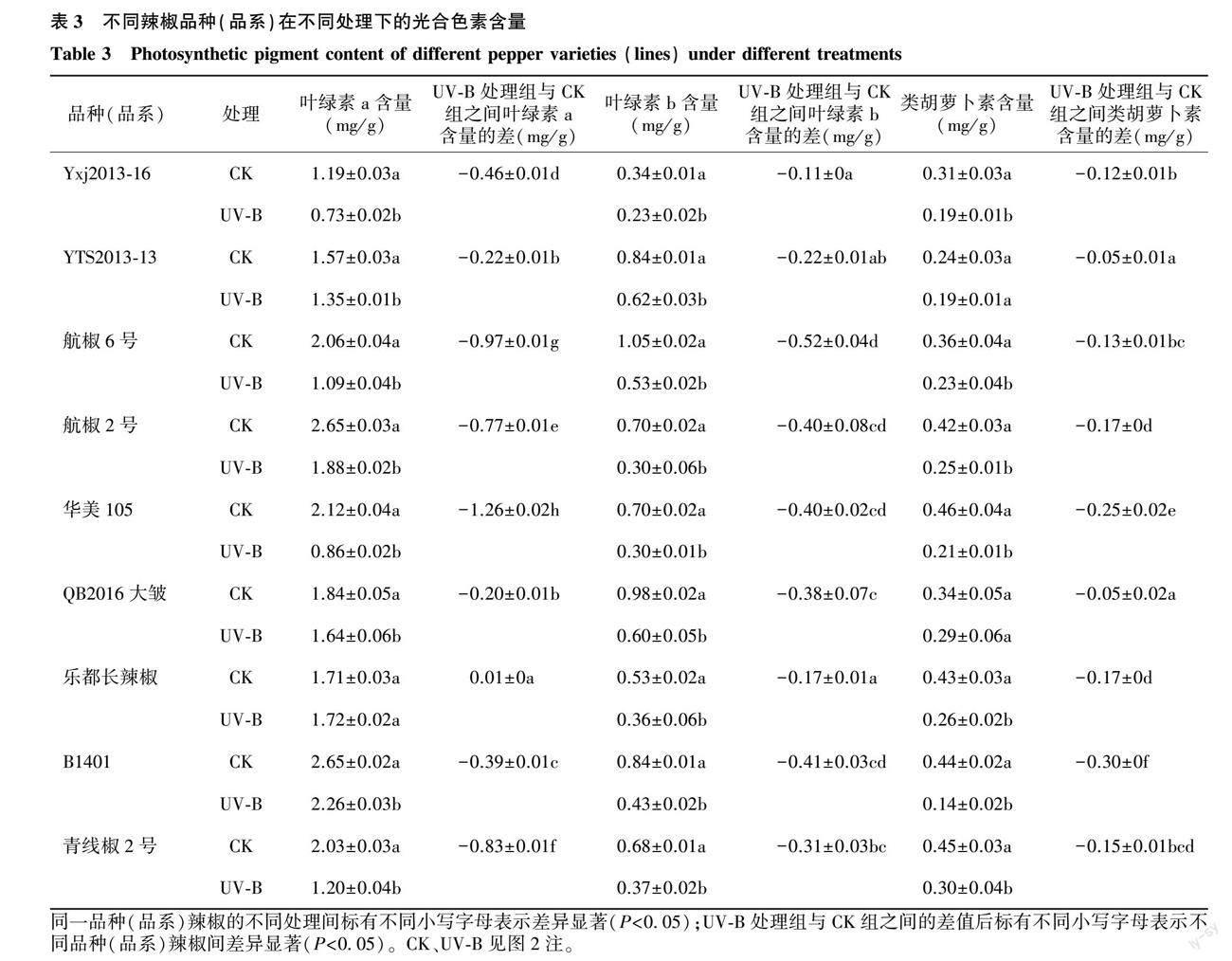
2.2.1 UV-B辐射对不同品种(品系)辣椒幼苗光合色素含量的影响 由图3可以看出,在补增UV-B处理下,除乐都长辣椒外,各辣椒品种(品系)的叶绿素a、叶绿素b、类胡萝卜素含量都低于CK。叶绿素a含量降幅最大的辣椒品种(品系)是华美105,较CK下降了59.43%,乐都长辣椒的叶绿素a含量较CK略有升高(图3A);在UV-B处理下,与CK相比,叶绿素b含量降幅最大的是华美105和航椒2号,都下降了57.14%,叶绿素b含量降幅最小的为YTS2013-13,下降了26.19%(图3B);在UV-B处理下,与CK相比,类胡萝卜素含量降幅最小的是QB2016大皱,较CK降低了14.71%,降幅最大的是B1401,为68.18%(图3C)。表3的t检验结果显示,与CK相比,UV-B辐射对乐都长辣椒的叶绿素a含量无显著影响,但显著降低了其他8個辣椒品种(品系)的叶绿素a含量(P<0.05);与CK相比,UV-B辐射显著降低了各辣椒品种(品系)的叶绿素b含量;与CK相比,UV-B处理显著降低了除QB2016大皱、YTS2013-13外的其他辣椒品种(品系)的类胡萝卜素含量。方差分析结果表明,经UV-B处理的各辣椒品种(品系)叶绿素a、叶绿素b和类胡萝卜素含量与CK的差值间有显著差异。
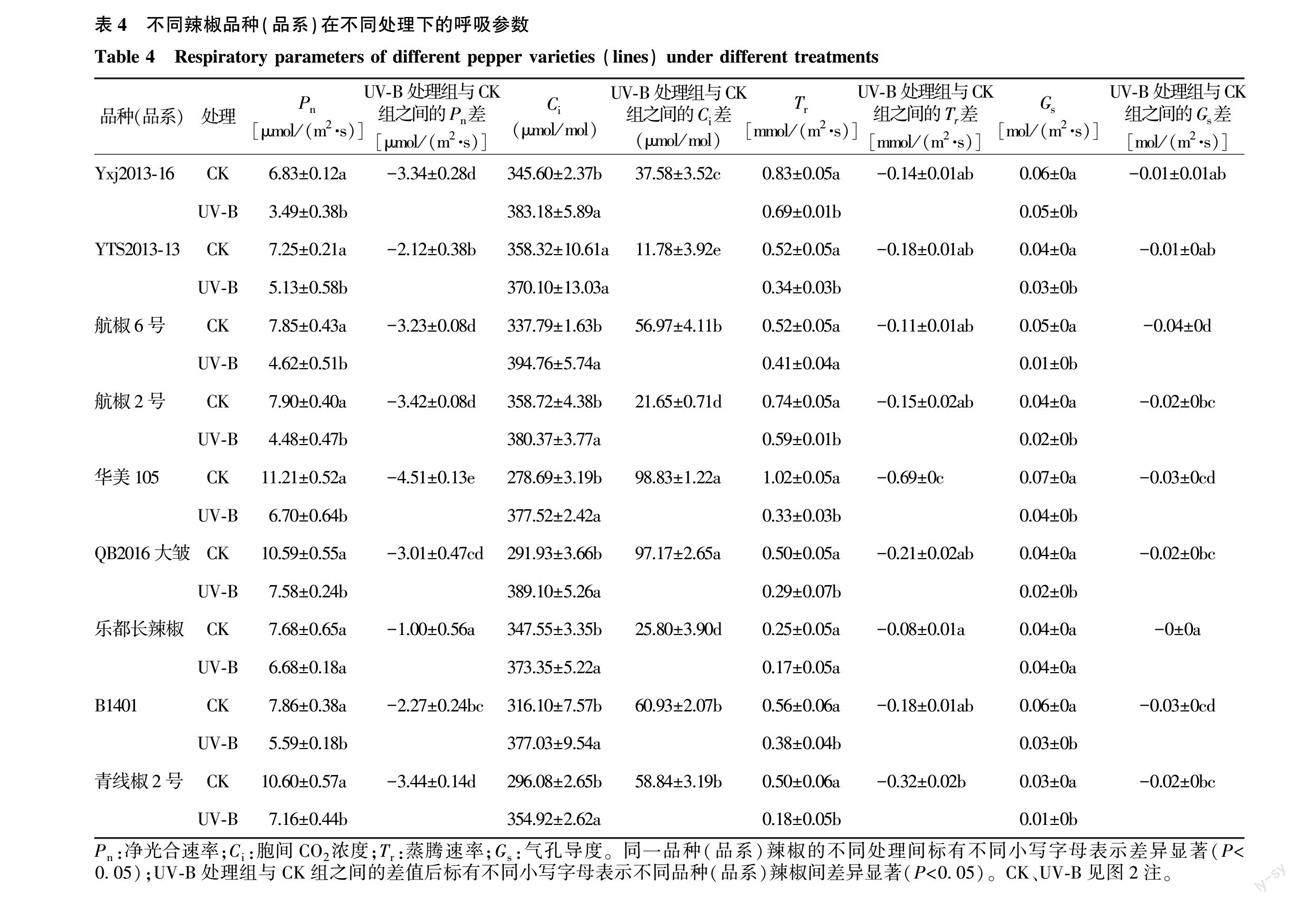
2.2.2 UV-B辐射对不同品种(品系)辣椒幼苗呼吸参数的影响 经UV-B辐射后,与CK相比,各辣椒品种(品系)叶片的净光合速率、气孔导度、蒸腾速率均表现为下降趋势,但胞间CO2浓度却表现为上升趋势(图4)。经UV-B处理后,与CK相比,乐都长辣椒的净光合速率降幅最小,为13.02%,Yxj2013-16的净光合速率降幅最大,达48.90%(图4A)。经UV-B处理后,与CK相比,华美105、青线椒2号的叶片蒸腾速率降幅较大,其中华美105的蒸腾速率较CK下降了67.65%(图4B)。在胞间CO2浓度方面,与CK相比,华美105的胞间CO2浓度上升幅度最大,为35.46%(图4C)。就气孔导度而言,与CK相比,乐都长辣椒的变化幅度最小,航椒6号的变化幅度最大,航椒6号的气孔导度较对照下降了80.00%(图4D)。由表4看出,与CK相比,UV-B辐射对乐都长辣椒的净光合速率、气孔导度无显著影响,但显著降低了其他8个辣椒品种(品系)的净光合速率、气孔导度(P<0.05);与CK相比,UV-B处理对YTS2013-13的胞间CO2浓度无显著影响,但显著增加其他几个辣椒品种(品系)的胞间CO2浓度;与CK相比,UV-B处理对乐都长辣椒、航椒6号的蒸腾速率无显著影响,却显著降低了其余辣椒品种(品系)的蒸腾速率。方差分析结果表明,经UV-B处理的各辣椒品种(品系)净光合速率、气孔导度、蒸腾速率、胞间CO2浓度与CK的差值间存在显著差异。
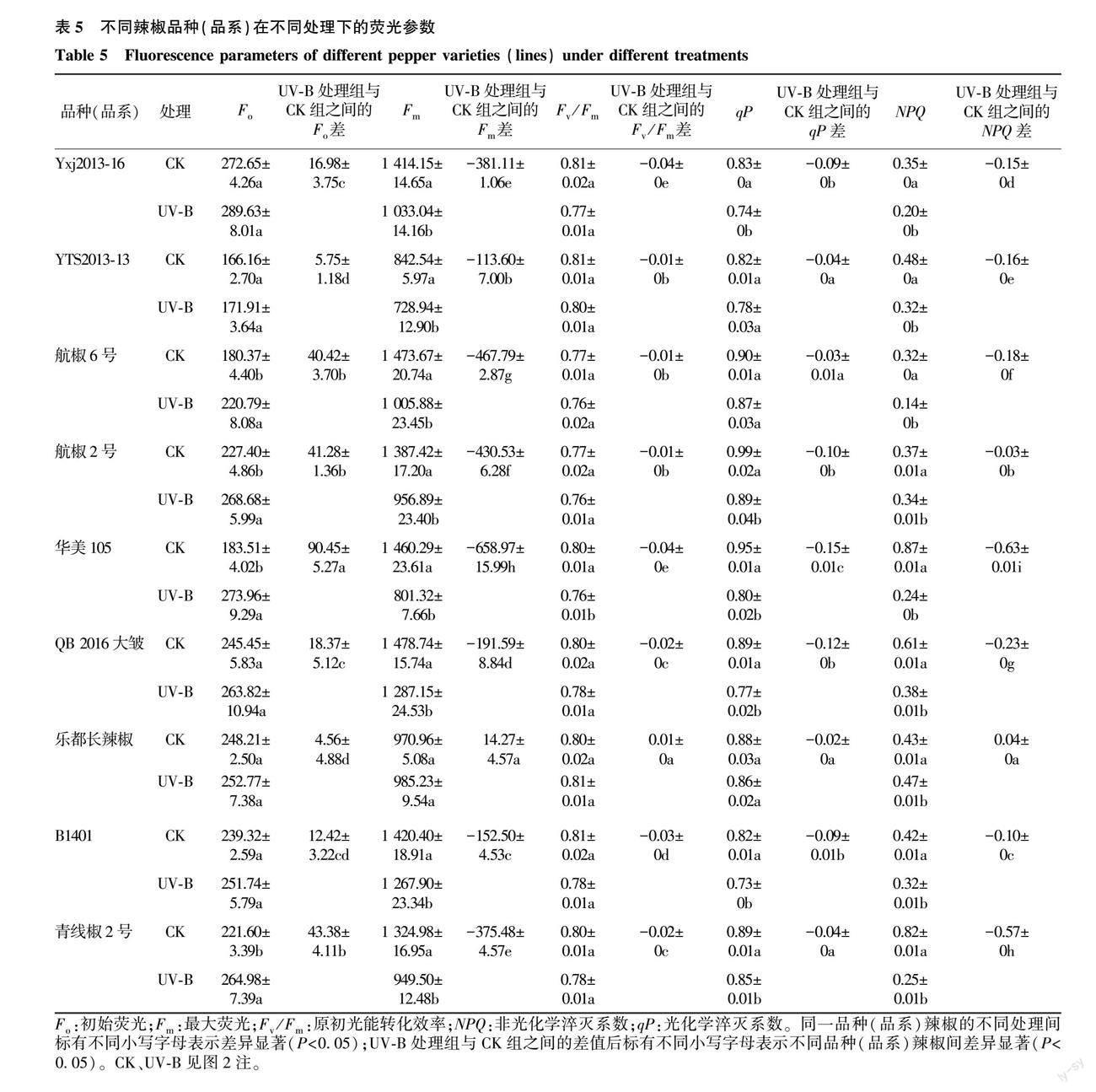
2.2.3 UV-B辐射对不同品种(品系)辣椒幼苗荧光参数的影响 经UV-B辐射处理后,与CK相比,所有参试辣椒品种(品系)的初始荧光均较高,其中乐都长辣椒的变幅最小,华美105的变幅最大,比CK高49.29%(图5A)。经UV-B辐射处理后,与CK相比,乐都长辣椒的Fm变幅最小,华美105的Fm变幅最大,比CK低45.13%(图5B)。除乐都长辣椒外,各辣椒品种(品系)的原初光能转化效率表现为CK>UV-B处理,其中UV-B处理对华美105的影响显著,其他辣椒品种(品系)在处理后与CK间的差异不显著(图5C、表5)。在UV-B处理下,各辣椒品种(品系)的光化学淬灭系数整体上表现为CK>UV-B处理,UV-B对华美105的qP影响最大,较CK降低了15.79%(图5D)。除乐都长辣椒外,其他辣椒品种(品系)的非光化学淬灭系数经UV-B处理后均小于CK,其中乐都长辣椒的非光化学淬灭系数与CK相比变幅较小,华美105的非光化学淬灭系数与CK相比变幅最大,比CK低72.41%(图5E)。由表5看出,与CK相比,UV-B辐射处理显著提高了航椒6号、航椒2号、青线椒2号和华美105的Fo(P<0.05),但对其他几个辣椒品种(品系)的Fo无显著影响;乐都长辣椒的Fm经UV-B处理后与CK相比无显著变化,但其他8个辣椒品种(品系)的Fm却显著降低;与CK相比,UV-B处理对除华美105外的其他各辣椒品种(品系)的Fv/Fm无显著影响;与CK相比,UV-B处理显著降低了Yxj2013-16、航椒2号、华美105、QB2016大皱、B1401、青线椒2号的qP;与CK相比,UV-B处理显著降低了大多数辣椒品种(品系)的NPQ。方差分析结果表明,经UV-B处理前后,总体上各辣椒品种(品系)的Fo、Fm、qP、NPQ相较于CK的变化量之间存在显著差异,UV-B处理对华美105荧光参数的影响最明显,对乐都长辣椒各个指标的影响较小。
2.3 UV-B辐射对辣椒幼苗紫外相关基因UVR8表达量的影响
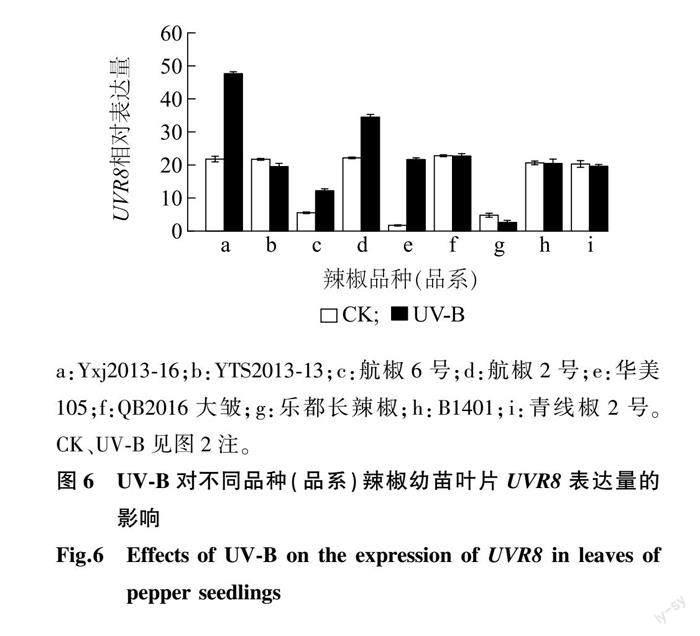
对9个辣椒品种(品系)的叶片进行UV-B辐射处理后,对其叶片中UVR8的相对表达量进行检测分析。如图6所示,与CK相比,UV-B辐射处理的部分辣椒品种(品系)的UVR8相对表达量表现出上调趋势。经UV-B辐射处理后,与CK相比,华美105、Yxj2013-16叶片中的UVR8相对表达量增幅较大,其中华美105的UVR8相对表达量较CK增加了1 158.14%,而乐都长辣椒、YTS2013-13的UVR8相对表达量较CK降低。由表6可以看出,UV-B辐射显著增加了Yxj2013-16、华美105的UVR8相对表达量(P<0.05),除了航椒2号、航椒6号以及青线椒2号,UV-B处理对其他几个辣椒品种(品系)的UVR8相对表达量有显著影响;方差分析结果表明,经UV-B处理后,各辣椒品种UVR8相对表达量相较于CK的变化量之间存在显著差异。
3 讨论
UV-B辐射对植物有强烈的负面效应,会使植物体内蛋白质、叶绿体发生损伤,并能抑制植株生长(包括农作物减产等)[15-16]。叶片作为光合作用的靶器官,受到自然光中UV-B的辐射后会使其内部生理代谢过程受到影响[4]。叶片受损伤严重时,还会出现黄斑、褐变、坏死等现象[17-18]。在本研究中,除了乐都长辣椒在处理前后无明显变化外,其他辣椒品种(品系)经UV-B辐射后叶片表面均出现不同程度的紫色锈斑,这与前人的研究结果[17-18]一致。株高作为植物所有生物学特性中最容易被观察的性状,经常被作为抗性指标进行相关研究。前人在对杂草[19]、大豆[20]、小麦[21]等研究中发现,高剂量紫外线辐射均可抑制植物株高生长。在本研究中,绝大多数参试辣椒品种(品系) 的株高经UV-B处理后有不同程度的下降,与前人研究结果[19-21]一致。
UV-B辐射除了能改变株高、叶片表观形态发生外,还能影响植物光合系统[22]。UV-B辐射对于植株光合作用的抑制一方面直接通过损伤内囊体膜的完整性[23]、抑制光系统Ⅱ的活性或者减少CO2固定和色素含量来实现[24];另一方面,UV-B可促使叶片气孔关闭以抑制气体交换,引起气孔导度下降,进而间接影响到光合作用[15]。叶绿素荧光参数是植物光合作用的内在探针,通过分析叶绿素荧光动力学参数可以揭示植物光能吸收、转化、传递等过程的生理状态[25]。韩雯等[26]研究发现,短时间的UV-B辐射对拟南芥叶绿素荧光特性有影响,原初光能转化效率、光化学淬灭系数等指标降低,NPQ呈上升趋势。在本研究中,与CK相比,UV-B处理明显降低了参试辣椒品种(品系)的光合色素(叶绿素a、叶绿素b、类胡萝卜素)含量和气孔导度,从而直接或间接影响植株的光合作用,这与陈慧泽等[15]的研究结论一致。张玉红等[27-28]研究发现,经UV-B辐射后的植株,其原初光能转化效率、光化学淬灭系数等降低,这与本研究中9个辣椒品种(品系)的Fv/Fm、qP经UV-B处理后的变化基本一致。在本研究中,与CK相比,UV-B处理后的辣椒叶片NPQ下降,这与祁虹等[29]对棉花的研究结论相反,可能是因为本研究中紫外线处理剂量较高,导致光系统受损。
UVR8是UV-B辐射后的关键应答基因[30]。前人研究发现,经UV-B辐射后,茄子幼苗子叶、白桦树叶片中的UVR8相对表达量均上调,说明UVR8能够积极响应UV-B胁迫[31-32]。在本研究中,华美105的UVR8相对表达量经UV-B处理后上升幅度最大,说明其感受UV-B的能力最强,即华美105的抗UV-B能力可能较弱,华美105的大部分光合指标也受到了抑制,导致其光能利用率降低,推测此品种为UV-B敏感品种。
4 结论
综上,辐射剂量為28.56 kJ/(m2·d)的UV-B处理通过损伤辣椒幼苗叶片生物膜,降低了辣椒叶片中叶绿素含量、类胡萝卜素含量以及大多数呼吸参数和荧光参数,使得辣椒幼苗的光能利用率降低,从而抑制辣椒幼苗叶片的光合作用。此外,UVR8相对表达量表达模式分析结果表明,用28.56 kJ/(m2·d) UV-B处理后,UVR8的相对表达量在华美105叶片中的上升幅度最大,因此28.56 kJ/(m2·d) UV-B辐射在一定时间内也能影响该基因的转录表达。本研究对高原地区辣椒幼苗期增加紫外线辐射,初步探索了UV-B处理后辣椒幼苗期光合特性、关键应答基因相对表达量的变化,为高原地区抗紫外线品种筛选和资源创新提供了研究思路和方向,为抗紫外线劣变育种技术创新与新品种选育奠定了技术基础。
参考文献:
[1] 付秋实,李红岭,崔 健,等. 水分胁迫对辣椒光合作用及相关生理特性的影响[J]. 中国农业科学,2009,42(5):1859-1866.
[2] 彭书练. 辣椒功能成分的综合提取技术研究[D]. 长沙:湖南农业大学,2007.
[3] 叶 超. 青海循化线辣椒带富撒拉人家[N]. 西部时报,2008-11-25(13).
[4] 钟 楚,陈宗瑜,王 毅,等. UV-B辐射对植物影响的分子水平研究进展[J]. 生态学杂志,2009,28(1):129-137.
[5] 李 元,王勋陵. 紫外辐射增加对春小麦生理、产量和品质的影响[J]. 环境科学学报,1998(5):58-63.
[6] 杨景宏,陈 拓,王勋陵. 增强紫外线B辐射对小麦叶绿体膜组分和膜流动性的影响[J]. 植物生态学报,2000(1):102-105.
[7] 林文雄,吴杏春,梁义元,等. UV-B辐射胁迫对水稻叶绿素荧光动力学的影响[J]. 中国生态农业学报,2002,10(1):8-12.
[8] TEVINI M,TERAMURA A H. UV-B effects on terrestrial plants[J]. Photochemistry and Photobiology, 1989, 50(4): 479-487.
[9] 杜照奎. UV-B辐射增强对花生的生理生态影响及其分子机制[D]. 重庆:西南大学,2014.
[10]ARNON D I . Copper enzymes in isolated chloroplasts[J]. Plant Physiology, 1949, 24(1): 1-15.
[11]边海燕,钟启文,黄思杰,等. 干旱胁迫下大蒜果聚糖代谢关键酶基因的表达分析[J]. 分子植物育种,2018,16(20):6770-6776.
[12]赵 敬. 辣椒内参基因的筛选及NBS-LRR类抗病基因同源序列的鉴定[D]. 南京:南京农业大学,2012.
[13]张 宇,周自云,夏鹏国,等.干旱胁迫对柴胡中皂苷合成关键酶基因表达及皂苷含量的影响[J]. 中国中药杂志,2016,41(4):643-647.
[14]LIVAK K J, SCHMITTGEN T. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ct method[J]. Methods,2001,25(4):402-408.
[15]陈慧泽,韩 榕. 植物响应UV-B辐射的研究进展[J]. 植物学报,2015,50(6):790-801.
[16]CALDWELL M M, BJRN L O, BORNMAN J F, et al. Effects of increased solar ultraviolet radiation on terrestrial ecosystems[J]. Journal of Photochemistry and Photobiology B: Biology, 1998, 46(1/2/3): 40-52.
[17]李 俊,牛金文,杨 芳,等. 不同马铃薯品种(系)对增强UV-B辐射的形态响应[J]. 中国生态农业学报,2016,24(6):770-779.
[18]JANSEN M A K, GABA V, GREENBERG B M. Higher plants and UV-B radiation: balancing damage, repair and acclimation[J]. Trends in Plant Science,1998,3(4):131-135.
[19]BARNES P W, FLINT S D, CALDWELL M M. Morphological responses of crop and weed species of different growth forms to ultraviolet-B radiation[J]. American Journal of Botany, 1990,77(10):1354-1360.
[20]LI Y, ZU Y Q, CHEN J J. et al. Intraspecific responses in crop growth and yield of 20 soybean cultivars to enhanced ultraviolet-B radiation under field conditions[J]. Field Crops Research,2002,78(1):1-8.
[21]王传海,郑有飞,何都良,等. 南京地区近地面紫外辐射UV-B强度对小麦生长及产量影响的评估[J]. 农业环境科学学报,2003(2):147-149.
[22]蔡锡安,夏汉平,彭少麟. 增强UV-B辐射对植物的影响[J]. 生态环境,2007(3):1044-1052.
[23]SWARNA K, BHANUMATHI G, MURTHY S. Studies on the UV-B radiation induced oxidative damage in thylakoid photofunctions and analysis of the role of antioxidant enzymes in maize primary leaves[J]. Bioscan,2012,7: 609-610.
[24]CHEN H Z, HAN R. He-Ne laser treatment improves the photosynthetic efficiency of wheat exposed to enhanced UV-B radiation[J]. Laser Physics,2014,24(10): 105602.
[25]耿东梅,单立山,李 毅,等. 土壤水分胁迫对红砂幼苗叶绿素荧光和抗氧化酶活性的影响[J]. 植物学报,2014,49(3):282-291.
[26]韩 雯,韩 榕. 不同时间的 UV-B 辐射对拟南芥幼苗生长的影响[J].植物学报,2015,50(1): 40-46.
[27]张玉红,陈路瑶,刘 彤,等.增补 UV-B 辐射对药用植物黄檗幼苗生长及光合生理影响[J]. 中国农学通报,2018,34(4):76-82.
[28]李雪莹,殷 红,战莘晔,等.添加 UV-B 辐射对灌浆期水稻剑叶光合特性及抗氧化能力的影响[J]. 沈阳农业大学学报,2017,48(3): 271-276.
[29]祁 虹,段留生,王树林,等. 全生育期UV-B辐射增强对棉花生长及光合作用的影响[J]. 中国生态农业学报,2017,25(5): 708-719
[30]刘明雪,孙 梅,王 宇,等. 植物 UV-B 受体及其介导的光信号转导[J]. 植物学报,2012,47(6): 661-669.
[31]张君豪. 茄子UVR8基因的克隆与表达分析[D].上海:上海交通大学,2019.
[32]李曉一,詹亚光,娄晓瑞,等. 白桦BpUVR8基因的序列与表达模式分析[J]. 植物生理学报,2016,52(5): 685-692.
(责任编辑:徐 艳)
