基于高通量定量蛋白组学技术的全脂牛乳和脱脂牛乳蛋白组差异分析研究
2023-12-03董雅静张九凯邢冉冉董立雅潘明飞
董雅静,张九凯,邢冉冉,董立雅,于 宁,潘明飞,逯 刚,,陈 颖,
(1.天津科技大学食品科学与工程学院,天津 300457;2.中国检验检疫科学研究院,北京 100176;3.内蒙古蒙牛乳业(集团)股份有限公司,内蒙古呼和浩特 011517)
蛋白质是牛乳中重要的营养成分,主要包括酪蛋白和乳清蛋白,在机体免疫调节、抗炎和抗氧化等方面发挥着重要的生物活性作用[1-2]。脱脂牛乳是指以生牛乳为原料,经低温预热后高速离心脱脂制成的乳制品,脂肪含量一般低于0.5%[3-4]。由于过多摄入饱和脂肪会增加心脑血管疾病以及高血压等疾病发病风险[5-8],为减少日常饮食中脂肪的摄入,很多人青睐于食用脱脂乳以满足低脂高蛋白的饮食和健康需求。澳大利亚一项关于长期饮用全脂牛乳、低脂牛乳、脱脂牛乳和豆奶产品与心血管疾病死亡率的回顾性研究表明,患病人群以低脂肪含量牛乳代替全脂牛乳可以降低31%~41%心血管疾病死亡率[9]。虽然有很多关于全脂乳和脱脂乳营养功能对比的研究,但在脱除脂肪的同时,对乳中关键营养物质-蛋白质的变化情况研究并不深入。Zorana 等[10]对离心后牛羊乳的上清液和沉淀物分析时发现,离心脱脂会导致乳中蛋白质沉淀,且预热温度和离心力的升高对沉淀产生具有显著性影响。Katarzyna 等[11]对山羊奶研究时发现,脱脂会降低山羊奶总氮含量,但对蛋白质组成无显著性影响。因此全面了解脱脂处理后牛乳全蛋白的变化情况对消费者选购及生产优化具有重要意义。
蛋白组学在乳品分析中具有广泛的应用,从细胞蛋白质表达层面揭示了不同乳制品差异,是乳品营养价值和生物活性分析的有力工具[12-14]。TMT(Tandem Mass Tags)是一种化学标记技术,用于基于质谱(Mass Spectra,MS)的生物大分子(例如蛋白质、多肽和核酸)的定量和鉴定。该技术利用等量同位素试剂对不同样品中肽段N 端进行标记,质谱检测过程中标记试剂碎裂释放报告基团(Reporter Group),根据报告基团的信号强度可以对样本蛋白相对丰度进行检测[15-16]。TMT 作为一种高通量定量蛋白组学技术,具有蛋白质覆盖面广,结果重现性好等优势,主要用于不同样品间差异蛋白分析[17-19]。Li 等[20]应用TMT 技术对不同热加工条件下骆驼乳蛋白图谱进行分析,共鉴定到807 个差异蛋白,涉及RNA 运输、PPAR 信号通路等生物学过程,阐明了热处理对骆驼乳蛋白影响。在牛乳相关研究中,Li 等[13]对不同品种水牛乳进行分析,共鉴定到62 个差异蛋白,大多数蛋白与免疫活性相关,为水牛品种间的营养差异分析提供了依据。Zhang 等[21]通过对不同热处理奶牛乳清蛋白中营养成分进行分析,阐明了热处理对牛乳营养成分的影响,拓展了对乳清蛋白热敏程度的认知。然而,目前还未发现利用TMT 标记定量技术探究脱脂对牛乳蛋白影响的研究。
因此,本研究采用TMT 标记定量蛋白组学技术对全脂牛乳和脱脂牛乳中差异蛋白进行分析,重点分析了牛乳中主要生物活性蛋白及乳脂球膜蛋白变化,并且阐明了脱脂处理产生的差异蛋白主要涉及的生物学功能及代谢过程,明确脱脂对牛乳全蛋白组的影响。研究结果旨在为消费者购买脱脂牛乳提供知识参考,也可为脱脂牛乳品质优化提供理论支持。
1 材料与方法
1.1 材料与仪器
生牛乳 某大型乳品公司自有牧场;丙酮 国药集团化学试剂有限公司;TMT 标记试剂盒、BCA 试剂盒 美国赛默飞世尔科技公司;二硫苏糖醇(Dithiothreitol,DTT)、碘乙酰胺(Iodacetamide,IAA)、蛋白酶抑制剂 生工生物工程(上海)股份有限公司;胰蛋白酶 北京华利世科技有限公司;三乙基碳酸氢铵缓冲液(Triethyl-ammonium Bicarbonate Buffer,TEAB)美国Sigma 公司;磷酸酶抑制剂 瑞士罗氏公司。
Agilent 1100 高效液相 美国安捷伦科技有限公司;Orbitrap Fusion 质谱仪、EASY-nLC 1000 液相美国赛默飞世尔科技公司。
1.2 实验方法
1.2.1 全脂牛乳和脱脂牛乳制备 将同一批原料乳分成A、B 两组,分别用于全脂牛乳和脱脂牛乳制备。根据工厂液态乳生产工艺,B 组原料乳预热到45~60 ℃,随后在55 ℃下以5000 r/min 离心脱脂,使其脂肪含量降低到0.1 g/100 mL。脱脂后的牛乳和原料乳均采用管式灭菌设备在137 ℃下灭菌4 s,冷却后储存于-80 ℃以备后续实验使用。
1.2.2 蛋白提取与定量 采用冷丙酮沉淀法对牛乳蛋白质进行提取。10 mL 样品中加入磷酸酶抑制剂和蛋白酶抑制剂,使其终浓度为1 mmol/L,然后加入5 倍体积的预冷丙酮,置于-40 ℃过夜沉淀。沉淀后的样品在4 ℃下12000 r/min 离心10 min,收集沉淀,自然风干。将干燥后的样品溶解于SDS 裂解液,并再次于4 ℃下12000 r/min 离心10 min,取上清液。利用BCA 试剂盒测定上清液中蛋白质含量,并用SDS-PAGE 凝胶电泳检测蛋白质提取质量。
1.2.3 蛋白质酶切和TMT 标记 按照测定的浓度,取50 μg 蛋白加入DTT,使其终浓度为1 mmol/L,并在55 ℃下孵育30 min。冷却到室温后加入IAA,使其终浓度为10 mmol/L,避光反应15 min。反应结束后加入6 倍体积丙酮于-40 ℃过夜沉淀蛋白,并于4 ℃下8000×g 离心10 min 收集沉淀。沉淀中加入TEAB(200 mmol/L)复溶,并以蛋白质和酶质量比1:50 加入胰蛋白酶,于37 ℃下过夜酶解。酶解后样品冻干保存在-80 ℃。
利用TMT 标记试剂盒对样品进行标记。干燥后的样品中加入TEAB,根据说明书取适量TMT 试剂加入到样品中,涡旋混匀后反应1 h,并用5%羟胺终止反应。全脂牛乳和脱脂牛乳样品每组做三次重复实验,命名为WM-1、WM-2、WM-3、SM-1、SM-2和SM-3,分别用TMT 试剂盒中编号为131C、132N、132C、133N、133C 和134N 的同位素标签进行标记。
1.2.4 肽段分馏和LC-MS/MS 检测
1.2.4.1 高pH 反相液相色谱分离 参考Zhang 等[22]液相及LC-MS 方法并加以修改,对处理后的样品进行分析。标记后的多肽使用Agilent Zorbax Extend C18(5 μm,2.1×150 mm)色谱柱在Agilent 1100 HPLC 系统上进行分离。流动相A 为2%(v/v)乙腈的水溶液(pH10),流动相B 为10%(v/v)水的乙腈溶液(pH10)。液相洗脱条件为0~8 min,98% A;8~8.01 min,98%~95% A;8.01~48 min,95%~75% A;48~60 min,75%~60% A;60~60.01 min,60%~10%A;60.01~70 min,10% A;70~70.01 min,10%~98%A;70.01~75 min,98% A。在210 nm 下收集8~60 min洗脱下来的样品,平均分装在15 个管中,冷冻干燥备用。
1.2.4.2 LC-MS/MS 检测 采用EASY-nLC 1000液相和Orbitrap Fusion 质谱对标记后的肽段进行检测。溶解后的样品以300 nL/min 的流速上样到预柱Acclaim PepMap100(100 μm×2 cm),再经分析柱Acclaim PepMap RSLC(75 μm×50 cm)分离。流动相A 为0.1%(v/v)甲酸的水溶液,流动相B 为0.1%(v/v)甲酸和19.9%(v/v)水的乙腈溶液。液相分离条件为0~40 min,5%~28% B;40~60 min,28%~42%B;60~65 min,42%~90% B;65~75 min,90% B。一级MS 质量分辨率设为60000,自动增益控制值为1e6,最大注射时间为50 ms。质谱扫描荷质比m/z范围350~1500,所有MS/MS 图谱采集使用数据依赖型的正离子模式下的高能碰撞裂解完成,碰撞能量设为36 eV。MS/MS 的分辨率设为30000,自动增益控制为1e5。
1.3 数据处理
1.3.1 数据采集和蛋白质鉴定 利用Proteome Discover 2.4 软件对原始数据进行分析,背景数据库为Uniprot-Bos Taurus-9913-2022.1018.fasta。具体参数设置如下:胰蛋白酶最大漏切位点为2,MS1 允许偏差10 ppm,MS 允许偏差0.02 Da,FDR≤0.01,且特异肽≥1。依据每组样品平均值的差异倍数(Fold Changes,FC,FC=SM/WM)对差异蛋白进行筛选。FC>1.2 或FC<0.83,且P<0.05(T 检验)的蛋白质被认为是差异蛋白[18]。
1.3.2 生物信息学分析 基于基因本体GO 数据库(http://www.geneontology.org)和京都基因和基因组百科全书KEGG 数据库(http://www.genome.jp/kegg/)对差异蛋白进行功能注释和富集分析,利用超几何分布检验计算代表功能集在差异蛋白列表中是否显著富集的P值。根据P值对结果进行筛选与分析,利用Origin 2021 绘图。
2 结果与分析
2.1 牛乳蛋白质谱鉴定结果
采用TMT 标记定量技术对全脂牛乳和脱脂牛乳蛋白质组变化进行分析,LC-MS/MS 共检测出二级谱图377413 个,其中有效谱图共有18870 个。在FDR 为1%的条件下对谱图进行筛选,全脂牛乳和脱脂牛乳中共鉴定出6902 条肽段,这些肽段来自1724 个牛乳蛋白。经过数据过滤,两种牛乳中共检出1352 个可信蛋白。鉴定出的蛋白质(74%)分子量集中在10~20 kDa(250)、20~30 kDa(283)、30~40 kDa(222)、40~50 kDa(210)、50~60 kDa(190)和60~70 kDa(125)(图1)。鉴定到的大多数肽段氨基酸个数为6~20,检测结果中蛋白质分子量和肽段长度均符合胰蛋白酶消化特性[19],质谱数据可以满足后续的分析要求。
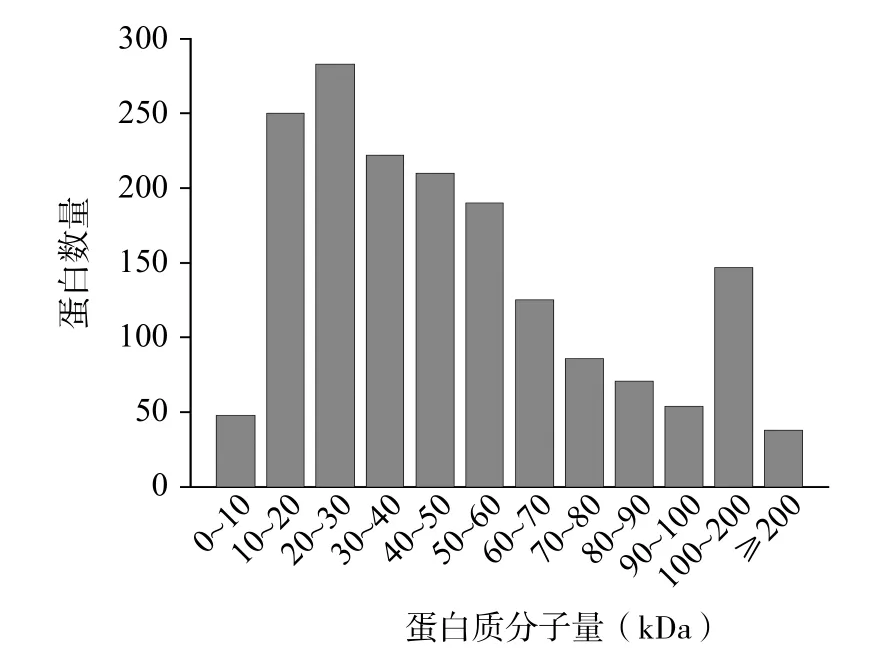
图1 鉴定出的蛋白质分子量分布Fig.1 Molecular weight distribution of the identified proteins
2.2 全脂牛乳和脱脂牛乳差异蛋白筛选
依据差异倍数和T 检验结果在全脂牛乳和脱脂牛乳中共筛选出199 个差异蛋白,火山图(图2)较直观地表示出脱脂前后牛乳蛋白质组变化。与全脂牛乳相比,脱脂后有67 个蛋白上调,132 个蛋白下调,且差异蛋白多集中于对称轴,表明离心脱脂后牛乳差异蛋白变化较小。参考牛乳蛋白研究的相关文献,根据蛋白在牛乳中的含量、文献已检出的乳脂球膜蛋白和与脂质相关性较大的蛋白进行筛选,确定了48 种蛋白进行分析(表1)。鉴定出的可信蛋白中包括酪蛋白、乳清蛋白和乳脂球膜蛋白等牛乳中主要的营养蛋白,差异蛋白在功能分类上包括细胞骨架蛋白、代谢(脂质、氨基酸、核苷酸)相关蛋白、信号转导和伴侣蛋白以及细胞内贩运、分泌和囊泡运输相关蛋白等。

图2 全脂牛乳和脱脂牛乳差异蛋白火山图Fig.2 Volcano plots of differentially abundance proteins(DAPs) in whole milk and skim milk
2.2.1 牛乳中主要活性蛋白 酪蛋白和乳清蛋白是牛乳蛋白的主要组成部分,也是牛乳中主要的生物活性蛋白[2,23]。牛乳中酪蛋白主要包括αs1-酪蛋白、αs2-酪蛋白、β-酪蛋白和κ-酪蛋白4 种亚型,在全脂和脱脂牛乳中均有检出。酪蛋白具有免疫调节、抗炎、降血压等多种生物学活性,是生物活性肽的主要前体蛋白[24-25]。牛乳中αs1-酪蛋白、αs2-酪蛋白和β-酪蛋白脱脂前后相对丰度差异倍数分别为0.95、1.12 和1.08,不属于差异蛋白,其相对含量变化不明显,而κ-酪蛋白(FC=0.82,P<0.05)在脱脂后相对含量降低。牛乳中酪蛋白各种亚型通过磷酸钙及疏水相互作用连接,多以酪蛋白胶束的形式存在,热稳定性较好。但由于各亚型的结构不同,因此在胶束中主要存在的位置也不同,κ-酪蛋白多存在于胶束外层[26],在脱脂处理中可能易受到影响。κ-酪蛋白也是唯一发生糖基化的酪蛋白亚型[27],其水解产物十一肽MAIPPKKNQDK(残基106~116)是食源性抗血栓肽主要来源[24]。乳清蛋白作为牛乳蛋白中的主要成分,占总蛋白的20%,包括α-乳白蛋白、β-乳球蛋白、乳铁蛋白和各种免疫蛋白[5],其中β-乳球蛋白和乳铁蛋白(FC 分别为1.43 和1.51,P<0.05)在脱脂后相对含量上升。β-乳球蛋白是脂质运载蛋白,除具有抗菌抗病毒等常规蛋白功能特性外还能够促进脂溶性维生素A 和维生素E 吸收[28]。乳铁蛋白作为铁结合型糖蛋白,在免疫调节、抗癌和抗病毒中发挥着重要的作用[29]。牛乳中乳清蛋白和酪蛋白存在着相互作用,加热温度及pH 均会影响乳清蛋白与酪蛋白的结合反应,其中β-乳球蛋白更易受环境的影响[30-31],脱脂过程中乳中主要的生物活性蛋白κ-酪蛋白和β-乳球蛋白更易受到影响。
2.2.2 乳脂肪球膜蛋白 乳脂肪球膜蛋白主要为糖基化蛋白,只占总牛乳蛋白的1%~4%,但其在抗菌抗病毒、炎症调节、婴幼儿生长发育及免疫系统构建等方面发挥着重要的作用[17,32]。乳脂肪球是牛乳中脂肪存在的主要形式,在离心脱脂过程中被大量去除。由于离心过程中温度及机械力的作用,部分脂肪球膜被破坏,造成乳脂肪球膜蛋白含量发生改变[33]。全脂和脱脂牛乳中鉴定出的乳脂肪球膜蛋白主要包括黄嘌呤脱氢酶/氧化酶(XDH/XO)、muc1 黏蛋白(MUC1)、脂肪酸结合蛋白(FABP)、嗜乳脂蛋白(BTN)和乳凝集素(LADH)[34],除BTN 和LADH 在脱脂后相对含量显著性下降外,其余鉴定出的脂肪球膜蛋白含量无显著性变化,FC 均在0.83~1.2 之间。XDH/XO 占乳脂肪球膜蛋白的12%~13%,是牛乳中主要的免疫蛋白,可以有效抑制致病菌生长[35]。在热加工过程中,UHT 及喷雾干燥等高温处理均可显著性降低(P<0.05)XDH/XO 活性[36],但脱脂对其含量不具有显著性影响(FC=0.97)(P>0.05)。同样地,作为肿瘤生物学标志物的MUC1[37]和与癌症相关的FABP[38]在脱脂后含量未发生显著性变化,其FC 分别为0.87 和0.95。BTN 和LADH 在脱脂后相对含量显著性下降(FC 分别为0.61 和0.66,P<0.05)。BTN1A1 是一种Ⅰ型跨膜糖蛋白,属于免疫球蛋白超家族中的一员,可以与黄嘌呤脱氢酶以及脂肪分化相关蛋白(ADFP)相互作用,在乳脂分泌过程中十分重要[39]。LADH 在免疫防御中发挥关键作用,具有抗菌和抗病毒功能,在改善人类骨骼肌发育中也具有重要作用[40]。在婴幼儿配方奶粉研发的过程中,可以重点关注脱脂后含量降低的蛋白乳脂肪球膜蛋白,从而更加全面地补充婴幼儿所需营养物质。
2.2.3 其它蛋白
2.2.3.1 代谢相关蛋白 KOG 数据库是真核生物蛋白质组数据库,根据其蛋白功能注释,筛选出25 个与代谢相关差异蛋白,涉及脂质、氨基酸、核苷酸、碳水化合物以及无机离子的运输和代谢。除与核苷酸代谢相关的外核苷三磷酸二磷酸水解酶5 和核苷二磷酸酯激酶B 以及与碳水化合物代谢相关的ATP 依赖型6-磷酸果糖激酶外,其余所有代谢相关蛋白脱脂后相对含量均显著性降低(FC<0.83,P<0.05)。脱脂对脂质运输和代谢影响较大,脱脂处理后溶血磷脂酸酰基转移酶和4-羧基固醇-3-脱氢酶差异倍数分别为0.46 和0.43。溶血磷脂酸酰基转移酶(AGPAT6)属于AGPAT 家族,是某些哺乳动物组织中甘油脂和三酰基甘油生物合成的关键酶,与代谢相关慢性病如糖尿病、肥胖症和心血管疾病密切相关[41-42]。4-羧基固醇-3-脱氢酶也与胆固醇合成相关,且是阿尔茨海默症治疗时的药物靶标[43]。
2.2.3.2 细胞骨架蛋白 细胞骨架包括微丝、中间丝及微管,在细胞运动和细胞形态的维持中发挥着重要的作用[44]。脱脂处理对牛乳中骨架蛋白也有影响。Arp2/3 复合体、Coronin-1A 和前纤维蛋白-1 是肌动蛋白形成及发挥作用的辅助因子[45-47],在心血管疾病和癌症治疗等方面具有重要作用,在全脂牛乳中相对含量更高(FC 分别为0.75,0.75 和0.81,P<0.05)。细丝蛋白A、微管蛋白β-3 链和微管蛋白α-1C 链在脱脂牛乳中相对含量更高(FC 分别为1.20,1.22 和1.32,P<0.05),其中微管蛋白二聚体由α-和β-微管蛋白聚合而成,是微管的亚基。微管作为细胞骨架的重要组成部分之一,在维持细胞结构和介导多个细胞反应(染色体分离和细胞内运输)中起着至关重要的作用[48]。
2.2.3.3 核糖体蛋白 核糖体结构蛋白是核糖体的重要组成部分,在核糖体生物合成和蛋白质合成中发挥着重要作用。对白脚虾蛋白受辐射处理影响的研究中发现,核糖体蛋白在氧化条件下容易被破坏,相比于1 kGy 和5 kGy 辐射处理,9 kGy 辐射会对核糖体蛋白造成更严重的影响[19]。脱脂过程中,核糖体蛋白两种亚基变化趋势相反,与全脂牛乳相比,脱脂处理后40S 核糖体蛋白S3a(FC=0.79,P<0.05)相对含量降低,而60S 核糖体蛋白L27 和L3(FC 分别为1.22 和2.38,P<0.05)相对含量升高。核糖体蛋白亚基含量变化可能与脱脂过程中离心力相关,较轻的蛋白在离心过程中更容易随着脂质去除而流失。除了蛋白质合成功能,60S 核糖体蛋白也可发挥抗菌活性,作为抗菌肽的前体物质[49]。
2.3 差异蛋白GO 功能注释分析
对脱脂前后差异蛋白质组做GO 功能注释分析,根据-log10(P-value)选取生物学过程、细胞组分和分子功能前10 的GO 注释信息做柱形图(图3)。结果表明,脱脂后变化蛋白主要涉及的生物学过程是对细菌的防御反应、先天免疫反应、对革兰氏阳性菌的防御反应和由抗菌肽介导的抗微生物体液免疫反应。防御和免疫反应是牛乳乳清蛋白主要参与的生物学过程,对人乳以及不同反刍动物乳清蛋白研究中发现类似结果[5,50]。差异蛋白发挥作用的细胞组分主要包括细胞外空间、细胞外区域、膜和内质网,在以往研究中发现,细胞外区域是牛乳乳清蛋白常见的细胞成分[50-51]。差异蛋白参与的分子功能主要是GTP结合、GTP 酶活性、肝素结合和GDP 结合。结合功能是牛乳蛋白的主要分子功能,也是牛乳和羊乳主要的差异功能[21,51]。蛋白质不仅与蛋白结合、与小分子物质、维生素等均有结合,脱脂后差异蛋白主要涉及的是与能量代谢相关分子物质结合。
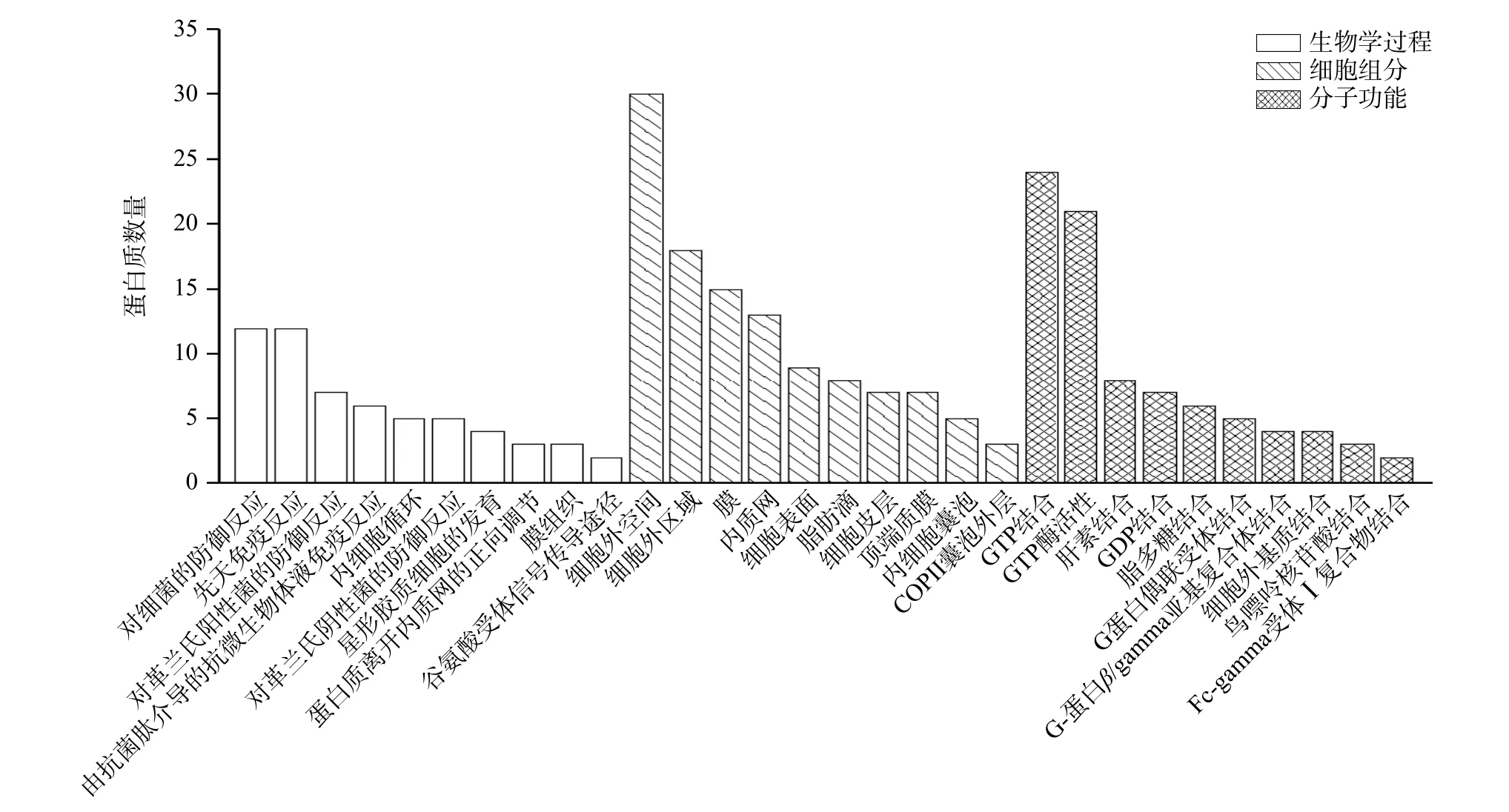
图3 全脂牛乳和脱脂牛乳差异蛋白GO 功能注释分析Fig.3 GO classification of differentially abundance proteins (DAPs) of whole milk and skim milk
2.4 差异蛋白KEGG 代谢通路分析
利用KEGG 数据库对全脂牛乳和脱脂牛乳中差异蛋白代谢通路进行分析(图4),共富集到218 条代谢通路,其中显著性富集(P<0.05)的有41 条。涉及的代谢通路包括5 个类型,其中有机系统和人类疾病相关的通路占主要部分。受脱脂影响的蛋白主要富集在中性粒细胞外陷阱形成、结核病、阿米巴病和金黄色葡萄球菌感染等代谢通路,与疾病调控代谢通路相关。在富集到的通路中唯一与脂质代谢相关的是类固醇合成,包括萜烯环化酶、3-β-羟基类固醇3-脱氢酶和甾醇-4-α-羧酸酯3-脱氢酶3 个蛋白。有研究表明,甾醇-4-α-羧酸酯3-脱氢酶的基因NSDHL是一种新型脂肪调节剂,可以通过降低3T3-L1 细胞中LXR-SREBP1 信号通路而减少脂肪的生成[52]。上述与脂质代谢相关的蛋白在脱脂处理后含量明显下降,可能与牛乳中总脂肪含量下降相关。

图4 全脂牛乳和脱脂牛乳差异蛋白KEGG 代谢通路分析Fig.4 KEGG analysis of differentially abundance proteins (DAPs) of whole milk and skim milk
3 结论
本文首次采用TMT 蛋白标记定量技术对全脂牛乳和脱脂牛乳全蛋白组进行分析,共鉴定出1352个蛋白,包括牛乳乳清蛋白、酪蛋白和乳脂肪球膜蛋白、骨架蛋白和代谢相关蛋白等。由于脂肪含量变化及脱脂过程中离心力和机械力作用,其中132 个蛋白在脱脂后相对含量显著下降,然而部分蛋白如β-乳球蛋白和乳铁蛋白在脱脂后含量升高。乳脂肪球膜蛋白作为牛奶中第三类重要的蛋白质,嗜乳脂蛋白和乳凝集素在脱脂后相对含量显著性降低。这两种蛋白在乳脂分泌以及免疫防御方面发挥着重要的作用,是牛乳中主要的脂球膜蛋白。从蛋白功能角度考虑,差异蛋白涉及细胞骨架、代谢以及信号传导等功能,其中大部分代谢相关蛋白在脱脂后相对含量显著性下降。对全脂牛乳和脱脂牛乳蛋白质组全面分析可以更清楚地了解脱脂对牛乳蛋白产生的影响,为以脱脂牛乳为原料开发功能性食品提供了理论基础,也可为消费者选购不同脂肪含量牛乳制品提供参考。
