脂多糖诱导大鼠胸主动脉内皮源性外泌体的miRNA表达
2023-12-02邓吉鋆
李 莹,邓吉鋆,姜 隽,李 涛
1.西南医科大学附属医院甲状腺外科(泸州 646000);2.西南医科大学心血管医学研究所·西南医科大学-四川省心血管疾病防治协同创新中心(泸州 646000)
脓毒血症的临床发病率和致死率高,发病年龄范围广,是威胁人类健康的重大疾病,一直是急重症医学的重点和难点[1-2]。生理环境下,血管内皮细胞大部分处于静态。当受到病理性刺激时,血清中相关调节因子发生改变,血管内皮细胞活化并参与各种病理生理过程,通过调节血管张力、血小板活性、平滑肌细胞增殖迁移、白细胞黏附聚集及血栓形成等过程,在各种疾病的发生发展中起重要作用。外泌体能将内部的小分子物质运输到邻近或远处细胞,从而实现细胞间的信息通讯。外泌体可能携带上千种miRNAs,但在脓毒血症早期外泌体所分泌的miRNA 方面的研究却不全面[3-4]。
本研究用细菌脂多糖(lipopolysaccharide,LPS)诱导处理血管内皮细胞,模拟脓毒血症早期的病理生理环境。内皮细胞分泌的外泌体含有多种miRNAs,对外泌体进行小RNA 测序,找出差异性表达的miRNA,分析miRNA 对下游靶器官作用的信号通路,为临床应用提供实验依据。
1 实验和方法
1.1 实验动物
SD 雄性大鼠由西南医科大学动物中心提供,8 周龄,SPF 级,30 只,每只质量约250 g。实验动物操作经西南医科大学动物实验伦理委员会审核批准(201903-258)。
1.2 分离和原代培养大鼠胸主动脉内皮细胞
大鼠腹腔注射50 mg/kg 戊巴比妥钠麻醉,仰卧位固定在操作台上。用75%酒精消毒胸前区皮肤,用剪刀依次剪开皮肤和肋骨,暴露出胸主动脉,剪下动脉置于预冷的磷酸盐缓冲溶液(phosphate buffer saline,PBS)中,迅速去除动脉外膜。沿动脉长轴剖开动脉,0.1%的胶原酶I 消化动脉内膜,消化液离心后的沉淀即为内皮细胞。内皮细胞培养基培养5~7 d后即可获得原代的内皮细胞,传4-7代后即可用于后续实验。
内皮细胞生长至70%~80%融合度时,换成无血清的培养基饥饿处理12 h。随机将细胞分为实验组和对照组,更换为完全培养基后,实验组细胞给予0.5µg/mL的细菌脂多糖(Millipore Sigma,美国)诱导处理,对照组给予等体积的PBS,6 h后收集培养基。
1.3 提取外泌体
收集的培养基依次在4℃500×g离心5 min、2 000×g 离心30 min 后,弃沉淀去除细胞碎片、微泡。用15 mL 离心管收集上清液,在上清中加入8%聚乙二醇6 000,4℃冰箱静置8 h。4℃10 000×g 离心60 min,弃上清。用PBS轻柔冲洗管壁,重悬外泌体,4℃120 000×g离心60 min,弃上清,去除残留的聚乙二醇6 000。100µL PBS重悬沉淀即得到外泌体。本研究经过差速超速离心后,成功分离了来源于血管内皮细胞的外泌体,在透射电镜(Hitachi H-9500)下观察到直径为40 nm~100 nm的盘状囊泡[5-6]。
1.4 对外泌体进行测序分析
从内皮细胞的条件培养基中提取的外泌体进行小RNA 测序(康成生物,上海)。外泌体携带miRNA,对miRNA进行差异表达分析。
1.5 统计学方法
采用T-test 分析实验组和对照组测序结果的miRNA 表达差异。筛选出差异表达倍数为>±2 的miRNA,P <0.05 表示差异有统计学意义。miRNA 的排序基于P值的大小,以升序排列。
2 结果
首先体外培养SD大鼠胸主动脉内皮细胞,给予细菌脂多糖诱导处理后,收集的培养基经过差速超速离心后获得外泌体。通过高通量测序检测内皮源性外泌体所携带的小RNA 表达,和静态培养的内皮细胞源性外泌体进行比较,找出差异性表达的miRNA,见图1。
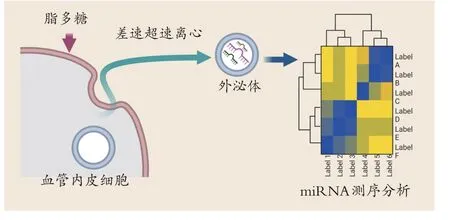
图1 实验流程Figure 1 Experimental procedure
2.1 原代培养大鼠胸主动脉内皮细胞鉴定
免疫荧光显示培养的细胞高表达血管性血友病因子(von Willebrand Factor,vWF)。vWF主要是由血管内皮细胞和巨核细胞产生的一种超大分子量的多聚体糖蛋白,是内皮细胞的生物学标记蛋白。免疫荧光实验的结果明确了培养的细胞为原代大鼠胸主动脉内皮细胞,见图2A。
2.2 提取外泌体的鉴定
细菌脂多糖诱导处理血管内皮细胞6 h后,收集培养基,通过差速离心法分离获得外泌体,在透射电镜下观察到直径为40 nm~ 100 nm 的盘状囊泡,即为细胞分泌的外泌体,见图2B。
2.3 血管内皮细胞分泌的外泌体测序
将两组提取的外泌体进行测序,和对照组的内皮源性外泌体相比,细菌脂多糖刺激下产生的内皮源性外泌体所携带的miRNA 种类和表达水平均有不同。其中13 个表达明显上调,5 个表达明显下调。和正常的内皮源性外泌体相比,细菌脂多糖刺激下产生的内皮源性外泌体所携带的miRNA 种类和表达水平均发生改变。本表列出了差异表达的miRNA的ID、完整序列、种子(seed)序列、差异表达情况、差异倍数和P 值(n=3),见图3。
大多数LPS 刺激下内皮源性外泌体所携带的miRNA 都是对心肌细胞生存和修复有效的,这就可以解释在脓毒血症早期,内皮细胞分泌的外泌体可以增加心肌的活性和减少心肌凋亡,见表1。
3 讨论
脓毒血症整体的死亡率为30%,重症脓毒血症的死亡率可达50%,脓毒血症休克的死亡率则高达80%,一直以来都是重症医学的难点和热点[7]。
内皮细胞是排列在血管、淋巴管和心内膜上的单层扁平细胞,在循环血液、淋巴液和组织之间形成屏障,控制液体、小分子和白细胞的流动。它们具有调控血管张力、血液凝固、吞噬异物、血管新生以及参与炎症反应等功能[8-9]。在脓毒血症中,当病原体进入机体循环后,内皮细胞是最早和病原体及其代谢物接触的宿主细胞。
外泌体是一种产生自真核细胞内体的细胞外囊泡,直径30~150 nm,在生物体血液、尿液、脑脊液、乳汁和唾液中均有发现,在体外培养的细胞也可将外泌体释放到培养基中。它们含有起源细胞的各种分子成分,包括蛋白质和RNA(mRNA 和miRNA)。外泌体的含量和携带的分子随起源细胞变化,从而反应起源细胞的状态。检测外泌体的动态变化可以为疾病的检测提供很有效的窗口[10-11]。
外泌体具有良好的稳定性、靶向性和高效转运等优势,其携带的mRNA成分可以进入靶细胞翻译成蛋白质.外泌体也可以将携带的miRNA 运送到靶细胞,靶向调节细胞中mRNA 及其下游蛋白表达的水平,从而实现细胞之间的信息通讯,进一步参与多种病理生理学过程[12]。外泌体参与细胞间通讯和信号传导,但有关外泌体分泌、摄取、组成以及运输等相关分子机制的研究还不够深入。目前对外泌体的研究兴趣日益增长,无论是研究其功能还是了解对靶器官的作用信号通路俨然成为热点。
miRNA 是一类由19~27 个核糖核苷酸组成的高度保守的内源性非编码的单链RNA,通过抑制mRNA翻译或者促进其降解,从而负向调节靶基因的表达,在发育、细胞增殖、凋亡等过程均发挥作用[13-14]。在心血管领域,miRNA 的相关机制研究也有报道,miRNA 不仅参与胚胎干细胞向心肌细胞的成熟分化,而且在诸多心血管疾病的病理生理过程也发挥重要作用[15-18]。
本研究利用细菌脂多糖诱导处理血管内皮细胞,分离其产生的外泌体并进行miRNA 测序。研究发现,相对于对照组,细菌脂多糖诱导处理血管内皮细胞分泌的外泌体携带的miRNA 表达发生明显改变。其中,miR-221-3p,miR-222-3p,miR-221-5p,miR-155-5p,miR-1247-3p,miR-129-5p,miR-148a-5p,miR-222-5p 的表达明显上调,位于表达变化显著的前8 位。通过生物信息学分析可以推测作用靶基因、预测功能富集以及可能发挥作用的信号通路等。
既往有关于根据miRNA 的表达来判断临床相关疾病发生的研究报道[19-20],但在脓毒血症早期外泌体所分泌的miRNA 的研究却不全面[3-4]。本研究推测内皮细胞分泌外泌体携带的miRNA 作用靶细胞是心肌细胞,显著高表达的多个miRNA 对心肌有抗凋亡的功能。比如miR-221/222 有提高心肌细胞的存活,并且也有助于胚胎干细胞向心肌细胞分化成熟。在缺氧的环境下,miR-221-5p可以抑制心肌细胞凋亡减轻心脏受损[21]。大鼠心梗术后,miR-221-5p 可以增加心肌的存活率和改善心功能[22]。在病毒心肌病的小鼠模型中,miR-148a 可以抑制心肌恶性扩张,提高小鼠生存率[23]。在扩张性心肌病,miR-148a 可以增强小鼠的心功能,提高动物生存率[24]。而miR-155-5p对心脏有正反两面的作用效应[25-26]。
4 结论与启示
本研究结果显示,在脓毒症早期,细菌脂多糖刺激内皮细胞表达特异的miRNA,并推测这些miRNA 作用靶点是心肌细胞。因此我们提出这些差异表达的miRNA可能对脓毒症早期的心血管病理生理学发挥了重要作用。本研究结果为下一步临床防治败血症提供了新的靶点和方向。
