贵阳奥陶系湄潭组海林檎Hemicosmitoid和Glyptocystitoid多样性及古地理分布
2023-11-29李佳乐
李佳乐, 兰 天, 王 约
贵州大学资源与环境工程学院,贵阳 550025
0 引言
海蕾亚门(Blastozoa)由Sprinkle于1973建立,属棘皮动物门,早寒武世—中二叠世均有分布。其中奥陶纪常见的共有5纲[1],分别是始海百合纲、菱孔海林檎纲、双孔海林檎纲、海蕾纲、拟海蕾纲。奥陶纪是棘皮动物演化的重要时期[2],但棘皮动物的碳酸钙质骨骼在死后会迅速分离,所以化石多以破碎的萼板为主,属种多样性记录较低[3-4]。
雕刻海林檎Glyptocystitoida和半海林檎Hemicosmitoida是海蕾亚门菱孔海林檎纲孔菱海林檎目Rhombifera中的两个超科,在北美、欧洲、亚洲的奥陶纪—泥盆纪地层中均有报道[5-7]。前人[8-10]已完成对呈球状或椭球状立体保存海林檎萼部骨骼化石部分属种的分类和鉴别工作。例如:陈重泰等[8]根据海林檎萼部骨骼化石形态特征对滇西保山施甸县奥陶系中的海林檎化石进行分类,分为施甸组中的Sphaerolites、Echinosphaerolites、Sinocystis,蒲缥组中的Ovocystis、Aristocystites、Echinosphaeronites、Heliocrinites、Caryocystites、Paracaryostites、Airstocystis、Pseudoaristocystis等共计11属化石,以及贵州遵义董公寺中—上奥陶统十字铺组中的Sinocystis、Aristocystis、Caryocystis、Sphaeronites等4属化石[9];夏树芳等[10]对陕西省南郑县梁山地区奥陶系西梁组和湖南桃源九溪下奥陶统盘家嘴组的海林檎化石鉴定为Sinocystis和Homocystites两个种;Paul等[4]对贵州三都地区奥陶纪弗洛阶地层中零散的萼部骨骼进行描述,整理出大量的Hemicosmitoid和Glyptocystitoid类海林檎的萼板和茎节化石,并指出这些破碎的海林檎萼板化石为研究早奥陶世棘皮动物提供了新的数据和信息。但Hemicosmitoid和Glyptocystitoid两类化石的属和种一级分类位置仍需要更多的萼部骨片结构信息去进一步完善和证实。Hemicosmitoid类化石在华南产出较少,研究基础薄弱,已发现Hemicosmitoid特征是具有内部呼吸管,排列在一个菱形中形成隐孔菱(cryptorhombs),其中一个半菱形的所有孔都很简单,而另一个半菱形的所有孔都是一簇簇的小孔,称为复合孔[11-12]。本文不仅对萼板表面垂直管形态结构及分布特征进行了补充,还为海林檎的古地理分布增加了新的资料,以期为研究多细胞动物的起源和早期演化提供相应的证据。
1 地质背景
贵州省贵阳乌当地区位处贵阳市北东向约6 km处(图1a),位于扬子板块南部黔南侏罗山式褶皱带的贵阳向斜东翼,归属于黔中隆起和黔南坳陷带过渡带上[13-14]。乌当地区整体处于海相环境,湄潭组时期海水较浅,处于潮下环境,生物碎片破碎程度高[15]。该区奥陶系出露有桐梓组(O1t)、湄潭组下段(O1m1)、湄潭组上段(O2m2)、牯牛潭组(O2g)、黄花冲组(O2-3h)[16](图1b)。湄潭组下段以灰绿--灰紫色页岩及黄灰色砂岩为主,上段以页岩及硅质岩为主。本文所有化石均采自大洼剖面A—A′。大洼剖面岩层倾向130°~145°,倾角为25°~48°,总厚为123.95 m。大洼剖面可分为3个岩性段(图1c):0~86.45 m主要为粉砂质泥岩、页岩、含砂质页岩、砂岩及生物碎屑灰岩;86.45~111.95 m由粉砂岩、页岩以及粉砂质泥岩组成;111.95~123.95 m以粉砂岩和粉砂质泥岩为主[17]。湄潭组化石丰富,包括笔石类、腕足类、三叶虫类、双壳类、软舌螺类、腹足类、海林檎类等化石和遗迹化石[18-20]。
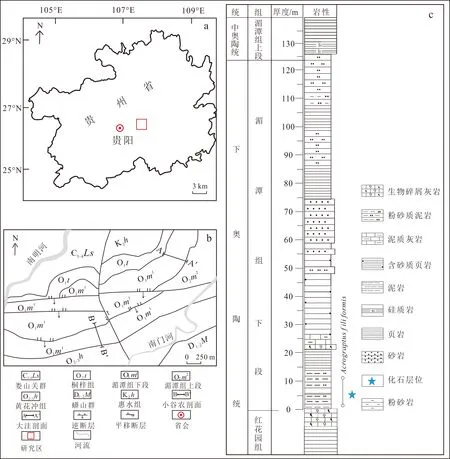
图b、c据文献[16]修改。
2 系统古生物学
乌当地区海林檎化石包括零散的萼板化石和茎节化石。一般来说,在系统古生物学中,萼板化石能为详细分类提供很好的依据。本文对萼板作了详细的分类研究,对茎骨节化石只是进行讨论。所有化石均采自大洼剖面的湄潭组下段,标本存放于贵州大学资源与环境工程学院。
海蕾亚门Subphylum Blastozoa Sprink, 1973
孔菱海林檎目 Order Rhombifera Zittel, 1879
半海林檎超科 Superfamily Hemicosmitoida Bather, 1899
科 Family Caryocrinitidae Bernard, 1895
属 GenusPolycosmitesJaekel, 1918
Polycosmitessp.
特征萼板上具有垂直管(perpendicular canals)和水平管(tangential canals)结构,以及垂直管在萼板上形成的多离散隐孔菱(multidisjunct-cryptorhombs)结构,口部区域未知[21]。
材料3块Polycosmitessp.萼板的内模,具有多离散隐孔菱、垂直管和半隐孔菱(half-cryptorhombs)结构。
描述3块萼板都具有垂直管。其中:标本GRCP2201(图2a)轮廓近似为不规则六边形,下半部分垂直管保留较好,数量较多;垂直管呈菱形分布,形成典型的多离散隐孔菱结构,且能看到水平管的分布(图2a,b)。标本GRCP2202(图2c)为近似六边形轮廓,垂直管集中在萼板中部,呈离散分布;萼板面积相对较小,轮廓呈近似五边形,最宽处为4.7 mm,最长为7.0 mm(图2c,d)。标本GRCP2203具有垂直管和水平管,相比较于标本GRCP2201,标本GRCP2203垂直管数量少,水平管保存较好(图2e,f);垂直管的半菱形分布形成两个半隐孔菱(图3)结构;萼板边缘未发现任何茎节的痕迹,推断其为侧板。

a. 标本GRCP2201,具有多离散的隐孔菱结构六边形骨骼;b. 标本GRCP2201素描图;c. 标本GRCP2202,轮廓为六边形,离散的垂直管分布;d. 标本GRCP2202素描图;e. 标本GRCP2203,两个半隐孔菱结构侧板内模;f. 标本GRCP2203素描图。标本编号前缀GRCP表示标本保存单位为贵州大学古生物研究中心。
讨论Polycosmites最早是依据波希米亚的一块五边形侧板所建立,其特征为多个离散的垂直管贯穿萼板内表面和外表面[6]。Polycosmites中萼管的复杂分布与多离散栉孔菱中孔菱的分布有很大的相似性,所以将这些萼管所组成的菱形称为多离散隐孔菱[22]。贵阳乌当地区所发现的Polycosmites萼板细节结构特征较为全面,与Paul等[4]在贵州省三都地区同高组中发现的Polycosmites骨板相似度最高。从产地来看,两者产地均为贵州下奥陶统弗洛阶时期的页岩和泥岩,在化石采集层不仅采集到了零散的海林檎萼板标本,还发现了海林檎茎节、藻类、三叶虫、腕足等生物化石。从萼板保存结构来看,两地采集到的萼板标本均为内模,未发现外模存在;虽然同高组中采集到的Polycosmites萼板只有1块骨板,但其萼板细节特征保存良好,水平管、垂直管以及垂直管所形成的隐孔菱都可以清楚地观察到;虽然萼板数量较多,但乌当地区未在同一块萼板上观察到清晰的水平管和垂直管同时存在的现象,在不同的萼板上也都发现了这3种结构的存在(图2)。从轮廓来看,贵州三都地区的Polycosmites萼板为1块五边形萼板,乌当地区的萼板五边形和六边形都有。从萼板大小来看,贵州省三都弗洛阶Polycosmites萼板最长为13.0 mm,最宽处为10.0 mm[4],泰国通帕蓬地区最大的Polycosmites萼板最长为9.5 mm,最宽为8.3 mm[22],贵阳乌当地区Polycosmites的萼板最长为10.0 mm,最宽为8.0 mm。由于三都地区Polycosmites萼板厚度的具体数据并没有记录下来,所以无法进行厚度上的对比。总的来说,贵阳乌当地区Polycosmites的发现增加了其在区域上的形态多样性,体现了该属对乌当地区环境的高度适应。
未识别的Hemicosmitoid类萼板化石
材料5块单独的萼板内膜,具有一定特征的萼孔结构。
描述这些萼板在轮廓上有五边形、六边形或者圆形,所有的萼板都有垂直管(图4)。不同萼板垂直管分布不同,部分萼板上垂直管清晰地排列在具有半菱形特征的三角形轮廓上(图4b左上方),部分萼板垂直管呈环形分布(图4d),部分萼板垂直管随机分布在萼板上(图4a,c)。

a. 标本GRCP2204,圆形轮廓的萼板,垂直管直径大小不一;b. 标本GRCP2205,轮廓为五边形的萼板,左上方显示半菱形结构;c. 标本GRCP2206,六边形的萼板,垂直管被填充;d. 标本GRCP2207,七边形轮廓萼板,垂直管直径由中心向四周逐渐增大;e. 标本GRCP2208,垂直管连接到一起的萼板;f. 标本GRCP2209,一个单独的Macrocystella萼板骨片,显示萼板壁外翻形成的脊;g. 标本GRCP2210,具较大径向棘刺的圆形轮廓茎节;h. 标本GRCP2211,具较大径向棘刺的圆形轮廓茎节。
另外,部分萼板具有垂直管孔隙直径较大、数量较多以及少部分垂直管有聚合在一起的现象(图4d,e);垂直管直径范围为0.1~0.5 mm,数量为10~40。
讨论半海林檎超科 (Hemicosmitida) 和雕刻海林檎超科(Glyptocystitoida)形态上的区别是后者具有栉孔菱(pectinirhomb)结构[3-4,6]。图4b—f中萼板表面上都具有垂直管孔隙结构,所以这些萼板应属于Hemicosmitoid类化石。贵阳乌当弗洛阶湄潭组下段Hemicosmitoid类萼板化石与贵州三都地区弗洛阶同高组的Hemicosmitoid类萼板化石相似度较高,但与典型的Hemicosmitoid类萼板化石存在差异。从垂直管分布来看:贵阳乌当的萼板化石最大的特点是垂直管的环形分布,典型的Hemicosmitoid类萼板化石垂直管为菱形或半菱形分布并在萼板上形成隐孔菱结构[11];从轮廓上来看,典型的Hemicosmitoid类萼板化石轮廓是五边形或六边形,而乌当地区萼板轮廓很不规则,有五边形(图4b)、七边形(图4d)和相对较圆的轮廓。将其与Hemicosmitoid类萼板化石中典型的3个科一级Hemicosmitidae、Caryocrinitidae和Thomacystidae萼板的图版[23]进行详细对比可发现:3个科中萼部具有隐孔菱结构的侧板,下侧板及底板的轮廓从五边形到七边形都有,且底板的底部通常会保存部分茎的结构[24];乌当和三都地区的Hemicosmitoid类萼板化石并未发现七边形萼板以及带有部分茎节的底板的存在。由此看来,乌当地区的Hemicosmitoid类萼板化石可能是部分保存;另外,乌当和三都地区萼板上垂直管都有聚合在一起的现象(图4e)且垂直管孔隙直径差异较大,三都地区萼板的垂直管孔隙直径最大能达到0.5 mm,最小为0.3 mm[4]。
总的来说,如果要对这些Hemicosmitoid类萼板化石进行更精确的分类,还需要更多的形态特征和信息。
孔菱海林檎目 Order Rhombifera Zittel, 1879
雕刻海林檎超科 Superfamily Glyptocystitoida Bather, 1899
科 Family Macrocystellidae Bather,1899
属 GenusMacrocystellaPaul,1968
Macrocystellasp.
材料单个Macrocystella属萼板骨片,显示骨板中的褶皱形成的主脊和辅助脊。
描述Macrocystella属萼板骨片,近圆形,显示外脊。骨板中间具有隆起,脊从隆起处辐射到侧面的中间,连接相邻萼板的中心。辅助脊平行于主脊形成类似菱形结构,这些类似菱形结构是由骨板中的褶皱形成的(图5)。这些外脊很薄,厚度大约为0.1 mm。

据文献[6]修改。
讨论目前世界各地所发现的Macrocystella多为零散萼板和茎节,对这些零散的标本化石进行分类时,其中最为关键的2个条件是近端茎节的形状、萼板上主脊和副脊的数量及分布特点[25-26]。1968年Paul等[6]认为脊的存在利于萼板的生长,其原理为萼板褶皱所形成的脊增加了气体交换面的面积,而气体交换量与表面积成正比,所以交换面积的增加表示交换量的增加,这进一步使氧的交换量和需求量相等或者交换量更大,从而利于萼板的生长。将乌当地区Macrocystellasp.萼板与其他属种对比:Macrocystellamariae萼板轮廓为圆形,主脊的横截面形状与乌当地区Macrocystellasp.近似,为圆柱形,不同点在于Macrocystellamariae主脊较高,利于与副脊区分[27];Macrocystellabohemicus通常有2个副脊,近端茎与M.mariae相似,但外部近端茎有少量近似刀状的凸缘存在[6];M.azaisi副脊的数量较多,一般为5~8个,近端茎轮廓近似为圆形,较为特殊的是其凸缘周围有细小的不规则颗粒或棘刺[28];M.bavarica的近端茎轮廓为多边形,副脊的数量较多,最多可以达到7对。综上所述,乌当地区Macrocystellasp.萼板与Macrocystellamariae萼板较为相似。
3 茎节化石
在棘皮动物离散的骨片化石中,茎节化石占有重要的地位,然而现有的标本采集和研究大部分都集中在萼板上,忽略了茎节化石的重要性[4]。
图4g、h为2块单独的茎节化石,其中腔(lumen)位于中心,也为圆形;从中腔至最外层有2个圆形轮廓,较小圆形直径为1.0 mm(在中心处),较大圆形直径为2.0 mm(在外围);边缘仅有1个较大的径向棘刺(radial spines),长度为1.5~2.0 mm,最宽处为1.0 mm。与Hemicosmitoid的茎节对比来看,乌当地区径向棘刺茎节直径较小,约为1.8 mm,而Hemicosmitoid的茎节直径较大,约为2.3 mm;乌当地区径向棘刺较大且只有1个,Hemicosmitoid的茎节具有多个较小径向棘刺且有网眼状空隙[4],但两者中腔形状相同。
2014年,Paul等[4]在对径向棘刺研究中发现径向棘刺可能存在两种作用,分别为应对被捕食和帮助其在水中浮游。笔者在乌当地区发现了较为完整的茎部化石,茎的两侧也长有棘刺,棘刺的长度和位置各不相同,但未找到萼部,所以我们推测茎节上这些大的棘刺可能为其茎部化石横截面。另外,笔者在乌当地区也发现了一些不带有径向棘的、轮廓呈圆形的茎节,其中腔较大,形状与具有径向棘的不同,暂未确定其亲属关系。
图6a为疑似棘皮动物化石,无茎结构,萼部长约为5.0 mm,宽为3.0 mm;萼部上方共有3个腕枝,长度均为2.0~4.0 mm,其中2个腕枝共有4个分枝长出,其中2个分枝长度相近,约为2.0 mm,另外2个分枝长度相差较大,分别为2.0 和0.3 mm。图6b为Glyptocystitoid类海林檎标本,萼部损坏严重,长为6.0 mm,宽为4.0 mm;茎保存较为完整,总长约为13.0 mm,分为近端(proximal stem)和远端(distal stem)两部分,其中近端茎长约为5.0 mm,远端茎长约为8.0 mm。从图6b可以看出近端茎到远端茎转变的过程,其特点是茎的直径越来越小,内茎(inner columnals)的高度逐渐增加,外茎的凸缘(flange)逐渐减少。
4 古地理分布
图7总结了奥陶纪雕刻海林檎Glyptocystitoida和半海林檎Hemicosmitoida部分属种地层分布情况以及Macrocystella和Polycosmites新的层位记录。根据围肛部(periproct)周围萼板的数目将两超科属种分成4类,分别为3或4块、5块、6或8块、2块。具体来说:Echinoencrinitidae和Callocystitidae 两科中围肛部为3或4块萼板;Cheirocrinidae科中除Sprinkleocystis外所有属种都为5块萼板;Pleurocystitidae科中大部分属种为6或8块萼板;最为特殊的是,Rhomifera围肛部非常小,只有2块萼板。
近年来,前人对寒武纪—奥陶纪大辐射时期的古生物地理和生物多样性模式进行了广泛的研究。然而,大多数研究集中在海洋无脊椎动物的3个类群(腕足类动物、软体动物和三叶虫),棘皮动物却被忽视了。在奥陶纪,棘皮动物经历了重要的多样性变化,并与腕足动物、软体动物和三叶虫一起,成为海洋底栖生物的主要组成部分[7,21]。前人[1,4-5]已统计出奥陶纪棘皮动物分布大致分为6个区域,分别是劳伦大陆(Laurentian Province)、波罗地大陆(Baltic Province)、冈瓦纳大陆西部(Western Gondwanan Province)、冈瓦纳大陆东部(Eastern Gondwanan Province)、阿瓦隆地区(Avalonia)和西伯利亚大陆(Siberian Province)。本文所统计的半海林檎Hemicosmitoida和雕刻海林檎Glyptocystitoida属种地理分布(图8)基本与前人[1,4-5]一致,只有西伯利亚大陆未发现两超科属种,未统计在内。经统计,在奥陶纪时期半海林檎Hemicosmitoida分类为3科6属,雕刻海林檎Glyptocystitoida分类为9科28属。雕刻海林檎Glyptocystitoida的属种数量较多,如图8a所示,古地理分布大概为5个区域,分别是劳伦大陆、波罗地大陆、阿瓦隆地区、冈瓦那大陆西部和冈瓦纳大陆东部。其中,冈瓦纳大陆东部分布的属种为Echinoencrinitessenckenbergii[7]、Macrocystella[23]、Scoliocystisthersites和Polycosmites,阿瓦隆地区为Homocystitesanatiformis和Homocystitesalter,波罗地大陆为Erinocystis、Gonocrinites、Glaphyrocystis、Fusicystis、Cheirocystisardmorensis、Cheirocystisradiatus。

圈中数字代表图7中海林檎属种。据文献[30]修改。
半海林檎Hemicosmitoida的属一级古地理分布大概为4个区域,分别是波罗地大陆、阿瓦隆地区、冈瓦纳大陆西部和冈瓦纳大陆东部(图8b)。其中,波罗地大陆分布的属种为Juglandocrinus,阿瓦隆地区为Caryocrinitesornatus和Caryocrinitesrugatus,冈瓦纳大陆西部为Thomacystis,冈瓦纳大陆东部为Paracaryocrinites、Polycosmites。
另外,两者属种古地理分布可能受纬度和温度的影响。例如,早奥陶世气温较高,雕刻海林檎Glyptocystitoida的属种分布相对分散,5个区域均有分布;中奥陶世相对温暖,属种分布区域变为3个低纬度地区;晚奥陶世属种分布区域全部集中在赤道附近,且从桑比期到赫南特期属种数量逐渐减少。这种分布趋势与Paul等[6]所统计的海蕾亚门动物类群纲一级的地理分布类似。从阿瓦隆地区和伊比利亚半岛分布着相同的属种Caryocrinitessp.可以看出,阿瓦隆地区在奥陶纪中晚期从冈瓦纳周围边缘迁移到波罗地大陆的过程[25]。属种的分布表明,在地理上遥远但位于相似纬度的地区,如波罗地大陆和冈瓦纳东部地区之间存在物种交流,比如Hemicosmitessp.。
5 结论
1)在贵阳乌当奥陶系弗洛阶湄潭组下段页岩和砂岩中,采集到大量的棘皮动物化石, 共计 2 属 2种(1 相似种和1未定种),包括:Caryocrinitidae,1 属 1 种(1 未定种),即Polycosmitessp.半海林檎类(Hemicosmitoid)萼板化石;Macrocystellidae,1 属 1 种, 即Macrocystellamariae萼板化石。
2)通过对全球Polycosmites、Macrocystella两个属的古地理分布情况统计可知:中国南部贵州湄潭组和同高组发现的Polycosmites将该属的地层分布从中奥陶统达瑞威尔阶扩展到下奥陶统弗洛阶;贵阳乌当湄潭组Macrocystellamariae的发现将其分布区域扩大为2个,分别为阿瓦隆地区和冈瓦纳大陆东部地区。
