哈尔滨兆麟公园繁殖期鸳鸯日行为模式比较研究
2023-11-25NGOTHIKIEUTRANG杨雨尘田一柳王佳明吕泓学邓文攸吴庆明
NGO THI KIEU TRANG,杨雨尘,李 浙,田一柳,王佳明,吕泓学,邓文攸,石 娟,吴庆明
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.黑龙江凉水国家级自然保护区管理局,伊春,153106)
行为节律是指在特定环境条件下,动物个体表现出来的行为适应性方面的活动规律[1-2],而时间分配是动物针对特定环境条件所表现出来的不同行为类型的时间组成[3-4]。鸟类行为的活动节律与时间分配直接关系到鸟类个体的生长、发育及种群的繁衍。随着环境条件的变化,活动节律与时间分配亦会相应改变[5-6]。探究鸟类繁殖期各类行为的时间分配和节律,对了解鸟类繁殖期行为模式和行为适应策略具有重要意义,对分析其他生活史阶段相对应的行为具有重要的参考价值[7-9]。
鸳鸯(Aix galericulata)隶属于雁形目(Anseriformes)鸭科(Anatidae),是国家二级重点保护野生鸟类。全球多个国家均有分布,如中国、俄罗斯、日本和韩国等[10]。中国东北地区是鸳鸯主要繁殖区。哈尔滨兆麟公园(以下简称“兆麟公园”)自1995 年以来,每年4—7 月鸳鸯均来此繁殖,不论是秋季南迁时的群体数量,还是夏季繁殖的窝数及雏鸟数量,均显著上升,目前已成为兆麟公园吸引游客的特殊生态景观。关于鸳鸯的生态研究较多,主要集中在越冬群体数量、空间分布和性比[11]、越冬日行为活动时间分配[12-13]、越冬栖息地特征[14]、繁殖生态[15-17]、繁殖期食性[18-19]和卵的巢寄生[20-21]等方面,但繁殖期城市公园鸳鸯日行为模式的研究尚未见报道。本研究以兆麟公园内自由生活的野生鸳鸯为研究对象,对鸳鸯的日行为模式进行研究,试图了解城市公园生境条件下不同性别、不同阶段及成幼间鸳鸯日行为时间分配及行为节律,探讨鸳鸯在城市公园的行为适应及繁殖对策,为公园公众赏游条件下鸳鸯的生态管理及其种群的健康发展提供基础数据和科学依据。
1 研究区概况
兆麟公园位于黑龙江省哈尔滨市道里区中心地带(45°46'31.56″—45°46'43.51″ N,126°37'15.49″—126°37'28.86″ E),邻近松花江畔,始建于1906 年,占地面积8.4 hm2,是集休闲、文化和游憩为一体的综合型非营利性公园,游客流量一直较大。该区域属于典型的中温带大陆性季风气候,四季分明,夏季凉爽短暂,7月平均气温为23 ℃,降水集中在6—9月[22]。公园植被相对丰富,乔木包括榆(Ulmus pumila)、红皮云杉(Picea koraiensis)和樟子松(Pinus sylvestrisvar.mongolica)等,共计38种1 000余株;灌木包括丁香(Syringaspp.)、榆叶梅(Prunus triloba)、连翘(Forsythia suspensa)、玫瑰(Rosa rugosa)等,共24 种2 000余株[23]。
公园近中心区域建有“鸳鸯岛”,岛外围为环形人工湖,湖边围有游园路,面积约为1.74 hm2。岛上有粗大的乔木,可作为鸳鸯巢树,还有假山,可阻隔湖边的游人干扰,这些能为鸳鸯安全繁殖提供稳定的平台和隐蔽环境。环形人工湖及湖边区域能为鸳鸯觅食、休息提供持续的、安全的场所。每年4 月初,野生鸳鸯陆续迁来,部分个体留在鸳鸯岛繁殖,部分个体在岛边湖休息。2021 年,兆麟公园有20 余巢鸳鸯 繁殖(https://www.sohu.com/a/469047017_349336),秋季迁徙集群达到500 余只(http://hlj.ifeng.com/c/8AAYOV4Lykj)。因鸳鸯形成稳定的特定生态景观使得兆麟公园成为哈尔滨市公众和媒体关注的景点。每年4—9 月,兆麟公园管理部门采用定时定点人工投饲及专人看护等方式,为鸳鸯的繁殖、休息和觅食等提供持续的正向安稳环境。
2 研究方法
行为观察为2021 年4—7 月,为鸳鸯卵期和雏期。行为观察始于2021 年4 月13 日,4 月13—22 日为行为预观察阶段,主要搜集当年和历年公园内鸳鸯的迁来信息、群体数量、性别组成、栖息位置、洞巢位点和繁殖巢数等,进而确定行为观察点、行为取样时间段、行为取样时间间隔和行为取样样本数量等。同时,结合以往研究成果[13,24]对鸳鸯的行为谱通过行为预观察进行完善,确定本研究记录的8 种行为类型(表1)。
观测点选择有鸳鸯集群活动、数量多并利于观察的湖岸边位点。根据研究区日出日落时间、可视度、研究对象活动规律及园内游客的活动时间,确定日行为观察时间为06:00—17:00。卵期观察设定在4 月23 日—5 月26 日,选取鸳鸯样本20 只(10♀,10);雏期观察设定在6 月9 日—7 月10 日,选取鸳鸯样本20只(10只成体,10只幼体)。采用瞬时扫描法,利用双筒望远镜(8×42)和单反相机,每5 min对视野内的研究对象进行1 次行为观察取样,行为观察采用1 人固定观察、1人固定记录的方式以减少数据收集的误差。对于不确定的行为,用单反相机拍照、录像进行最后确定。整个观察期间,观察区域内鸳鸯最大的群体数量为86 只,能保证行为观察的样本量。理论上,鸳鸯行为样本量应为175 560 个,筛选掉因公园施工或游客等因素所致的噪点行为数据,有效观察样本量为173 246个,有效率为98.68%,累积观察726 h。
行为数据以平均值±标准误表示。采用卡方检验对鸳鸯日行为时间分配的性别差异、阶段差异和成幼差异进行分析。数据统计分析与图表制作均在Excel和SPSS 25.0中完成。
3 结果
3.1 鸳鸯日行为频次分配差异
3.1.1 卵期性别差异
卵期,不同性别鸳鸯的优势行为相似,均以静栖占绝对优势,雄性(40.63%±2.91%),雌性(39.54%±2.28%),其次是站立、游走、理羽和觅食(图1)。但日行为频次分配存在显著不同,鸳鸯用于站立(χ2=24.431,p<0.05)、觅食(χ2=56.773,p<0.05)和打斗(χ2=32.071,p<0.05)行为频次比例在性别上具有显著差异,雄性站立和打斗行为频次显著高于雌性,觅食行为频次显著低于雌性;而二者游走、静栖、理羽、警戒和飞行行为频次差异不明显。
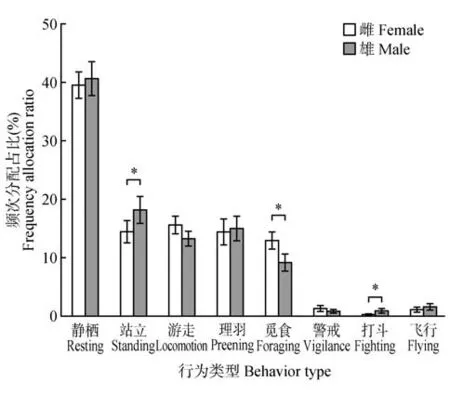
图1 鸳鸯不同性别日行为频次分配差异Fig.1 Gender difference in time allocation of mandarin ducks’ daily behaviors
3.1.2 卵期与雏期差异
卵期和雏期,鸳鸯日行为频次分配的优势行为相似,均以静栖占绝对优势,卵期(40.49%±1.98%),雏期(39.23%±3.77%);其次是站立、游走、理羽和觅食(图2)。日行为频次分配方面存在显著不同,鸳鸯用于站立(χ2=277.192,p<0.05)、游 走(χ2=88.355,p<0.05)、理羽(χ2=277.192,p<0.05)和觅食(χ2=415.416,p<0.05)行为频次比例在卵期和雏期具有显著差异,卵期游走和觅食行为频次显著高于雏期,站立和理羽行为频次显著低于雏期;而卵期和雏期中静栖、警戒、打斗和飞行行为频次分配差异不明显。
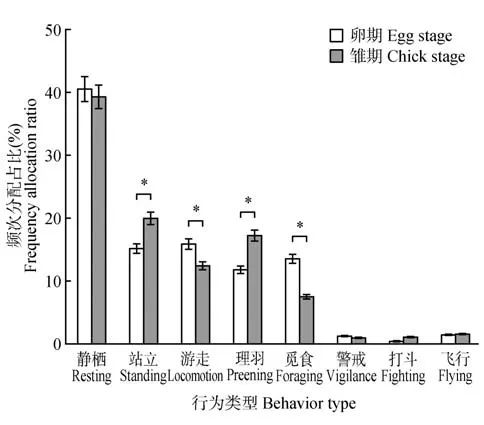
图2 鸳鸯卵期与雏期日行为频次分配差异Fig.2 Comparison of time allocation of mandarin ducks’ daily behaviors between egg stage and chick stage
3.1.3 成幼差异
鸳鸯成体、幼体的优势行为存在显著差异,静栖(41.09%±2.60%)为鸳鸯成体的主要行为,其次为站立、理羽、游走和觅食;而幼体的优势行为是游走(45.55%±4.91%)和觅食(20.13%±6.50%),其次是静栖、站立和理羽(图3)。卡方检验表明,鸳鸯用于静栖(χ2=551.371,p<0.05)、游走(χ2=2 217.747,p<0.05)、觅食(χ2=131.249,p<0.05)、站立(χ2=340.475,p<0.05)和理羽(χ2=374.850,p<0.05)行为的频次比例在成幼间具有显著差异,成体用于静栖、站立和理羽行为频次占比显著高于幼体,用于游走和觅食行为频次占比显著低于幼体;而警戒、打斗和飞行行为频次分配差异不明显。
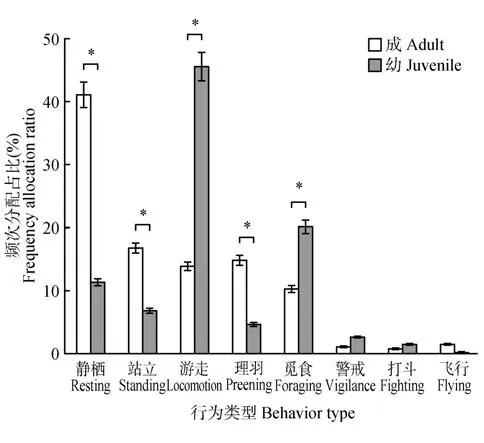
图3 鸳鸯成鸟和幼鸟日行为频次分配差异Fig.3 Comparison of time allocation of adult and juvenile mandarin ducks’ daily behavior
3.2 鸳鸯日行为节律差异
3.2.1 卵期性别差异
卵期,鸳鸯行为频次节律存在性别差异(图4)。其中,静栖行为,不同性别鸳鸯的节律一致,均呈非对称的正态分布,仅有1 个峰值(09:00—10:00),但雌性静栖行为频次在15:00—16:00 出现明显拐点;站立行为,不同性别的节律均呈左低右高的U 形,有2个峰值,均为07:00—08:00和16:00—17:00;游走行为,二者均呈波浪状分布,但雌性的游走行为频次与雄性相比,波动幅度较小,谷值不明显;理羽行为,不同性别在07:00—08:00 和13:00—14:00 存在明显峰值,其余时间段的行为频次分配呈螺旋式互补;觅食行为,在不同性别上的总体趋势基本一致,呈不显著的W 型,有3 个峰值(06:00—09:00、11:00—12:00、16:00—17:00),2 个谷值(09:00—10:00、12:00—15:00);警戒行为,不同性别鸳鸯均呈不规则的W 型,均有3 个峰值(06:00—07:00、10:00—11:00、16:00—17:00),2 个谷值(07:00—08:00、12:00—13:00);打斗行为,不同性别呈上下互补的双螺旋结构,二者在07:00—08:00有1个明显峰值,而雌性在12:00—14:00 和15:00—17:00 未出现打斗行为;飞行行为在不同性别上的总体趋势基本一致,呈左高右低的非对称L 形,有明显的2 个峰值(06:00—07:00、14:00—16:00)。
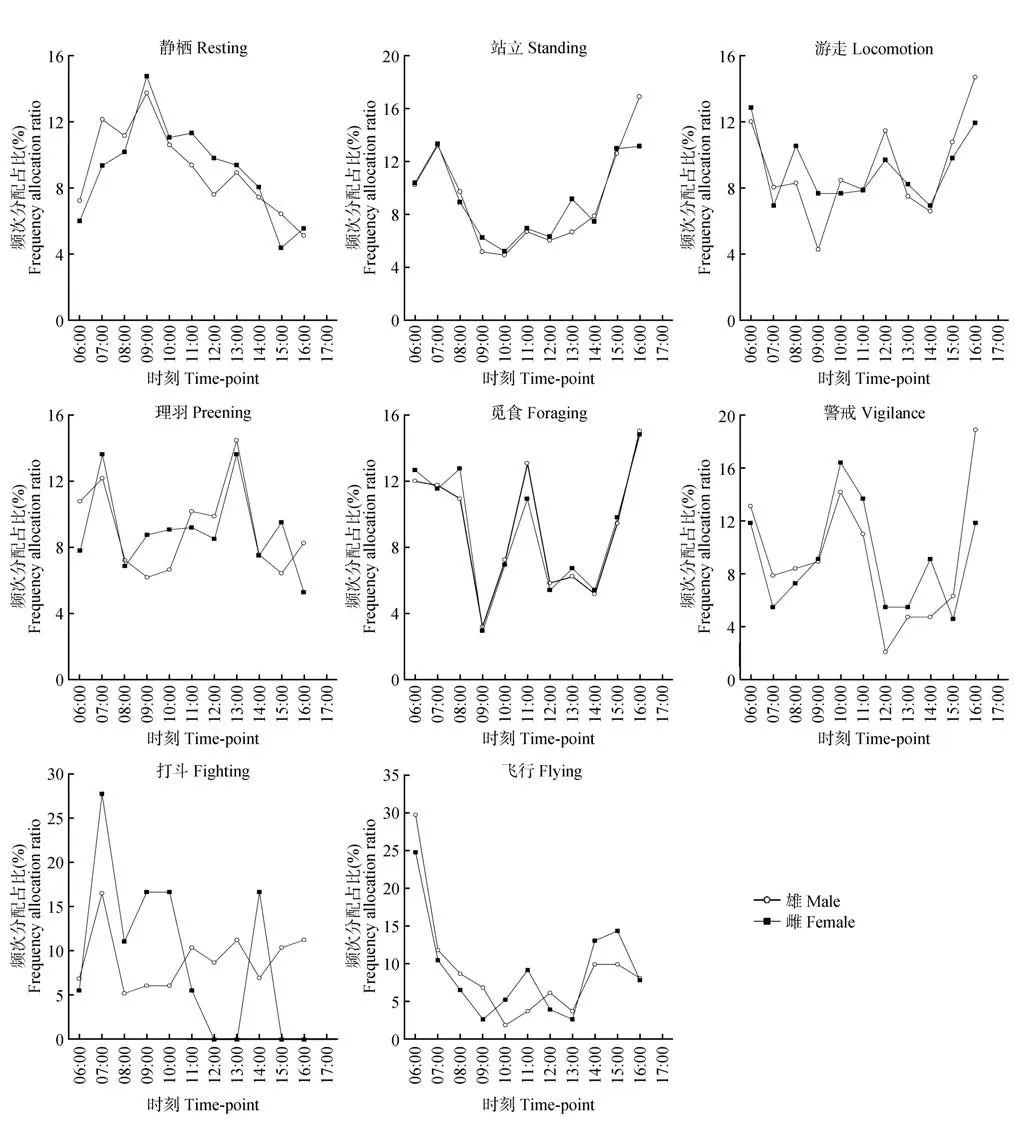
图4 鸳鸯不同性别日行为节律Fig.4 Daily behavioral rhythms of female and male mandarin ducks
3.2.2 卵期与雏期差异
鸳鸯在卵期和雏期,静栖(非对称的正态分布,仅1 个峰值,09:00—10:00)、站立(U 形分布,2 个峰值,07:00—08:00 和16:00—17:00)、游走(W 形分布,有3 个 峰值,06:00—07:00、12:00—13:00、16:00—17:00,2 个谷值,09:00—10:00、14:00—15:00)、理羽(左低右高的2 个峰值,07:00—08:00和13:00—14:00)和觅食(W 形分布,第1 个峰值,卵期06:00—08:00和雏期08:00—09:00有区别,后2 个峰值相同,均为11:00—12:00 和16:00—17:00,2个谷值均为09:00—10:00和12:00—15:00)行为频次节律总体趋势一致;警戒行为,2 个时期的节律频率整体趋于一致,有3个峰值,分别为06:00—07:00、10:00—11:00 和16:00—17:00,其余时段二者呈不规则的互补分布;在打斗行为频次上,相较于雏期,卵期的波动较大,2个时期完全呈上下波动的螺旋结构;飞行行为,卵期和雏期在06:00—07:00 均为最高峰,卵期在15:00—16:00 达到次峰值,雏期则不明显(图5)。
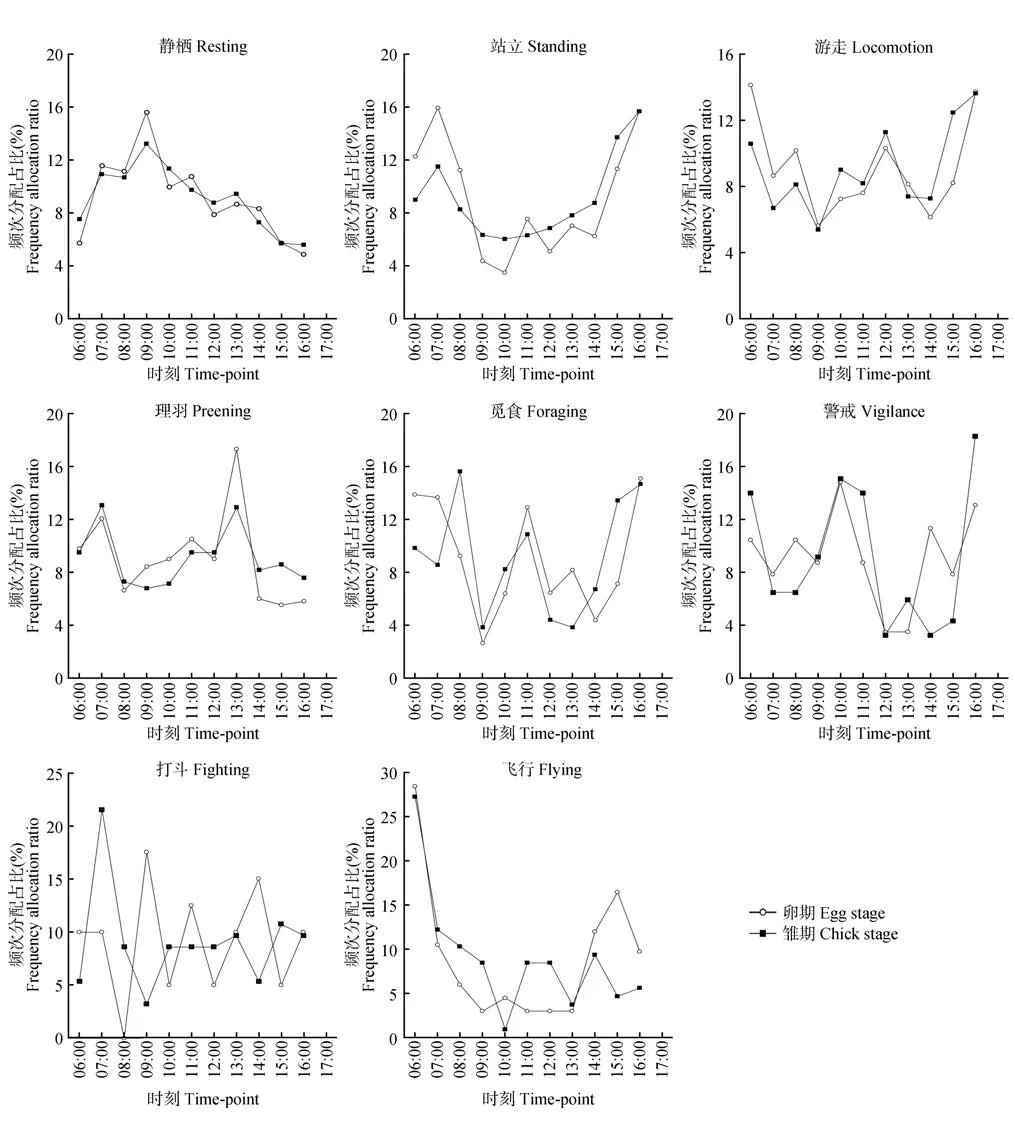
图5 鸳鸯卵期与雏期日行为节律Fig.5 Daily behavioral rhythms of mandarin ducks between egg and chick stages
3.2.3 成幼差异
鸳鸯成体与幼体的行为节律存在显著差异。静栖行为,幼体存在明显峰值,为12:00—14:00,而成体上午的活动节律高于下午,但没有明显峰值与谷值;站立行为,幼体存在明显峰值,为09:00—12:00,而成体的站立在全时段均有发生,但峰值与谷值不明显;游走行为,成体和幼体全时段均有发生,但时间分配呈互补的螺旋结构;理羽行为,幼体13:00—17:00发生频次为全时段最高,其余时段发生频次较低或不发生(10:00—13:00),而成体全时段均理羽,但不存在明显的峰值和谷值;觅食行为,成体与幼体全时段均有发生,但峰值存在差异,11:00—12:00和14:00—15:00 为幼体的2 个觅食峰值,而成体的峰值为11:00—12:00 和16:00—17:00;警戒行为,幼体存在明显的峰值,为11:00—14:00,其余时段警戒频次较低或没有警戒,而成体全时段均有警戒,但没有峰值和谷值且频次分配较低;打斗、飞行行为的节律与警戒基本一致,只是幼体的峰值存在差异,分别为10:00—11:00和12:00—13:00(图6)。
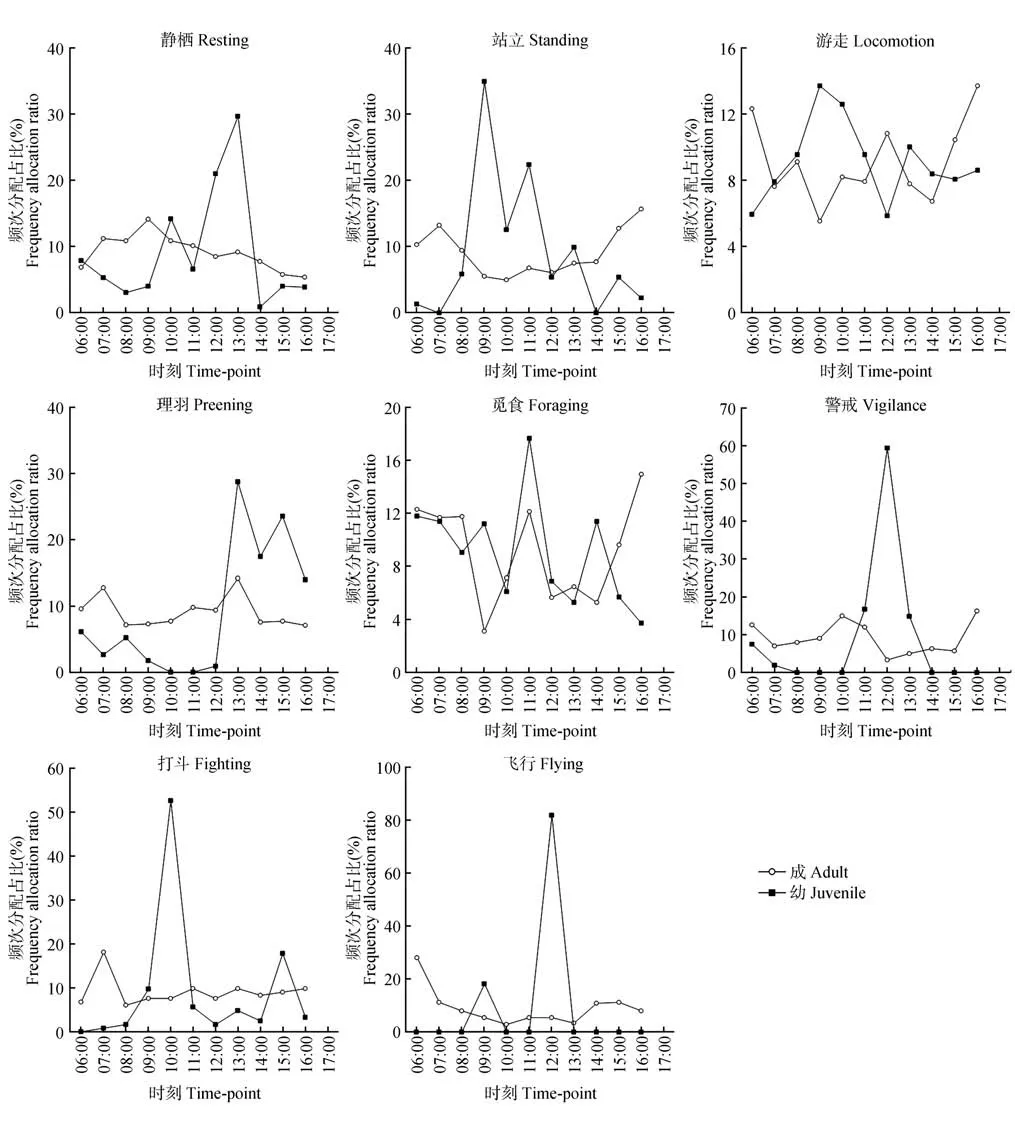
图6 鸳鸯成幼日行为节律Fig.6 Daily behavioral rhythm of adult and juvenile mandarin ducks
4 讨论
鸟类的生存状态通过行为模式表达,并受能量消耗影响[25]。鸟类的行为频次分配不仅是对环境条件的一种适应,更是其全部影响因素作用下的综合表现[5],要理解鸟类个体对时间的最佳利用,就必须清楚鸟类对各种行为类型的频次分配。
从鸳鸯8 种主要行为频次分配的特点可以看出,静栖、游走和站立行为占70%以上,其他行为所占频次比例不足30%。这意味着兆麟公园内在人类活动高强度正向干预下,鸳鸯卵期和雏期整体上是以休息状态为主。鸟类在春季迁飞前会积蓄能量,体质量明显增加,为远距离迁徙做准备[8],当到达繁殖地后,与觅食、积蓄能量相关的行为已经不再是主要行为,主要行为时间切换到繁殖后代的系列活动中,本研究从侧面印证了这一结论。
从不同性别鸳鸯日行为频次分配的比较可以看出,雄性的主要行为是静栖和站立,且站立时间显著高于雌性,而雌性觅食时间显著高于雄性,因为雄性不参与孵化[26],孵卵和育雏职责完全由雌性承担,没有雄性的递食和换孵,雌性所有的能量来源均靠自给[15]。这种现象导致雄性更倾向于休息,雌性则把更多的时间用于除人工投喂外的觅食行为,为孵卵和育雏提供充足的能量。鸳鸯已连续多年在兆麟公园繁殖,且兆麟公园人流量较大,鸳鸯长期处于人类活动干扰的环境下,因此适应性地降低了警戒行为的强度[27]。本研究中记录到的警戒行为绝大多数是雌性对于其他个体进入巢树树洞而表现出来的防御行为,曾记录到攻击、威胁其他入侵个体的现象,而对于游人等产生的警戒行为较少。兆麟公园鸳鸯的打斗行为多出现在人工喂食时争抢食物或繁殖期配对过程中异性之间的挑衅,其他因素引发的打斗行为较少,这一现象可能是雌鸳鸯的择偶标准发生了变化,抑或是对环境适应的表现,有待于进一步研究。
从鸳鸯卵期和雏期日行为频次分配的比较可以看出,卵期游走和觅食显著高于雏期,而雏期的站立和理羽显著高于卵期。原因可能是来此的野生鸳鸯经过长距离迁徙后,雌雄个体需先进行能量补充,一旦配对后,雌性则更倾向于消耗更多的时间投入到觅食等相关行为中,以便积蓄更多能量,为后续的孵卵、育雏做准备。育雏只由雌性个体承担,在观测点观察到的雄性个体多单独活动,昼间多用于静栖等休息行为,这使得站立和理羽等休息行为占比较大。
从鸳鸯日行为频次分配的成幼差异可以看出,成体在静栖、站立和理羽等行为频次的占比显著高于幼体,而幼体的游走和觅食行为频次的占比显著高于成体。原因是幼体在生长发育阶段,对食物的需求量较大,仅依靠早、中、晚的人工投喂不能满足生长发育的要求。相对应的以觅食为目的的游走行为在幼体中频次较高。由于已适应人类活动,鸳鸯成体在警戒和打斗日行为频次的分配上低于幼体,这一结论与其他研究结果[28]相似。
每一个物种都有自己的行为频次分配和节律,一个与环境相适应的活动节律有助于维持物种更好的生存状态[29]。据本次研究,卵期和雏期鸳鸯的打斗、觅食和静栖行为都有明显的节律性变化,这或与觅食行为的节律变化相关。清晨,动物需要立刻补充能量进食,只有在身体能量得到保证的前提下才会发生理羽和游走等行为[30]。由于兆麟公园在早、中、晚人工投喂,导致鸳鸯的觅食节律亦对应出现3 个峰值,呈明显的周期变化。在人工投喂过程中,个体间因为争夺食物常常出现打斗行为,相比之下其他时间内打斗行为出现较少。由此,打斗行为与觅食行为亦呈现明显的正相关。觅食结束后,鸳鸯通常会休息,静栖和理羽行为便成为主要的行为类型[31]。鸳鸯日间活动几乎全部集中在水面和栏杆上,游走和站立便成为其余各类行为的衔接,而飞行行为属高能耗的行为方式,除非遇惊扰而逃避或改变活动区域,否则飞行很少发生,相对应的,游走行为就成为主要的过渡性行为类型。
据笔者观察,卵期和雏期鸳鸯都有较明显的领域行为,雄性会对游走在其周围的单只或成对鸳鸯张嘴鸣叫或伸长脖子鸣叫,甚至会从栖息的水面飞起驱逐,雌性也会对侵入孵卵树洞的鸳鸯进行警戒和示威,但不如雄性表现激烈。从不同性别鸳鸯日行为节律的比较可以看出,雄性鸳鸯打斗行为整体维持在较高水平,且具有偶发性,与日时段无关,主要取决于其他雄性鸳鸯的入侵;虽然雌性鸳鸯打斗行为记录较少,但在警戒行为频次上要高于雄性。雄性的领域行为充分表现出了对食物、配偶方面的维护,雌性的警戒行为则在卵和雏鸟的保护上频繁发生,这种行为模式在野生赤膀鸭(Mareca strepera)的行为研究中同样存在[24]。游走、觅食、静栖和理羽等行为差异不明显。
通过对鸳鸯卵期和雏期行为节律比较可知,2个时期的静栖、理羽、站立和游走行为总体趋势一致;卵期觅食行为结束之后,警戒行为达到峰值,打斗行为也进入高潮。这或是因为人工投喂的过程中,所有个体均混群于同一投饲场,除个体间争抢食物以外的其他攻击行为被忽略掉,但当喂食结束,鸳鸯会恢复配对,恢复领域行为,进而警戒行为达到峰值,打斗行为也进入高潮。另外,雏期的领域行为随着雏鸟体尺生长发育逐渐减弱,打斗和警戒行为发生频率亦下降。
幼体的日行为节律在游走和觅食上占比均匀,其他行为波动较大。这是因为雏期幼体处于生长发育阶段,食物需求量大,几乎全天都要进行食物的大比例摄入以满足身体发育需求,觅食行为自然成为主要行为类型,其他行为则受到干扰时才出现。人工饲喂后,成体会出现觅食行为以外的其他休息及社群行为,且各类行为呈周期性波动。此外,由于内、外条件的限制,本研究只进行了单次记录,未做重复检验,这是今后要弥补的;设计中对部分行为未做精细划分,这也是在深入研究中要修正的;对行为记录对象未做必要的标记,这是未来研究中要精准的方向。
