哈密市鹅喉羚夏秋季种群数量与生境适宜性评估
2023-11-25董培培高文佳程勇翔赛甫拉苏巴阿不都艾尼克力木王蕊蕊夏米西丁阿不都热依木
董培培,高文佳,程勇翔,赛甫拉·苏巴,阿不都艾尼·克力木,王蕊蕊,韩 廷,夏米西丁·阿不都热依木,4,5
(1.石河子大学生命科学学院,石河子,832003;2.哈密市林业和草原局(野生动植物保护中心),哈密,839099;3.伊吾县林业和草原局野生动植物保护中心,伊吾,839300;4.北京大学生命科学学院,北京,100871;5.石河子大学绿洲城镇与山盆系统生态兵团重点实验室,石河子,832003)
鹅喉羚(Gazella subgutturosa)隶属于鲸偶蹄目(Cetartiodactyla)牛科(Bovidae)羚羊属(Gazella),分布于欧亚大陆半干旱区,是一种典型的荒漠、半荒漠有蹄类物种。因雄性鹅喉羚在发情期喉部会膨大(即甲状腺肿),形似鹅喉,故称鹅喉羚。鹅喉羚已被列为国家二级重点保护野生动物[1],IUCN 红色名录易危(VU)级[2],《中国脊椎动物红色名录》易危(VU)物种[3]。鹅喉羚在中国境内分布于新疆准噶尔盆地、塔里木盆地、吐哈盆地、甘肃、青海和内蒙古等地,对海拔变化适应较强,从200 m的盆地到3 500 m的高原均有分布。由于自然因素(全球气候异常)和人为因素(牧业发展、修建水渠、道路建设)等影响,导致该物种栖息地缩减,种群数量也迅速减少[1,4-5],生境丧失和破碎化是加速物种灭绝和生物多样性下降的重要原因[6]。
遗传学研究表明,鹅喉羚新疆地理种群具有较低的核苷酸多样性[7-9],且南疆和北疆种群具有较高的遗传分化[8-11]。相比鹅喉羚新疆其他种群,哈密种群具有较高的单倍型多样性和较低的核苷酸多样性,并表现出较高的遗传分化[8]。在系统发生关系上,相比北疆种群,哈密种群与南疆种群相近。线粒体研究表明北疆和南疆种群之间存在基因流[10]。从地理分布看,哈密市处于东天山末端,由于天山山脉将准噶尔盆地(北疆)和塔里木盆地的鹅喉羚种群(南疆)隔开,所以哈密种群可能在这2 个种群的基因交流中起着重要作用,同时,哈密种群可能在新疆种群与青海、甘肃和内蒙古种群相互基因交流中扮演桥梁作用。
种群数量大小对于物种保护状态的评估极其重要,是IUCN红色名录和国家重点保护动物名录制定的基本指标,是合理保护野生动物和制定管理对策的基础[12-13]。在伊犁河谷,主要针对鹅喉羚种群密度和种群结构开展调查,发现雌性和雄性的种群占比为52.0%和37.9%,且随季节变化[14]。蒙古国鹅喉羚种群数量为(6 046±1 398)只[15]。种群密度对平均群体大小有相当弱的影响。在国内,针对鹅喉羚种群数量的调查研究主要开展于1990 年代[16];近年来,主要对卡拉麦里山国家级自然保护区[17-18]和艾比湖国家级湿地自然保护区[19]的鹅喉羚开展不同季节种群数量调查,而针对非保护区区域的调查较少。高行宜等[20]2004年对哈密盆地鹅喉羚种群数量进行考察,估算鹅喉羚总量为(37 108±4 420)只,但只调查哈密分布范围的一小部分,时间上距今已近20年,种群大小可能有所变化。
生境是物种生存、繁殖及种群发展的重要条件,生境质量显著影响物种的分布、种群密度、繁殖率及存活率[21],因此,保护物种不仅仅是维持其数量,更需要保护其生活的环境[22-23]。MaxEnt模型对野生动物栖息地的分布预测是根据现分布所联系的环境变量计算其分布的最大熵来对潜在分布区进行合理预测[24-26]。该模型的优点是仅需要物种的实际分布数据就可对物种的栖息环境进行评估和预测,尤其在物种分布数据不全或很少的情况下,依然能取得良好的结果[27-28]。新疆是鹅喉羚的重要分布区,近些年对新疆鹅喉羚适宜生境的预测结果都表明哈密是适宜分布区之一[29-30],而且潜在的适宜生境都靠近山脉[31]。保护区内人为影响小,因此对保护区内鹅喉羚适宜生境进行预测[32-34],与非保护区相比还有差异。近些年,哈密市快速发展,城市扩张、气候变化等人为和自然因素对鹅喉羚适宜生境产生不利影响。对哈密市鹅喉羚适宜生境进行分析,并对鹅喉羚适宜生境进行重点保护,有助于鹅喉羚种群数量的恢复与增长,因此,本研究扩大了研究区域,对哈密市进行调查。采用样线法获取鹅喉羚分布的基础信息,并结合MaxEnt 模型对鹅喉羚适宜生境进行评价,分析其与主要环境因子间的关系,采用Distance软件估算鹅喉羚种群数量、分析种群变化趋势,为该区域鹅喉羚长期监测及有效保护管理与利用提供依据。
1 研究区概况
哈密市位于新疆东部,总面积约14.2万 km2,地跨东天山南北(40°52'—45°5' N,91°6'—96°23' E),属温带大陆性干旱气候,受天山山脉影响,南北气候差异明显,北部降水量100~250 mm,南部降水量30~50 mm,而天山山区降水量可达500~600 mm[35]。哈密市拥有众多国家级重点保护野生动物,如蒙古野驴(Equus hemionus)、雪豹(Panthera uncia)、秃鹫(Aegypius monachus)和金雕(Aquila chrysaetos)等国家一级重点保护野生动物,北山羊(Capra sibirica)、盘羊(Ovis ammon)、马鹿(Cervus canadensis)、狼(Canis lupus)、兀鹫(Gyps fulvus)、高山兀鹫(Gyps himalayensis)、靴隼雕(Hieraaetus pennatus)、黑鸢(Milvus migrans)、黑尾地鸦(Podoces hendersoni)和阿波罗绢蝶(Parnassius apollo)等国家二级重点保护野生动物。新疆哈密市共有鸟兽58 科221 种[36-37],共有高等维管束植物37 科111 属173 种,优势科现象明显,包括菊科(Asteraceae)、藜科(Chenopodiaceae)、豆科(Fabaceae)、禾本科(Poaceae)和蒺藜科(Zygophyllaceae)等9 科[38]。在荒漠地区,梭梭(Haloxylon ammodendron)、柽柳(Tamarix chinensis)等为建群植物。
2 研究方法
2.1 物种分布数据获取
2022 年7—9 月,在哈密市(伊州区、巴里坤哈萨克族自治县和伊吾县)采用样线法进行种群数量与物种分布调查[39-40]。鹅喉羚生活在视野开阔的荒漠,体形中等,容易观察,采用越野车作为交通工具进行调查。调查时,2 位具有野外调查经验的调查员坐在汽车两侧,使用8×42 Bushnell 双筒望远镜观察,车速不超过20 km/h,遇到鹅喉羚实体时,记录GPS 位点、性别、年龄(成体、亚成体(包括幼体))和数量,通过经验判断鹅喉羚距样线的垂直距离,2 个及以上个体的距离为其中心的垂直距离。每条样线仅记录第1 次遇见的鹅喉羚。当遇到鹅喉羚粪便[41-42]、足迹及尸体时,记录GPS 位点,仅作为物种分布点使用。
2.2 环境变量数据
依据前人对鹅喉羚适宜生境预测的研究结果及鹅喉羚的生活习性,选择地形因素(海拔、坡度、坡向)、气候因素(19个生物气候变量)、植被因素(归一化植被指数NDVI)和人为干扰因素(距道路距离)共4 类24 个环境因子作为鹅喉羚生境评价的影响因子[29-30]。地形数据来自30 m 的数字高程模型(DEM,中国科学院计算机网络信息中心地理空间数据云平台http://www.gscloud.cn)提取的海拔、坡度和坡向;气候数据来自世界气候数据库WorldClim 网站(https://www.worldclim.org)19 个生物气候变量,空间分辨率为30″;植被因素来自资源环境科学与数据中心(http://www.resdc.cn);人为干扰变量由国家基础地理信息系统提供的1∶100 万地理信息数据获取。所有环境变量均通过重采样转化为30 m×30 m分辨率的ASCII栅格数据,并统一为相同的坐标系统。
为减少上述环境变量多重共线性的影响,用SDM工具包[43]中的raster correlation and summary statisics 工具剔除空间相关性过高(|r|>0.8)的环境变量,最终筛选出11 个环境变量(bio1、bio3、bio10、bio11、bio15、bio19、NDVI、人为影响、海拔、坡度和坡向)用于构建鹅喉羚生态位模型。
2.3 MaxEnt模型
采用MaxEnt 3.4.4软件[44]对鹅喉羚的潜在分布区进行预测,将物种分布点数据和环境变量数据导入MaxEnt 模型中,将最大迭代次数设置为2 000,Bootstrap 重复运算10 次,并开启random seed,勾选绘制响应曲线(response curve)与刀切法(Jackknife检验)功能分析影响鹅喉羚分布的环境变量的重要性。随机选取75%的分布点数据用于建立模型,剩下25%的分布点数据用于模型检验,构建鹅喉羚生境模型进行模拟,取10 次模拟结果的平均值作为最终模拟结果[45]。
采用受试者工作特征曲线(receiver operating characteristic curve,ROC 曲线)法以评估模型的精确度[46]。ROC 曲线下的面积为AUC(area under curve)值,AUC值越大,表示环境变量与预测物种地理分布模型之间的相关性越大,越容易分辨该物种是否分布,预测效果越好。AUC 取值为[0,1],评价标准为 >0.5~0.6,失败;>0.6~0.7,较差;>0.7~0.8,一般;>0.8~0.9,好;>0.9~1.0,优秀[47]。
2.4 潜在适宜分布区划分
根据选择模型最大训练敏感性和特异性(maximum training sensitivity plus specificity,MTSS)、平衡训练遗漏率(balance training omission)、预测面积及阈值(predicted area and threshold value,TPT)的结果,将鹅喉羚的生境适宜性分成3 个等级:不适宜、次适宜和适宜,最终得到鹅喉羚在哈密市的潜在适宜区分布图。
2.5 种群数量研究方法
运用Distance 7.5 软件[48]提供的模型计算鹅喉羚种群密度。通过探测函数g(x)计算垂直距离x处的鹅喉羚或鹅喉羚群体被发现的概率,得到鹅喉羚种群密度。探测函数由主函数和级数展开两部分组成,主函数包括均匀分布、半正态分布、风险率和负指数分布4 种统计分布;级数展开包括余弦、简单多项式和赫米特多项式3 种,用来调整上述主函数。最后再结合保护区适宜生境的面积计算鹅喉羚种群数量。模型的选择以最小爱氏信息准则(AIC)值来确定[49]。
3 结果
3.1 分布数据
调查共记录哈密市鹅喉羚分布点312个(图1),数据分辨率约为1 km,因此设定缓冲半径为0.5 km[50]。为减少物种分布建模中使用的点空间自相关性的影响,利用ArcGIS 插件“SDMtoolbox”[43]中的spatially rarely occurrence data for SDMs 工具,对哈密市鹅喉羚原始分布点进行1 km 间距的空间处理,共筛选出160个分布点用于构建MaxEnt模型。
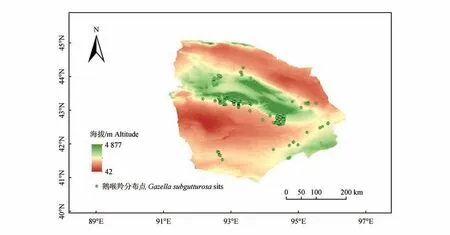
图1 研究区与鹅喉羚分布位点Fig.1 The map of study area and main distribution areas of Gazella subgutturosa
3.2 MaxEnt模型评价
哈密市鹅喉羚生境预测模型的平均训练集AUC值为(0.939±0.017)(图2),表明MaxEnt 的结果精确度达到优秀水平,适合用于哈密市鹅喉羚的生境适宜性评价。
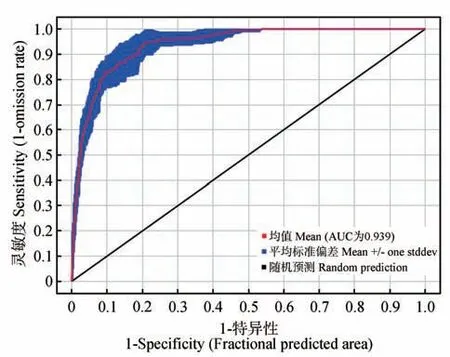
图2 MaxEnt模型的ROC曲线Fig.2 ROC curve of MaxEnt model
3.3 环境因子对鹅喉羚生境的影响
运用Jackknife测试法计算每个变量的相对重要性和贡献率。对鹅喉羚生境影响较大的是人为影响、海拔、等温性(bio3)和最冷季平均气温(bio11),贡献率分别为29.0%、17.7%、11.5%和11.1%(图3,表1),累计贡献率达69.3%,表明这4 个环境因子对鹅喉羚的分布具有重要影响。
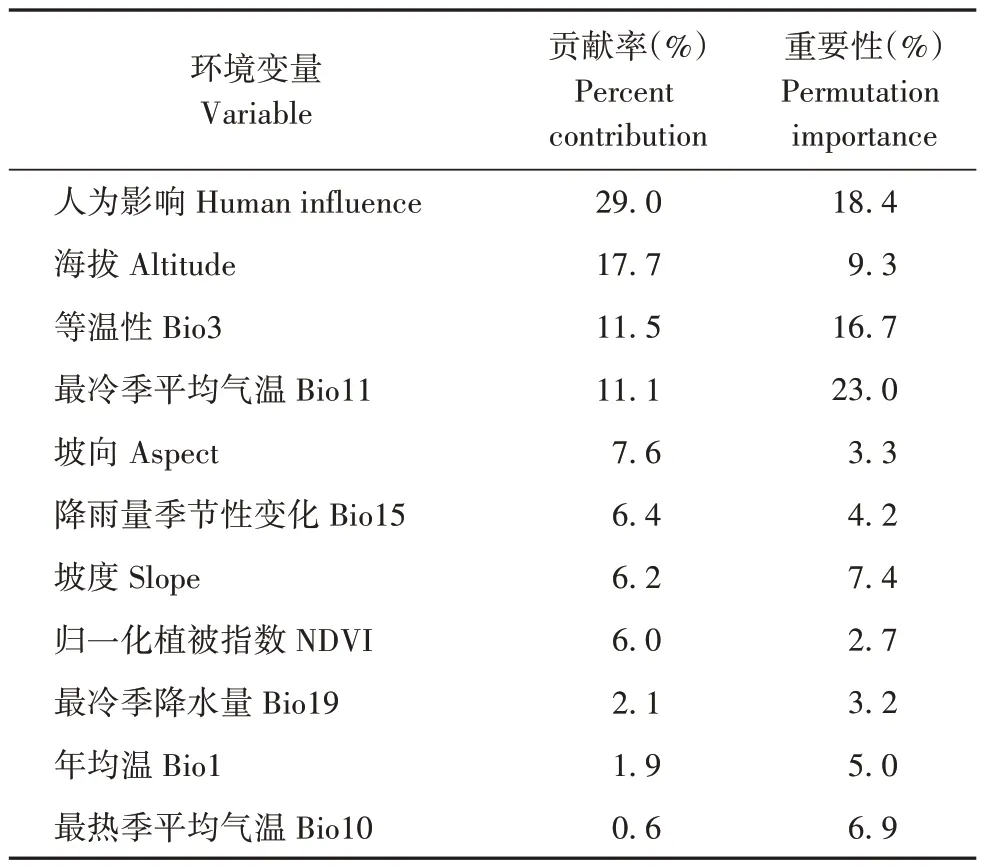
表1 影响鹅喉羚分布的重要环境变量Tab.1 Important environmental variables affecting the distribution of Gazella subgutturosa
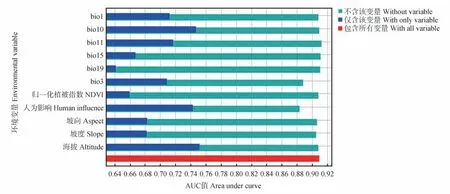
图3 刀切法检验各环境变量的重要性Fig.3 Importance of environmental variables tested by knife cutting method
通过鹅喉羚生境适宜性对主要环境变量的响应曲线(图4)发现,鹅喉羚偏好距道路较近的区域,这可能是对于一些乡道等土路,鹅喉羚并不受影响,但人为修建水渠往往集中在道路两侧,对其分布存在影响;鹅喉羚在海拔1 500 m的区域分布最多,海拔过低或过高都对分布存在影响;鹅喉羚偏好等温性在29 左右的区域活动;鹅喉羚偏好最冷季平均气温不低于-9 ℃的区域,鹅喉羚皮毛短少,并不能适应严寒气候。
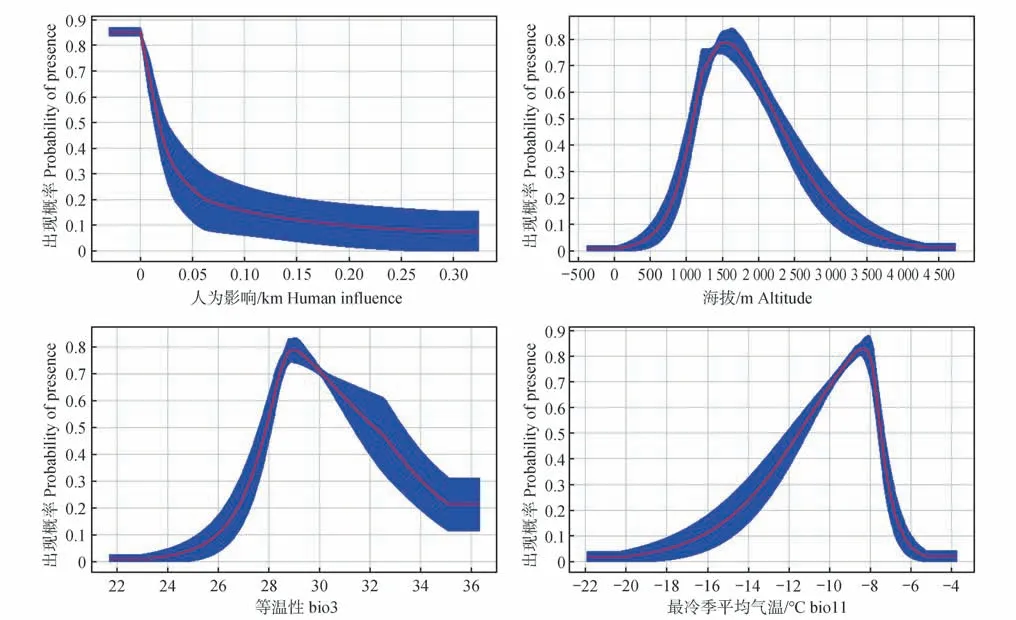
图4 生境适宜性对主导环境变量的响应曲线Fig.4 Response curves of habitat suitability to dominant environmental variables
3.4 潜在适宜生境分布
MaxEnt模型的MTSS和TPT值分别为0.140 1和0.411 9。因此,鹅喉羚适宜生境和次适宜生境的划分阈值分别为0.140 1 和0.411 9。采用上述阈值对模型输出结果进行重分类,得到哈密市鹅喉羚潜在适宜生境分布(图5)。经计算,哈密市鹅喉羚的适宜生境面积为1.725 9 万 km2,占哈密市面积的12.15%,主要位于天山南侧的荒漠地区及西南和东北海拔较高的区域;鹅喉羚次适宜生境面积为3.401 0万 km2,占哈密市面积23.95%,主要分布于适宜生境周边地区;不适宜生境面积为9.073 7万 km2,占哈密市面积和的63.90%,主要分布于哈密市低海拔及山脉等高海拔地区。
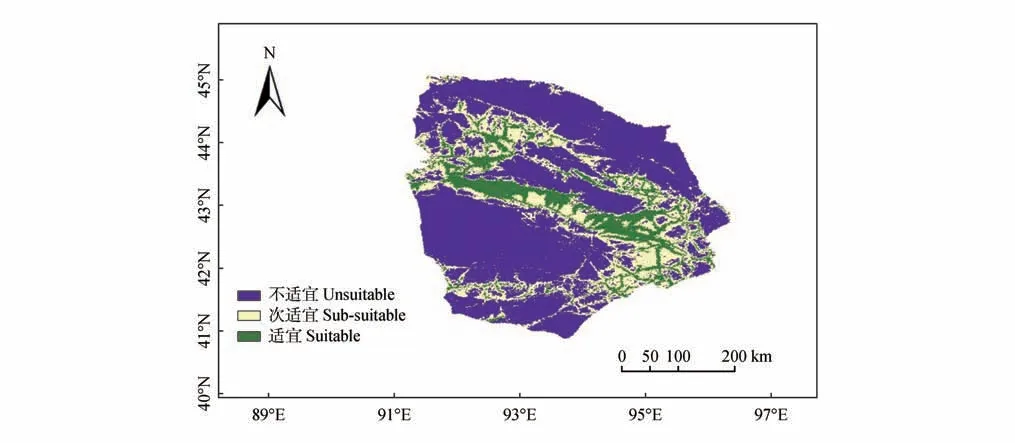
图5 哈密市鹅喉羚生境适宜性分布Fig.5 Habitat suitability distribution map of Gazella subgutturosa population in Hami
3.5 种群数量及结构
在2022 年夏季对鹅喉羚数量展开调查,在总长1 303 km 的59 条样线中,共发现303 群,1 029 只鹅喉羚个体。基于最小AIC 值得到最优探测函数组合为风险率探测+简单多项式(hazard-rate+simple polynomial)(表2)。根据该模型,调查样线的有效半宽(ESW)为181.68 m,哈密市鹅喉羚的种群密度为(2.013±0.432)只/km2(95% 置信区间为1.321~3.070 只/km2),平均种群大小为(3.396±0.161)只(95%置信区间为3.094~3.727 只),预期种群大小为(3.158±0.140)只(95% 置信区 间为2.895~3.445 只)。结合哈密市鹅喉羚适宜生境面积,得到种群数量为(34 742±7 455)只(95%置信区间为22 793~52 978只)。
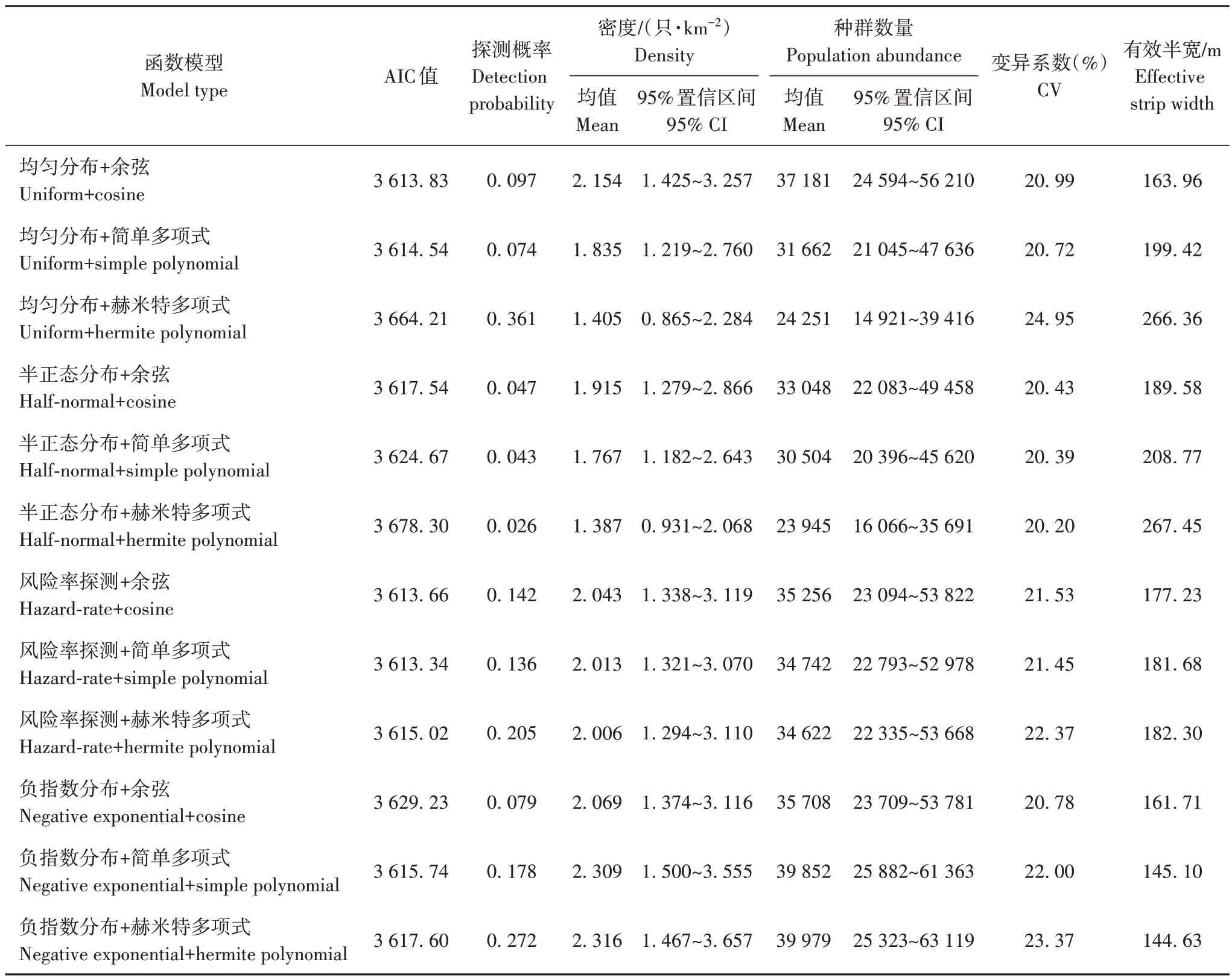
表2 鹅喉羚种群密度和估计Tab.2 Population density and estimate of Gazella subgutturosa in Hami,Xinjiang
将鹅喉羚集群分为雄性群(只包含雄性个体)、雌性群(只包含雌性个体)、混合群(包含雄性、雌性和亚成体)、雌雄群(只包含雄性和雌性成体)、雌亚群(只包含雌性和亚成体)、亚成体群(只包含亚成体)和独亚、独雄、独雌及未知(未能辨认性别)。在各类型群中,雌亚群出现次数最多,占35.97%,其次是雌性群,占16.83%(表3)。在此次调查中,共发现鹅喉羚303 群,平均群大小为3.396 只,最大群为21 只,最小群为1 只,多集中在10 只以下(图6)。将可辨别出性别、年龄的个体分为雄性成体、雌性成体和亚成体,其中雄性占15.11%,雌性占56.32%,亚成体占28.57%。
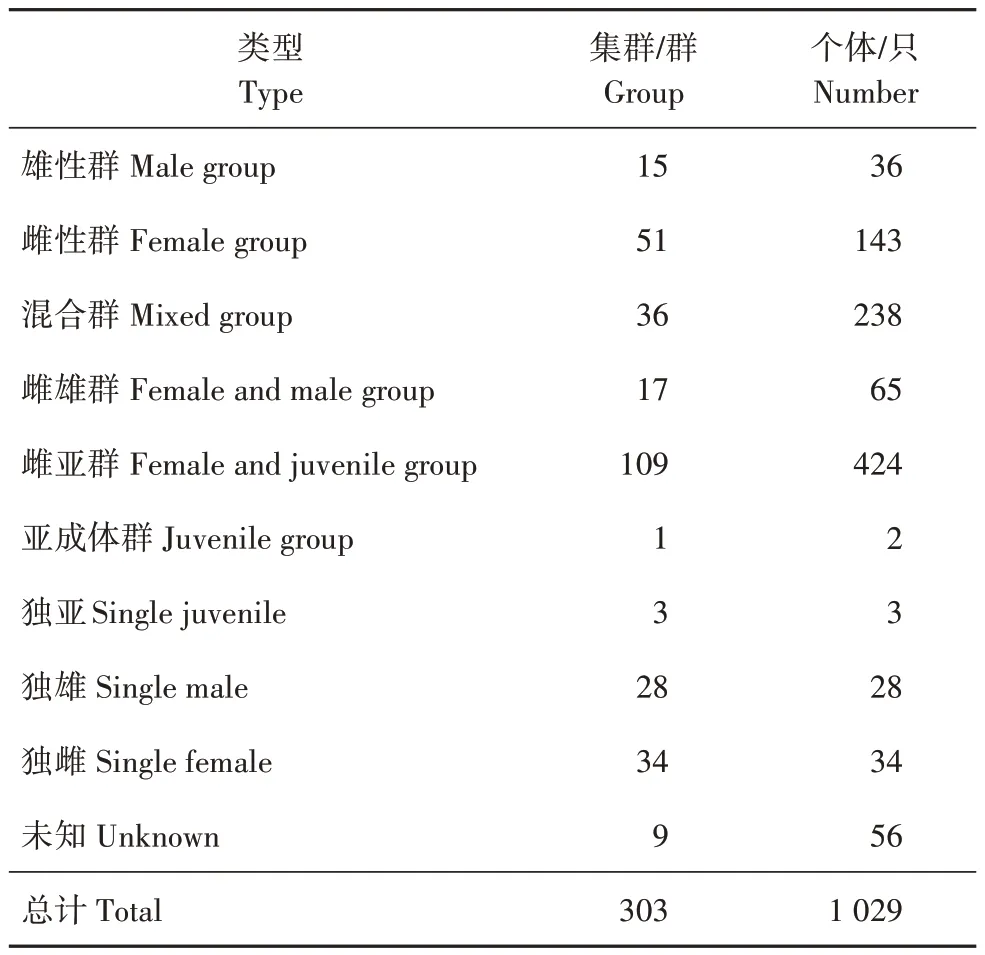
表3 鹅喉羚哈密种群年龄、性别结构Tab.3 Gender and age structure of Gazella subgutturosa Hami population
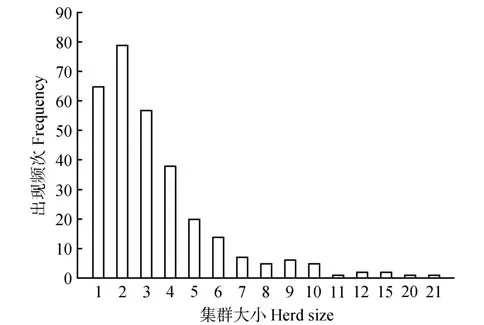
图6 鹅喉羚哈密种群集群大小及其频次Fig.6 Herd size and frequency for Gazella subgutturosa Hami population
4 讨论
4.1 影响潜在适宜生境的因素
本研究发现,对鹅喉羚分布有重要影响的生物因子是最冷季平均气温(bio11)和等温性(bio3),这与前人研究结果[30-31]相似,可能是鹅喉羚皮毛短少,抵御寒冷的能力差,所以偏好分布在气温变化不大的区域。在地形因素中,对鹅喉羚分布影响最大的是海拔,这与苏比奴尔·艾力等[29]的结果相同,鹅喉羚集中分布在1 500 m 左右的区域,可能是夏季炎热,中高海拔地区温度较低,并且低海拔地区植被过于稀少,所以鹅喉羚分布于海拔较高区域。坡度和坡向对鹅喉羚的分布影响不明显,主要是鹅喉羚行动敏捷,活动范围广阔,对地形要求不高。
人为因素是影响鹅喉羚分布的重要因素。人为因素主要为距道路距离,道路包含高速、国道、省道、县道、乡道及其他道路。鹅喉羚偏好分布于道路的周围,可能是修建水渠的影响,水渠分布在道路两侧,而鹅喉羚的生存离不开水,并且我国对野生动物的保护经过多年的管理,野生动物已慢慢接受这种境遇,这与程亮等[34]对艾比湖保护区鹅喉羚生境适宜性评价的结果类似。鹅喉羚采食地的选择主要受植物种数和植被盖度的影响[51],过密过高的植被会阻碍视野,不利于发现天敌,因此鹅喉羚不会在植被覆盖度过高的区域觅食。采用MaxEnt 模型得到鹅喉羚在哈密市的适宜生境面积为1.725 9 万 km2,占哈密市面积的12.15%。通过高行宜等[20]在2004 年调查鹅喉羚的分布点计算哈密市适宜生境面积为1.950 0 万 km2,本研究结果与之相比适宜生境面积减少,不适宜生境增加。适宜分布的生境向天山脚下靠近,可能是近些年来哈密市的发展进一步压缩鹅喉羚的生存面积。有时甚至可以看到鹅喉羚在村庄周围活动。人类活动不仅减少了鹅喉羚的适宜生境面积,而且导致生境破碎化,阻碍了种群之间的基因交流,长此以往,使得遗传多样性降低,数量减少,严重危害了鹅喉羚的生存与繁衍。
4.2 鹅喉羚种群数量和结构的变化
本研究使用样线法进行种群数量调查,样线法作为大范围内开展大、中型有蹄类动物数量调查的基本方法,特别适用于平坦开阔、视野较好的区域[39],并且使用Distance 软件选择最适合的探测函数,也提高了预测精度[52]。样线法要求调查时尽可能取得不同垂直距离的信息,垂直距离是影响概率密度函数的关键变量。但在实际调查中不可能准确测量遇见的每个个体或群体到样线的垂直距离,因为在调查过程中,与鹅喉羚相遇时,其会迅速逃跑。因此,观测个体与样线的垂直距离主要以目测为主,目测的准确性主要与观察者的经验、疲劳程度及天气有关,这些都能影响探测函数。调查时要根据实际情况适当校正观测值。通过Distance 软件进行分析,可以优选出拟合度最高的探测函数,得出较为精确的种群密度。即使小的偏差不可避免,但不同调查的结果会相当一致[52]。再依据MaxEnt模型得出适宜生境面积计算种群数量,以此确保结果的准确性。
本研究调查哈密市鹅喉羚密度为(2.013±0.432)只/km2,结合哈密市鹅喉羚的适宜生境面积,得到其种群数量为(34 742±7 455)只。从种群数量来看,鹅喉羚的数量由其密度与适宜生境面积所决定,适宜生境面积又是种群数量的重要影响因素,而根据MaxEnt 模型可以很好地预测鹅喉羚在哈密市的适宜生境[27-28]。从种群密度来看,哈密市鹅喉羚种群密度和数量均高于卡拉麦里山和艾比湖自然保护区[17-19]。而在2004 年,高行宜等[20]通过截线抽样法对哈密盆地鹅喉羚的种群数量进行调查,发现高密度区为(3.330±0.240)只/km2,中密度区为(0.950±0.055)只/km2,而低密度区为(0.056±0.042)只/km2,鹅喉羚总数为(37 108±4 420)只。相比之下,本研究扩大了研究区域,但鹅喉羚数量却更少,这表明在近20 年哈密市鹅喉羚种群数量大幅下降。首先,高行宜等[20]采用样线法结合研究区域面积计算鹅喉羚数量,本研究采用样线法与MaxEnt 模型结合,计算适宜生境面积,得出鹅喉羚数量,所以产生差异;其次,虽然国家对于野生动物的保护在近些年越来越重视,但在调查过程中,发现人为修建水渠严重破坏了植被的生长,进而影响鹅喉羚的生存。而且在鹅喉羚分布较多的区域,如沁城、柳树泉等地,鹅喉羚与家养牛羊共处一地,与其食性相似[42,53],存在食物竞争。另外,在调查过程中多次遇见鹅喉羚被撞死在路边,而且距离道路越近,鹅喉羚投入警戒时间越多[54]。哈密拥有数量众多的鹅喉羚,但缺乏系统保护,近10 年数量明显下降,究其原因,还是人为因素占据主导地位。
本研究结果表明,鹅喉羚多以单独或2~10 只的小群活动,占总数的89.70%,这与前人研究结果[55-56]相似,小群鹅喉羚更有利于觅食,减少被天敌发现的风险。鹅喉羚的主要集群类型为雌性群和雌亚群,调查时鹅喉羚生产时间刚过,雌亚群的数量最多,并且双羔率达28.40%。鹅喉羚青年和老年个体多产单羔,3~7 岁的雌性多产双羔[57],因此,哈密市雌性鹅喉羚年龄多为青年(1~3 岁)和老年(7 岁以上)。而在性别比例方面,雌∶雄=3.728∶1,雌∶亚=1.971∶1,这与高行宜等[56]于1992 年对新疆鹅喉羚种群结构调查的结果不同,可能是与鹅喉羚在冬季发情,翌年春天产下幼仔,雌性独自带领幼体生活有关。调查过程中发现独羚75 次,占总计的24.75%,乔建芳等[55]对新疆卡拉麦里山保护区鹅喉羚的研究也有类似情况,这可能是为了避免被捕食的风险。
4.3 展望与建议
虽然哈密市目前没有设立专门的鹅喉羚保护区,保护区的建立可以有效对物种进行保护,也可能造成鹅喉羚栖息地的破碎化,产生种群之间的隔离,阻碍基因交流。哈密市已修建的交通路线有G7、G30 高速公路和兰州到乌鲁木齐市的高速铁路和货运铁路,已建和新建的交通路线给鹅喉羚分布与生境带来一定威胁,未来如果建立保护区,需注意保护区之间的连通性。此外,哈密市尤其是巴里坤县和伊吾县主要以放牧为主,家畜主要有绵羊、山羊、骆驼、马和牛等,这些家畜与鹅喉羚有食物竞争,并有疾病传染等威胁,因此需要进一步评价其影响。
基于本研究的鹅喉羚种群数量、结构和生境评价结果,结合调查过程中发现的相关问题,提出建议:(1)设置禁牧区,严格管理家畜,避免与鹅喉羚竞争食物资源,在野外调查期间,经常看到家畜与鹅喉羚共同进食,食物竞争是造成哈密市鹅喉羚种群下降的重要原因。(2)在鹅喉羚分布较多区域放置警示牌,设置减速带,防止鹅喉羚横穿道路被撞。(3)水渠修建应考虑鹅喉羚饮水,或者设立饮水点,便于饮水,保障基本需求,以减少对鹅喉羚种群的影响。
