三种壳色硬壳蛤(Mercenaria mercenaria)的生长性状差异及通径分析
2023-11-21王元宁李华煜李金泽董诗雨李永仁郭永军
王元宁, 李华煜, 孟 良, 李金泽, 董诗雨, 李永仁, 郭永军, 张 涛
三种壳色硬壳蛤()的生长性状差异及通径分析
王元宁1, 2, 李华煜1, 2, 孟 良1, 2, 李金泽1, 2, 董诗雨1, 2, 李永仁1, 郭永军1, 张 涛2
(1. 天津农学院 水产学院 天津市水产生态及养殖重点实验室, 天津 300384; 2. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
壳色是影响商品贝类经济价值的重要性状。本研究以不同壳色的硬壳蛤()群体为繁殖亲贝, 采用个体间随机交配的方法, 成功选育了白、红和杂3种壳色的子代群体, 并在池塘培育至360日时测量了其壳长(1)、壳高(2)、壳宽(3)及活体质量(), 并使用相关性分析、主成分分析和通径分析等方法, 探究了不同壳色硬壳蛤在幼虫期和稚贝期的生长性状差异及壳形态性状(壳长、壳高、壳宽)对活体质量的影响。研究结果表明: 在幼虫期, 与白色和红色群体相比, 杂色群体生长速度更快、变态率更高, 但存活率低(<0.05); 在稚贝期, 白色群体表现出显著的生长优势(<0.05)。相关性分析结果显示, 3种壳色硬壳蛤的壳形态性状(1、2、3)与活体质量()的相关系数均达极显著水平(<0.01)。主成分分析和通径分析结果表明, 硬壳蛤的壳长(1)是影响其活体质量()的主要因素。本研究结果能够为不同壳色硬壳蛤良种选育提供重要理论依据和基础数据。
硬壳蛤(); 壳色; 生长性状; 通径分析
硬壳蛤()俗称美贝、美国红或美国螺, 属于双壳类、帘蛤科、硬壳蛤属, 是一种埋栖型双壳贝类, 其个体较大, 最大体重可超过200 g[1]。因其自然分布的地理纬度与我国纬度相似, 中国科学院海洋研究所张福绥院士等人于1997年首次从美国将硬壳蛤引种至我国[2], 并开展基础生物学和生理生态学等方面研究。硬壳蛤对高温、低盐的耐受能力强[2], 能够适应泥质和泥沙质环境, 适宜养殖于我国大部分地区的滩涂和海水池塘。经过20余年的示范和推广, 目前硬壳蛤已成为我国海水池塘养殖的重要经济贝类之一。
海洋双壳贝类壳色繁多, 壳色是海洋双壳贝类良种选育的重要指标, 壳色的不同可能影响其经济价值。大量研究表明, 壳色受遗传因素决定[3-4]。不同壳色的同种贝类在生长、存活和抗逆能力等方面表现出明显差异[5-7]。迄今为止, 以壳色和生长速度为选育指标的遗传育种实验大多聚集于马氏珠母贝[8]、海湾扇贝[9]、四角蛤蜊[10]和菲律宾蛤仔[11]等贝类。而在硬壳蛤中, 相关研究报道较匮乏。
亲贝选择是贝类遗传育种的重要环节, 壳形态性状(壳长、壳高、壳宽)和活体质量是亲贝选择的重要依据[12]。贝类的壳长、壳高和壳宽便于测量, 而活体质量的测量需借助电子称等工具, 且称重前需擦拭壳表面多余水分, 容易导致错误的测量结果[13]。为解决这一问题, 常亚青等[14]通过多元回归和通径分析调查了1龄虾夷扇贝中壳长、高和宽与活体质量之间的联系, 查明了与活体质量直接关联的壳形态性状, 对虾夷扇贝的良种选育具有重要意义。目前, 多元回归和通径分析已广泛应用于多种双壳贝类的良种选育中, 例如栉孔扇贝[15]、近江牡蛎[16]、日本镜蛤[17]和文蛤[18]。
本研究以白色、红色和杂色硬壳蛤群体为繁殖亲贝, 采用个体间随机交配的方法, 选育出3种壳色的子代群体, 并比较了不同壳色群体在幼虫期和稚贝期的生长性状差异。此外, 我们使用相关性分析、主成分分析和通径分析方法, 查明了影响活体质量的主要壳形态性状。本研究结果为硬壳蛤良种选育提供重要的理论依据和基础数据。
1 材料与方法
1.1 亲贝来源
硬壳蛤亲贝购自于天津滨海新区, 随后暂养于山东省莱州市长渔水产有限公司养殖车间(图1), 选择前我们从亲贝群体中随机选取200只杂色硬壳蛤(自然群体)作为对照组亲贝, 再按照个体发育良好、性腺饱满、壳型规整且无伤损的亲贝中挑选出白色和红色硬壳蛤各200只作为选育组亲贝, 其壳长、壳高、壳宽和活体质量数据见表1。

图1 白色、红色和杂色硬壳蛤
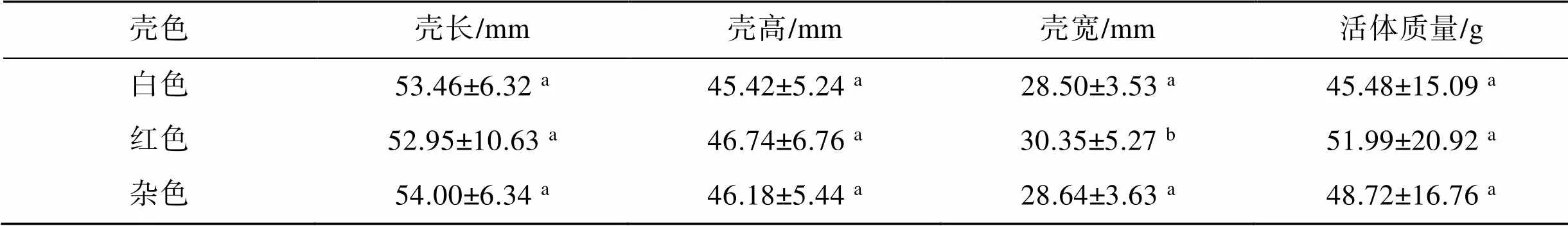
表1 硬壳蛤亲贝的壳长、壳高、壳宽和活体质量
注: 同一列中, 右上角标有不相同字母表示差异显著(<0.05)
1.2 产卵和培育
本实验采用个体间随机交配的方法, 选育出3种壳色硬壳蛤群体。我们挑选出白色、红色和杂色硬壳蛤亲贝各200个, 室内阴干5 h后, 将3组壳色亲贝放入新鲜海水的20 m³育苗池中充气流水刺激, 3 h后亲贝排精、卵。孵化期间, 海水温度为23~24 ℃, 盐度为25。
约24 h后, 受精卵发育为D形幼虫。我们测量幼虫数量后, 将其放入20 m3育苗池进行培养, 幼虫密度为4个/mL, 随幼虫生长定期调整密度, 实验设置3个平行。室内培育期间, 3组幼虫的水温、盐度、密度和饵料等培育条件均保持一致。当稚贝的壳长规格达到5 mm时, 将其移至室外池塘进行培育。在培育期间, 我们定期观察稚贝的生长状态并调整密度, 更换不同目数的网袋。
1.3 数据测量
幼虫期监测: 在第1、5、10和15日龄分别从各育苗池内随机取30个幼虫, 在显微镜下测量其壳长, 并统计第5、10日龄的存活率和第15日龄的变态率。其中, 存活率测定方法: 在3组育苗池内随机取样3次, 每次取3 mL加入碘酊溶液杀死幼虫后, 用显微镜统计幼虫个数, 并计算每次取样每毫升D形幼虫数占初始取样每毫升D形幼虫数的比例。变态率测定方法: 利用塑料长管吸取各池底幼虫, 在显微镜下统计出现次生壳幼虫的个数, 并计算第15日龄时单位体积出现次生壳的幼虫数量与面盘幼虫数量的百分比。
稚贝期监测: 从3种壳色硬壳蛤群体中随机取各50粒稚贝, 使用电子游标卡尺(精度0.01 mm)在第50、100、200和280日龄时测量其壳长, 第360日龄测量其壳长(1)、壳高(2)和壳宽(3); 利用电子天平(精确到0.01 g)称量其活体质量()。壳长为壳前端与后端间的最大距离, 壳高为壳顶部到壳腹缘的最大高度, 壳宽为壳左右两侧间的最大厚度[18]。根据下列公式计算3种壳色硬壳蛤的生长速度[19]。
= (1−0) / (1−0),
式中,0为第1次测量壳长,1为最后一次测量壳长,0为第一次测量日龄,1为最后一次测量日龄,为绝对生长率。
核聚变(核融合)需要极端温度才能工作。进行聚变反应时,两种形式的氢气将被注射到一个名为托卡马克装置的反应器中并且加热到1.5亿摄氏度,这个温度比太阳的温度还要高10倍。为了阻止反应器融化,过热的等离子体将借助地球上一些最强大的磁体悬浮在真空环境中。
1.4 数据分析
利用Excel 2016软件(Microsoft)统计3种壳色硬壳蛤生长性状测量结果, 导入SPSS 22.0软件(IBM SPSS)中获得各项表型参数统计量, 进行单因素方差(ANOVA)分析和Tukey多重比较分析。此外, 通过各性状间正态检验、相关性分析、壳形态性状主成分分析和通径分析, 查明硬壳蛤壳形态性状对活体质量的直接作用和间接影响, 并计算决定系数。通过多元逐步回归, 建立壳长、壳高和壳宽对活体质量的最优回归方程[20]。<0.05表明差异显著,<0.01表明差异极显著。
2 结果
2.1 三种壳色群体生长性状比较
3种壳色群体幼虫的平均壳长及生长速度见表2。1日龄时, 白色和红色群体壳长无显著差异(0.05), 杂色群体壳长显著小于白色和红色群体(<0.05); 5日龄至15日龄期间, 3种壳色群体壳长均呈现出较快的生长速度, 其中杂色群体的生长速度显著快于白色和红色群体(<0.05)。
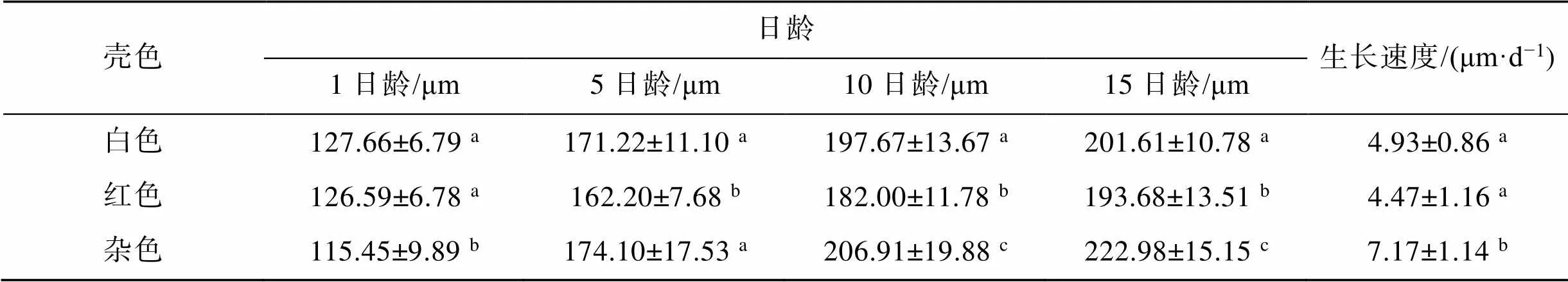
表2 三种壳色群体幼虫的平均壳长及生长速度
注: 同一列中, 右上角标有不相同字母表示差异显著(<0.05)
3种壳色群体幼虫的存活率及变态率见表3。2日龄至10日龄期间, 白色和红色群体的存活率显著高于杂色群体(<0.05), 但杂色群体的变态率(15日龄)显著高于白色和红色群体(<0.05)。

表3 三种壳色群体幼虫的存活率(2~10日龄)及变态率(15日龄)
注: 同一列中, 右上角标有不相同字母表示差异显著(<0.05)
3种壳色群体稚贝的平均壳长见表4。室内培育50日龄时, 白色和红色群体壳长无显著差异(>0.05); 100日龄时, 白色群体壳长显著高于红色和杂色群体(<0.05)。室外培育200日龄时, 红色群体壳长显著小于白色群体(<0.05), 杂色群体与白色群体壳长无显著差异(>0.05); 280日龄至360日龄期间, 白色群体壳长显著高于红色和杂色群体(<0.05)。
2.2 三种壳色硬壳蛤壳形态性状对活体质量的影响分析
本实验用Q-Q (Quantile-Quantile)检验法对360日时3种壳色硬壳蛤各性状数据进行正态检验。如表5所示, 所列各壳形态性状与活体质量的相关系数均达极显著水平(<0.01)。其中, 白色和红色硬壳蛤的壳形态性状与活体质量的相关系数大小顺序均为3>1>2; 杂色硬壳蛤壳形态性状与活体质量的相关系数大小顺序为1>3>2。从相关系数大小来看, 3种壳色硬壳蛤的壳长、壳高和壳宽性状间存在明显的共线性。
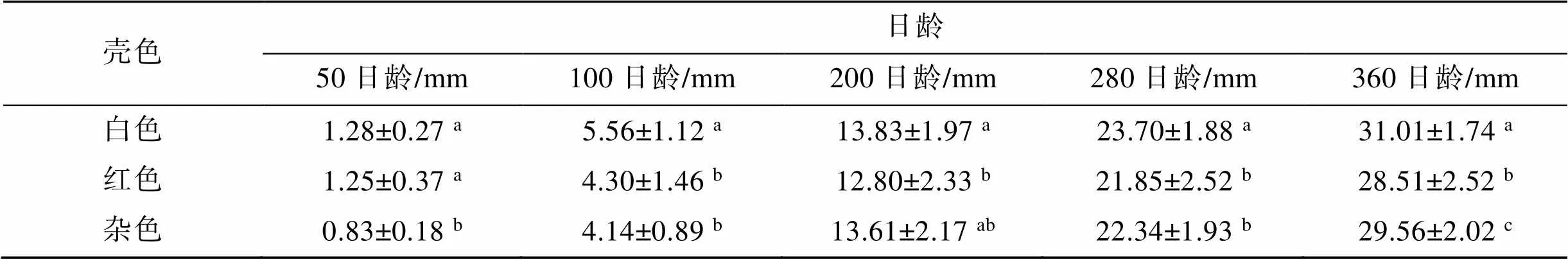
表4 三种壳色群体稚贝的平均壳长
注: 同一列中, 右上角标有不相同字母表示差异显著(<0.05)
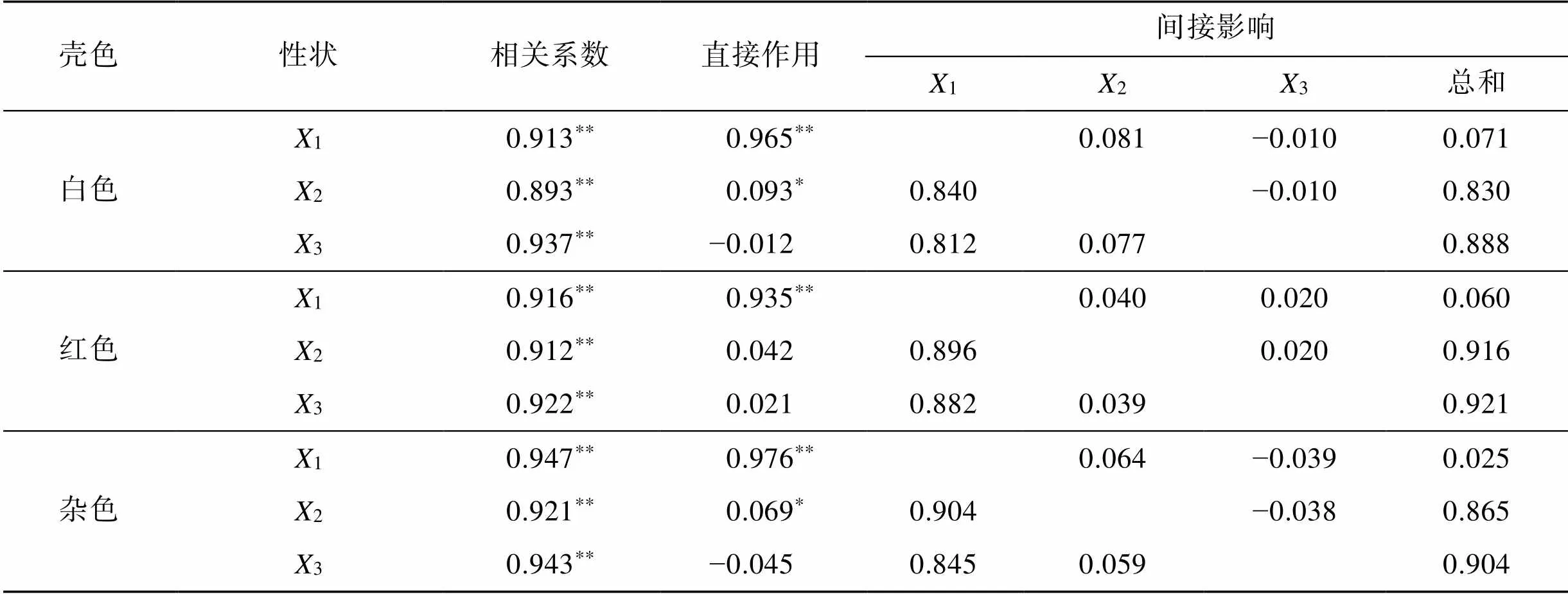
表5 三种壳色硬壳蛤壳形态性状对活体质量的通径分析
注:*表示差异显著(<0.05);**表示差异极显著(<0.01)
为使分析结果准确, 我们对各壳形态性状进行主成分分析。如表6所示, 3种壳色硬壳蛤只保留主成分1。白色硬壳蛤主成分公式为component 1= 0.9541+0.9492+0.9383; 红色硬壳蛤主成分公式为component 1=0.9861+0.9812+0.9763; 杂色硬壳蛤主成分公式为component 1=0.9711+0.9652+ 0.9433。不同壳色硬壳蛤主成分分析结果表明, 壳长、壳高和壳宽对该主成分均具有贡献价值。其中, 壳长对该主成分的影响最大, 而壳宽对主成分的影响较小。
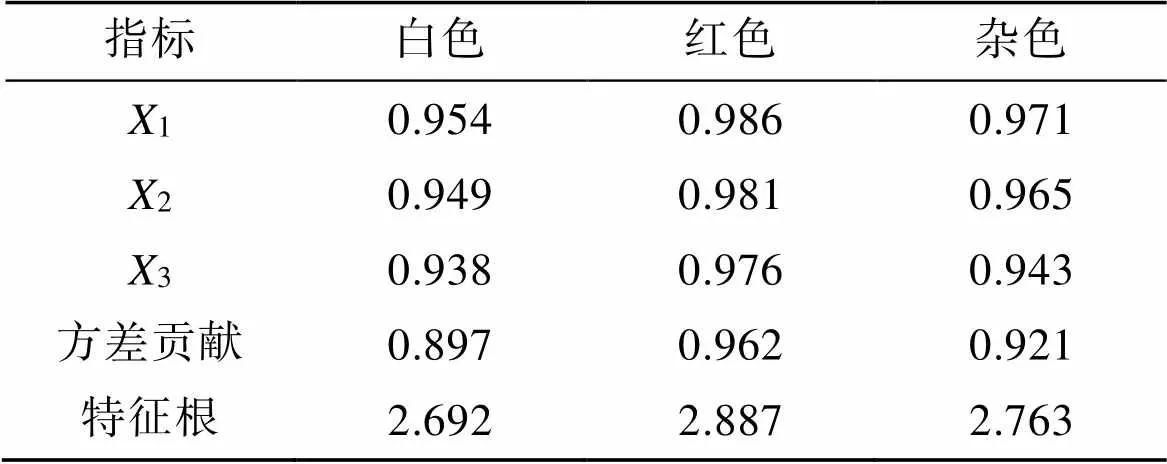
表6 三种壳色硬壳蛤数据主成分分析
单一性状对活体质量的决定系数为=2, 两两性状对活体质量的共同决定系数为= 2Pi[22]。各性状对活体质量的决定系数见表7。结果显示, 3种壳色硬壳蛤的壳长对活体质量的决定系数最大, 分别为93.1%, 87.4%和95.3%。白色、红色和杂色硬壳蛤总决定系数∑分别为1.075, 0.990和0.999, 其结果同各自相关指数2的数值近似相等, 表明上述所列3种壳色硬壳蛤的壳长、壳高和壳宽是影响活体质量的主要性状。
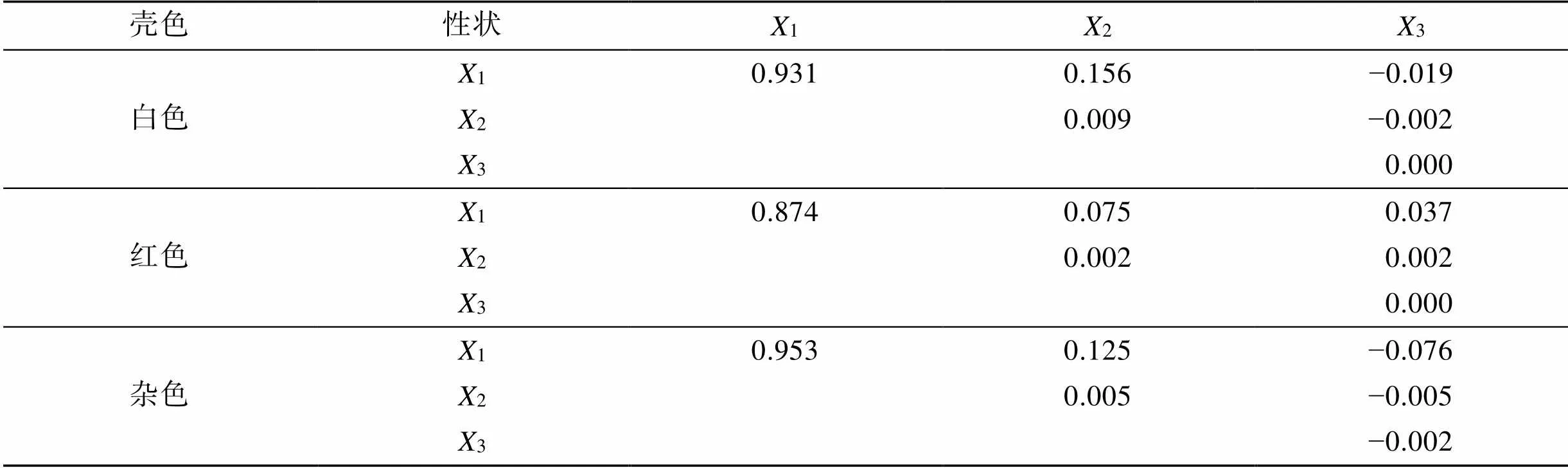
表7 三种壳色硬壳蛤壳形态性状对活体质量的决定系数
2.3 最优回归方程建立
根据通径分析结果, 以壳形态性状为自变量, 活体质量为因变量进行多元逐步回归, 分别建立3种壳色硬壳蛤壳长、壳高、壳宽对活体质量的最优回归方程:
白色= 8.066 + 0.8361+ 0.3132,
红色= 6.806 + 1.1501,
杂色=7.629+0.9791+0.2832.
上述方程对3种壳色硬壳蛤壳形态性状的偏回归系数进行显著性检验见表8。白色和杂色硬壳蛤方程中, 壳长、壳高对活体质量的偏回归系数均达极显著或显著水平(<0.01;<0.05), 而壳宽对活体质量的偏回归系数不显著(>0.05), 应剔除该自变量; 而红色硬壳蛤方程中, 壳长对活体质量的偏回归系数达极显著(<0.01), 而壳高、壳宽对活体质量的偏回归系数不显著(>0.05), 应剔除该自变量。经回归预测, 估算值与实际观察值间差异不显著(>0.05), 说明上述方程能有效反映3种壳色硬壳蛤壳形态性状与活体质量间的真实关系。
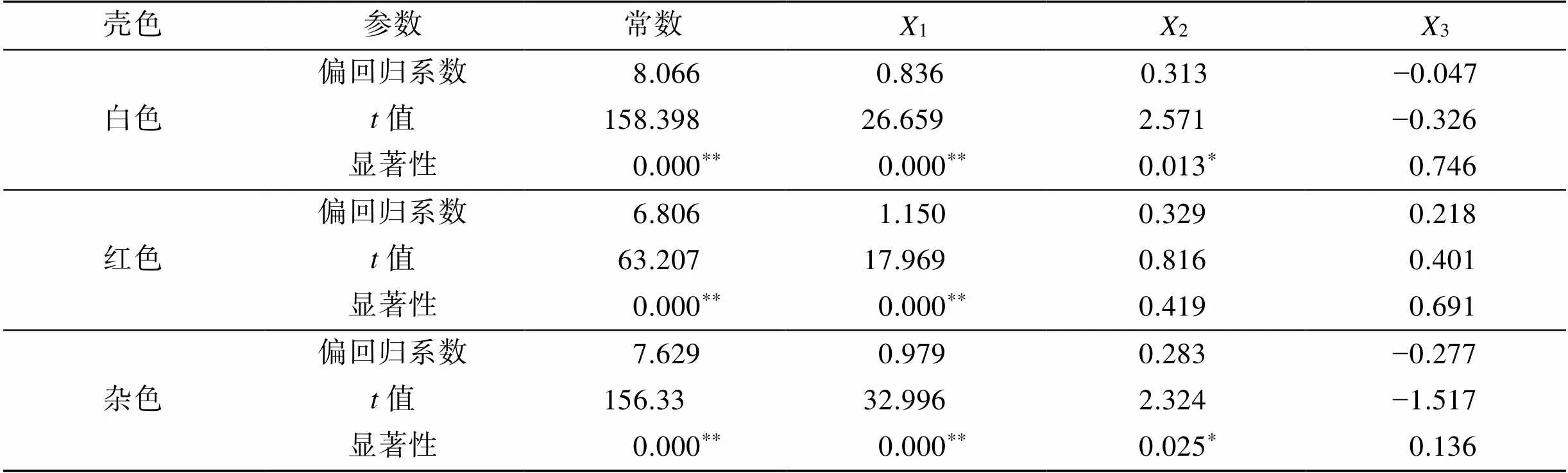
表8 三种壳色硬壳蛤壳形态性状偏回归系数检验
注:*表示差异显著(<0.05);**表示差异极显著(<0.01)
3 讨论
双壳贝类外壳颜色不仅和生态、行为相关[23-24], 还与其生长、存活性状有关[25]。王庆恒等[8]对马氏珠母贝的研究发现, 4个壳色品系幼虫的壳长、壳高存在显著差异。本研究中, 3种壳色群体在幼虫期的生长性状也存在着显著差异(<0.05), 而在稚贝期, 白色群体相较红色和杂色群体则表现出显著的生长优势(<0.05)。已有研究表明, 壳色较浅的双壳贝类更能适应高温养殖环境条件[26], 生长优势更明显。Newkirk[27]对贻贝的研究发现, 高温季节颜色浅的蓝色贝生长率要显著快于颜色深的褐色贝。郑怀平等[28]对海湾扇贝的研究发现, 在稚贝养成时期, 与橙色和紫色家系相比, 白色家系的生长速度更快、存活率更高。赵鹏等[29]对虾夷扇贝的研究发现, 在9月龄时白色贝除壳宽外其余各性状较褐色贝均具有极显著的生长优势。上述研究结果不能解释本研究中3种壳色群体生长性状间差异, 我们认为硬壳蛤对高温的耐受力强, 在室外池塘培育中往往被泥沙所覆盖, 不同壳色群体对光和热的吸收并无差异。壳色作为可稳定遗传的性状, 它与表型性状的联系可能与遗传和环境等因素有关[25]。由于本实验所有壳色群体在相同的养殖环境下培育, 并且选取同一室外养殖池塘的同龄亲贝, 基本可以消除环境因素对生长性状的影响。由此推断, 3种壳色群体在不同时期的生长性状差异可能与色素、遗传因素有关。
在动植物遗传育种中, 通过对各性状间相关性分析可提高选育效果。然而相关性分析不能全面考察两个变量间的真实关系, 使结果具有片面性。杨小刚等[30]对虾夷马粪海胆的研究发现, 其体积与活体质量的相关系数达到极显著水平, 但二者的通径系数却未达到显著水平。通径分析可以将相关系数划分为直接作用和间接影响, 其自变量大小不同对因变量产生的贡献程度也不同[31], 能准确反映出变量间的关系。本研究中, 所测白色、红色和杂色硬壳蛤壳形态性状与活体质量的相关系数均达极显著水平(<0.01), 且呈高度正相关, 这与其他多种贝类中的研究结果相同[32-33]。3种壳色硬壳蛤的壳长、壳高和壳宽性状间存在明显的共线性, 为保证分析结果准确, 我们在相关性分析的基础上进行了主成分分析、通径分析和决定系数的计算。主成分分析结果显示, 3种壳色硬壳蛤的壳长对该主成分影响最大, 壳宽对主成分的影响较小。另外, 在通径分析和计算决定系数时, 当相关指数2或各自变量对因变量的单独决定系数及两两共同决定系数的总和∑大于或等于0.85时, 说明影响因变量的主要自变量已找到[22]。在360日龄时白色、红色和杂色硬壳蛤壳形态性状对活体质量的2和∑均大于0.85, 说明本研究所选壳长、壳高和壳宽是影响活体质量的主要性状, 其他性状影响相对较小, 通径系数分析结果能真实反映壳形态性状与活体质量间的关系。
经逐步回归发现, 本研究选取的壳长、壳高和壳宽性状中, 影响3种壳色硬壳蛤活体质量的主要自变量均相同, 即壳长对活体质量的直接作用最大(0.965, 0.935, 0.947), 且直接作用较间接影响更大; 而壳宽对活体质量的间接影响大于通径系数, 表明壳宽主要通过壳长、壳高产生影响从而间接影响活体质量。宋坚等[34]对硬壳蛤的研究发现, 小规格个体的壳厚、中规格和大规格个体的壳长分别是影响其活体质量的主要因素。本研究结果与上述研究结果存在差异, 究其原因可能与硬壳蛤遗传背景、取样数量以及硬壳蛤日龄不同有关。在不同日龄的同种物种中, 影响其活体质量的主要壳形态性状不一致现象已在许多物种中得到证实, 例如安丽等[35]对中国明对虾的研究发现, 在不同日龄时, 中国明对虾的壳形态性状影响其活体质量的主要因素不同。Huo等[36]报道了不同日龄菲律宾蛤仔影响活体质量的主要壳形态性状也不相同。此外, 杨彦鸿等[37]对南澳青蛤的研究发现, 对活体质量影响最大的主要壳性状为壳长。但闫喜武等[38]对四角蛤蜊的研究发现, 活体质量主要受壳高的影响最大。刘辉等[39]对菲律宾蛤仔的研究发现, 壳宽是影响其活体质量的主要因素。以上研究结果表明, 由于不同双壳贝类的壳形态性状有所差异, 从而导致影响其活体质量的壳形态性状也不同[18]。根据决定系数的计算, 我们发现3种壳色硬壳蛤壳长对活体质量的决定系数最大, 其结果也与主成分分析和通径分析结果相同, 这表明在壳长、壳高和壳宽性状中, 通过壳长指标, 可使3种壳色硬壳蛤的活体质量取得较好的选育效果。
4 结论
本研究在相同养殖环境下调查了3种壳色硬壳蛤群体生长性状间差异, 发现杂色群体在幼虫期较白色和红色群体生长速度快、变态率高, 但存活率低(< 0.05); 在稚贝期, 白色群体则表现出显著的生长优势(<0.05)。此外, 我们查明了白色、红色和杂色硬壳蛤的壳长是影响其活体质量的主要因素, 壳宽为次要因素; 建立了壳长、壳高和壳宽对活体质量的最优回归方程:白色= 8.066+0.8361+0.3132,红色= 6.806+1.1501,杂色= 7.629+0.9791+0.2832。本文研究结果可为3种壳色硬壳蛤良种选育提供重要的理论依据和基础数据。
[1] 张涛, 杨红生, 刘保忠, 等. 环境因子对硬壳蛤稚贝成活率和生长率的影响[J]. 海洋与湖沼, 2003, 34(2): 142-149.Zhang Tao, Yang Hongsheng, Liu Baozhong, et al. Effects of environmental factors on the survival and growth of juvenile hard clam(Linnaeus, 1758)[J]. Oceanologia et Limnologia Sinica, 2003, 34(2): 142-149.
[2] 文海翔. 环境因子对硬壳蛤代谢与生长的影响[D]. 青岛: 中国科学院研究生院(中国科学院海洋研究所), 2004.Wen Haixiang. Effects of environmental factors on metabiolism and growth of hard clam[D]. Qingdao: Graduate University of Chinese Academy of Sciences (Institute of Oceanology, Chinese Academy of Sciences), 2004.
[3] LIU X, WU F C, ZHAO H E, et al. A novel shell color variant of the Pacific abalonesubject to genetic control and dietary influence[J]. Journal of Shellfish Research, 2009, 28(2): 419-424.
[4] KOZMINSKY E V. Inheritance of the background shell color in the snails(Gastropoda, Littorinidae)[J]. Russian Journal of Genetics, 2014, 50(10): 1177-1187.
[5] MITTON J B. Shell color and pattern variation inand its adaptive significance[J]. Chesapeake Science, 1977, 18: 387-390.
[6] ALFONSI C, PEREZ J E. Growth and survival in the scallopas related to self fertilization and shell colour[J]. Boletín del Instituto Oceanográfico de Venezuela, 1998, 37(1/2): 69-73.
[7] 丁鉴锋, 杨霏, 闫喜武, 等. 不同壳色菲律宾蛤仔免疫机能的比较研究[J]. 大连海洋大学学报, 2012, 27(5): 411-416.Ding Jianfeng, Yang Fei, Yan Xiwu, et al.[J]. Journal of Dalian Ocean University, 2012, 27(5): 411-416.
[8] 王庆恒, 邓岳文, 杜晓东, 等. 马氏珠母贝4个壳色选系F1幼虫的生长比较[J]. 中国水产科学, 2008, 15(3): 488-492.Wang Qingheng, Deng Yuewen, Du Xiaodong, et al. Growth comparison of F1larvaefour shell color[J]. Journal of Fishery Sciences of China, 2008, 15(3): 488-492.
[9] 郑怀平, 许飞, 张国范, 等. 海湾扇贝壳色与数量性状之间的关系[J]. 海洋与湖沼, 2008, 39(4): 328-333. Zheng Huaiping, Xu Fei, Zhang Guofan, et al. Relationships between shell colors and quantitative traits in the bay scallop,(Lamarck, 1819)[J]. Ocean and Limnology, 2008, 39(4): 328-333.
[10] NAKAMURA Y, HASHIZUME K, KOYAMA K, et al. Effect of salinity on sand burrowing activity, feeding and growth of the clamsand[J]Journal of Shellfish Research, 2005, 24(4): 1053-1059.
[11] 闫喜武, 张国范, 杨凤, 等. 菲律宾蛤仔莆田群体两个壳色品系生长发育的比较[J]. 大连水产学院学报, 2005, 20(4): 266-269.Yan Xiwu, Zhang Guofan, Yang Feng, et al. A comparison of the growth and development of of manila Clam () from two pedigrees[J]. Journal of Dalian Fisheries University, 2005, 20(4): 266-269.
[12] 张兴志, 张继彪, 刘辉, 等. 菲律宾蛤仔獐子岛群体两种壳型表型性状比较[J]. 水产科学, 2015, 34(10): 616-620. Zhang Xingzhi, Zhang Jibiao, Liu Hui, et al. Comparisons of phenotypic traits in two shell type strains of manila clam () in Zhangzi island[J]. Fisheries Science, 2015, 34(10): 616-620.
[13] 杜美荣, 方建光, 高亚平, 等. 不同贝龄栉孔扇贝数量性状的相关性和通径分析[J]. 水产学报, 2017, 41(4): 580-587.Du Meirong, Fang Jianguang, Gao Yaping, et al. Correlation and path analysis of quantitative traits of different-age[J]. Journal of Fisheries of China, 2017, 41(4): 580-587.
[14] 常亚青, 张存善, 曹学彬, 等. 1龄虾夷扇贝形态性状对重量性状的影响效果分析[J]. 大连水产学院学报, 2008, 23(5): 330-334. Chang Yaqing, Zhang Cunshan, Cao Xuebin, et al. Effect of morphometrical traits on weight traits in one-year old yesso scallop[J]. Journal of Dalian Fisheries University, 2008, 23(5): 330-334.
[15] 刘小林, 常亚青, 相建海, 等. 栉孔扇贝壳尺寸性状对活体重的影响效果分析[J]. 海洋与湖沼, 2002, 33(6): 673-678. Liu Xiaolin, Chang Yaqing, Xiang Jianhai, et al. Analysis of effects of shell size characters on live weight in chinese scallop[J]. Oceanologia et Limnologia Sinica, 2002, 33(6): 673-678.
[16] 孙泽伟, 郑怀平, 杨彦鸿, 等. 近江牡蛎养殖群体数量性状间的相关及通径分析[J]. 中国农学通报, 2010, 26(6): 332-336. Sun Zewei, Zheng Huaiping, Yang Yanhong, et al. Correlation and path analysis to quantitative traits for a cultured population of Jinjiang oyster[J]. Chinese Agricultural Science Bulletin, 2010, 26(6): 332-336.
[17] 张伟杰, 常亚青, 丁君, 等. 日本镜蛤(Reeve)壳尺寸与重量性状的相关与回归分析[J]. 海洋与湖沼, 2013, 44(3): 796-800.Zhang Weijie, Chang Yaqing, Ding Jun, et al. Correlation and regression of shell size and weight ofReeve[J]. Oceanologia et Limnologia Sinica, 2013, 44(3): 796-800.
[18] 方军, 肖国强, 张炯明, 等. 两种壳色文蛤壳形态性状对活体质量的影响[J]. 大连海洋大学学报, 2017, 32(3): 310-315. Fang Jun, Xiao Guoqiang, Zhang Jiongming, et al. Effect of morphological traits on live body weight in clamwith two kind color shells[J]. Journal of Dalian Ocean University, 2017, 32(3): 310-315.
[19] 郭文学, 闫喜武, 马贵范, 等. 两种四角蛤蜊()壳内色品系选育初探[J]. 海洋与湖沼, 2012, 43(2): 262-267. Guo Wenxue, Yan Xiwu, Ma Guifan, et al. A preliminary study on pedigree selection and ofwith two inner-shell colors[J]. Oceanologia et Limnologia Sinica, 2012, 43(2): 262-267.
[20] 巫旗生, 曾志南, 宁岳, 等. 钝缀锦蛤形态性状对活体质量的影响[J]. 水产科学, 2018, 37(1): 110-114. Wu Qisheng, Zeng Zhinan, Ning Yue, et al. Effects of shell morphological traits on live body weight of clam[J]. Fisheries Science, 2018, 37(1): 110-114.
[21] 王诚远, 费春平, 项国仕, 等. 乌龟形态性状对体重的影响效果[J]. 南昌大学学报(理科版), 2013, 37(1): 83-87, 92. Wang Chengyuan, Fei Chunping, Xiang Guoshi, et al. Effects of morphometric traits on body weight for turtle[J]. Journal of Nanchang University(Natural Science), 2013, 37(1): 83-87, 92.
[22] 吴杨平, 陈爱华, 姚国兴, 等. 大竹蛏表型性状通径和回归分析[J]. 南京师大学报(自然科学版), 2012, 35(2): 97-102. Wu Yangping, Chen Aihua, Yao Guoxing, et al. Path and regression analysis of phenotypic characters of[J]. Journal of Nanjing Normal University (Natural Science Edition), 2012, 35(2): 97-102.
[23] SMITH D A S. Polymorphism and selective predation inGmelin (Bivalvia: Tellinacea)[J]. Journal of Experimental Marine Biology & Ecology, 1975, 17(2): 205-219.
[24] BEUKEMA J J, MEEHAN B W. Latitudinal variation in linear growth and other shell characteristics of[J]. Marine Biology, 1985, 90(1): 27-33.
[25] 张跃环, 闫喜武, 张澎, 等. 贝类壳色多态的研究概况及展望[J]. 水产科学, 2008, 27(12): 680-683. Zhang Yuehuan, Yan Xiwu, Zhang Peng, et al. Research advances and prospects on shell color polymorphism of mollusks[J]. Fisheries Science, 2008, 27(12): 680-683.
[26] 孙秀俊, 杨爱国, 刘志鸿, 等. 2种壳色虾夷扇贝的形态学指标比较分[J]. 安徽农业科学, 2002, 36(23): 10008-10010, 10160. Sun Xiujun, Yang Aiguo, Liu Zhihong, et al. Comparative analysis of morphological indices ofwith 2 shell colors[J]. Journal of Anhui Agricultural Sciences, 2002, 36(23): 10008-10010, 10160.
[27] NEWKIRK G F. Genetics of shell color inand the association of growth rate with shell color[J]. Journal of Experimental Marine Biology & Ecology, 1980, 46(1): 89-94.
[28] 郑怀平, 张国范, 刘晓, 等. 不同贝壳颜色海湾扇贝()家系的建立及生长发育研究[J].海洋与湖沼, 2003, 34(6): 632-639. Zheng Huaiping, Zhang Guofan, Liu Xiao, et al. Establishment of different shell color lines of bay scallop(Lamarck, 1819) and their development[J]. Oceanologia et Limnologia Sinica, 2003, 34(6): 632-639.
[29] 赵鹏, 丁君, 常亚青. 两种壳色虾夷扇贝壳体尺性状对活体重影响效果的分析[J]. 大连海洋大学学报, 2011, 26(1): 1-5. Zhao Peng, Ding Jun, Chang Yaqing. Effect of shell size characters on live body weight in Japanese scallopwith two colors[J]. Journal of Dalian Ocean University, 2011, 26(1): 1-5.
[30] 杨小刚, 常亚青, 刘小林. 虾夷马粪海胆壳性状对活体质量的影响效果分析[J]. 水产科学, 2010, 29(2): 83-86. Yang Xiaogang, Chang Yaqing, Liu Xiaolin. Effects of shell size characters on live weight in sea urchin[J]. Fisheries Science, 2010, 29(2): 83-86.
[31] 杨月静, 向梦斌, 刘庭, 等. 13月龄养殖齐口裂腹鱼形态性状与体质量的关系[J]. 河南农业科学, 2019, 48(6): 139-144. Yang Yuejing, Xiang Mengbin, Liu Ting, et al. Relationship between morphological traits and body mass of cultured 13-month-old[J]. Journal of Henan Agricultural Sciences, 2019, 48(6): 139-144.
[32] 高玮玮, 袁媛, 潘宝平, 等. 青蛤()贝壳形态性状对软体部重的影响分析[J]. 海洋与湖沼, 2009, 40(2): 166-169. Gao Weiwei, Yuan Yuan, Pan Baoping, et al. The relationship between shell morphology and body weight of[J]. Oceanologia et Limnologia Sinica, 2009, 40(2): 166-169.
[33] 吴杨平, 陈爱华, 姚国兴, 等. 文蛤贝壳形态性状对活体重的影响分析[J]. 海洋渔业, 2010, 32(3): 320-325. Wu Yangping, Chen Aihua, Yao Guoxing, et al. The relationship between shell morphology and body weight of[J]. Marine Fisheries, 2010, 32(3): 320-325.
[34] 宋坚, 张伟杰, 常亚青, 等. 硬壳蛤形态性状对活体重的影响效果分析[J]. 安徽农业大学学报, 2010, 37(2): 273-277.Song Jian, Zhang Weijie, Chang Yaqing, et al. Effects of shell characters on body weight in hard clam[J]. Journal of Anhui Agricultural University, 2010, 37(2): 273-277.
[35] 安丽, 刘萍, 李健, 等. “黄海1号”中国明对虾形态性状对体质量的影响效果分析[J]. 中国水产科学, 2008, 15(5): 779-786. An Li, Liu Ping, Li Jian, et al. Mathematical analysis of effects of morphometric traits on body weight fornamed “Yellow Sea No. 1”[J]. Journal of Fisheries Science of China, 2008, 15(5): 779-786.
[36] HUO Z M, YAN X W, ZHAO L Q, et al. Effects of shell morphological traits on the weight traits of Manila clam ()[J]. Acta Ecologica Sinica, 2010, 30(5): 251-256.
[37] 杨彦鸿, 李朝霞, 郑怀平, 等. 南澳青蛤()野生群体数量性状间的相关及通径分析[J]. 海洋通报, 2010, 29(5): 550-553. Yang Yanhong, Li Zhaoxia, Zheng Huaiping, et al. Correlation and Path analysis of quantitative traits for natural population ofin Nan’ao Island[J]. Marine Science Bulletin, 2010, 29(5): 550-553.
[38] 闫喜武, 王琰, 郭文学, 等. 四角蛤蜊形态性状对重量性状的影响效果分析[J]. 水产学报, 2011, 35(10): 1513-1518. Yan Xiwu, Wang Yan, Guo Wenxue, et al. Effects of shell morphological traits on the weight traits of clamreeve along northern coast in China[J]. Journal of Fisheries of China, 2011, 35(10): 1513-1518.
[39] 刘辉, 张兴志, 鹿瑶, 等. 菲律宾蛤仔橙色品系壳形态性状对质量性状的通径及多元回归分析[J]. 大连海洋大学学报, 2015, 30(5): 514-518. Liu Hui, Zhang Xingzhi, Lu Yao, et al. Effects of shell morphological traits on the weight traits of orange strain of manila clam[J]. Journal of Dalian Ocean University, 2015, 30(5): 514-518.
Differences in the growth traits and path analysis of the three shell colors of
WANG Yuan-ning1, 2, LI Hua-yu1, 2, MENG Liang1, 2, LI Jin-ze1, 2, DONG Shi-yu1, 2, LI Yong-ren1, GUO Yong-jun1, ZHANG Tao2
(1. Tianjin Key Laboratory of Aqua-ecology and Aquaculture, Fisheries College, Tianjin Agricultural University, Tianjin 300384, China; 2. CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China)
Shell color is an important characteristic affecting the commercial value of shellfish. In this study, populations ofwith different shell colors were used as breeding parent shellfish, and the progeny populations of white, red, and mottled shell colors were successfully bred by random mating between individuals and were cultivated in a pond for 360 days. The shell length (1), shell height (2), shell width (3), and body mass () were measured, and correlation, principal component and path analyses were used to explore the different shell colorsof. Differences in the growth traits between larval and juvenile clams and the effects of shell morphological traits (1,2, and3) on body mass were assessed. The results showed that the mottled population grew faster and had a higher metamorphosis rate at the larval stage than the white and red populations but had a lower survival rate (0.05). The white group had a significant growth advantage at the juvenile stage (0.05). The correlation coefficients between the shell morphological traits (1,2, and3) and body mass () of the three shell colors ofall reached a very significant level (0.01). The results of principal component and path analyses revealed that the shell length (1) ofwas the main factor affecting its body mass (). The results of this study provide an important theoretical basis and basic data for the selection and breeding of different shell colors in.
; shell color; growth traits; path analysis
Sep. 11, 2022
[Tianjin Science and Technology Commission Project, No. 20YFZCSN00240; Tianjin Agricultural Committee Project No. 202103010; The Modern Agricultural Industrial Technology System Fund, No. CARS-49; Shandong Province Agricultural Major Applied Technology Innovation Project, No. SF1405303301]
S968.31
A
1000-3096(2023)8-0023-10
10.11759/hykx20220911001
2022-09-11;
2022-10-13
天津市科委项目(20YFZCSN00240); 天津市农委项目(202103010); 现代农业产业技术体系专项资金资助(CARS-49); 山东省农业重大应用技术创新项目(SF1405303301)
王元宁(1996—), 男, 河北唐山人, 硕士研究生, 主要从事贝类遗传育种研究, E-mail: ww123n@163.com; 张涛(1971—), 通信作者, 男, 山东青岛人, 研究员, 博士生导师, 主要从事贝类苗种繁育和增养殖、海洋牧场构建原理与技术研究, E-mail: tzhang@gdio.ac.cn
(本文编辑: 赵卫红)
