朱砂根成熟果实不同部位生理特性与种子胎生的关系
2023-11-20赵财宝王斌潘曲波王锦艾星梅孙媛媛
赵财宝,王斌,潘曲波,王锦,艾星梅*,孙媛媛
(1.西南林业大学 园林园艺学院,国家林业和草原局西南风景园林工程中心,云南 昆明 650224;2.云南省农业科学院生物技术与种质资源研究所,云南 昆明 650205)
朱砂根(Ardisia crenata)隶属于报春花科(Primulaceae)紫金牛属(Ardisia)常绿灌木,广泛分布于热带、亚热带地区温暖潮湿的山地林下。成熟果实为红色球形浆果,少数为黄果或白果[1],具有很高的观赏价值,果实密度大且挂果期长,能在母株上附着一年或更长时间,存在花果同期的现象,且耐阴性较强,可作为室内观赏和城市景观的理想树种。目前,对朱砂根的研究主要集中在资源调查[2]、逆境胁迫生理[3-4]、形态发育及分子鉴定[5-7]、居群遗传多样性分析[8]以及化学成分和药理作用[9]等方面。研究发现,朱砂根植株的根、茎、叶中含有香豆素类、皂苷类等多种生物活性化合物,具有显著的抗肿瘤、抗癌、抗炎特性[10-11],对病毒的转移具有重要的抑制作用[12]。
近年来,由于朱砂根具有较高的药用价值和观赏价值而被大量开发,导致野生资源栖息地碎片化,自然分布范围逐渐缩小,面临着灭绝的高风险[13],因此,应不断提高其繁育系数,朱砂根通常采用种子、扦插和种胚离体繁殖[14],尽管种子较多,但通过种子直播成苗较慢,优质扦插苗的成活率较低[15]。由于挂果期长,种子常表现出胎生现象,这可能与它们生活的温暖潮湿的林下弱光环境有关[16],胎萌后脱离母体可快速成苗,是繁殖方式的一种重要补充。前期研究发现,朱砂根成熟种子有些胎生,有些非胎生,除了受环境因子影响以外,还可能与自身的遗传特性有关,目前尚未见相关报道。因此,以朱砂根胎生和非胎生成熟果实为材料,分别剥离外果皮、中果皮、内果皮、胚乳及胎生种子胚根,测定不同部位的碳水化合物及酶活性变化,探讨其生理特征差异性与胎生的关系,为进一步研究朱砂根种子的胎生机理提供理论依据。
1 材料与方法
1.1 实验材料
本研究所用的材料均为5~6年野生朱砂根植株,结合前期研究基础,根据 Chen 和Pipoly 在《Flora of China》中的描述[17],对朱砂根植株进行了鉴定分析。移植于西南林业大学苗圃基地(25°03′ N,102°46′ E,亚热带高原季风气候,年平均温度12.5℃,年平均降水量约900 mm)。2020年11月中旬,从10株生长良好且性状稳定的植株上分别采集新鲜成熟的胎生与非胎生果实各200粒,果实饱满均匀无病虫害,立即带回实验室备用。
1.2 方法
用自来水将果实清洗干净后迅速擦干水分,分别剥离胎生与非胎生朱砂根果实的外果皮、中果皮、内果皮、胚乳及胎生种子胚根,将样品用液氮速冻后放入-80℃的冰箱中冷冻保存,分别测定不同部位的生理指标差异性。可溶性蛋白含量采用考马斯亮蓝法测定;可溶性糖、淀粉含量采用蒽酮比色法测定;过氧化物酶(POD)活性采用愈创木酚法测定,超氧化物氧化酶(SOD)采用氮蓝四唑法测定,α-淀粉酶和β-淀粉酶活性采用3,5-二硝基水杨酸法测定,以上方法均采用但方[18]的方法,重复3次,取平均值。
1.3 数据分析
采用Microsoft Excel 2013对结果进行数据统计,同时利用SPSS Statistics 23.0进行单因素方差分析(ANOVA)及Duncan多重比较,并进行相关性分析,计算样本平均值之间差异的显著性(P< 0.05)。
2 结果与分析
2.1 朱砂根胎生与非胎生果实可溶性蛋白含量差异
朱砂根胎生与非胎生成熟果实不同部位的蛋白质含量变化如图1所示,从不同部位来看,无论是胎生还是非胎生,同一果实中内果皮的蛋白质含量均显著高于其他部位(P <0.05),表现为内果皮>胚乳>外果皮>中果皮,胎生后胚根中的蛋白质含量也较高,仅次于胚乳;从胎生和非胎生果实来看,朱砂根非胎生果实外果皮、中果皮和内果皮的蛋白质含量均显著高于胎生果实(P< 0.05),而胚乳中的蛋白质含量均无显著差异性(P >0.05),说明朱砂根的胎生可能与果皮,尤其是中果皮的蛋白质含量下降有关,而受胚乳的影响较小。
2.2 朱砂根果实可溶性糖和淀粉含量差异
朱砂根胎生与非胎生果实不同部位的可溶性糖和淀粉含量变化如图2所示,可以看出,种子胎生后的胚根可溶性糖含量最高(图2a),为8.79 mg·g-1,显著高于其他部位(P <0.05);胎生和非胎生果实由外向内不同部位的可溶性糖含量均呈先升后降的趋势,表现为中果皮>内果皮>外果皮>胚乳;胎生后胚乳中的可溶性糖含量有一定的积累,但与非胎生果实相比,差异不显著(P >0.05),除此之外,胎生果实各部位的可溶性糖含量均低于非胎生果实,说明种子胎生可能与果皮中的可溶性糖含量下降有关。对两种果实不同部位的淀粉含量进行比较,结果如图2b所示,朱砂根非胎生果实内果皮的淀粉含量最高,为6.18%,其次为胚乳,均极显著高于外果皮和中果皮(P< 0.01),胎萌发生后,胚根中的淀粉含量最高,均显著高于其他部位(P< 0.05),而外果皮和中果皮的淀粉含量变化不显著(P >0.05),内果皮和胚乳中的淀粉含量则迅速下降,说明胚根中淀粉的积累可能主要来源于内果皮和胚乳。
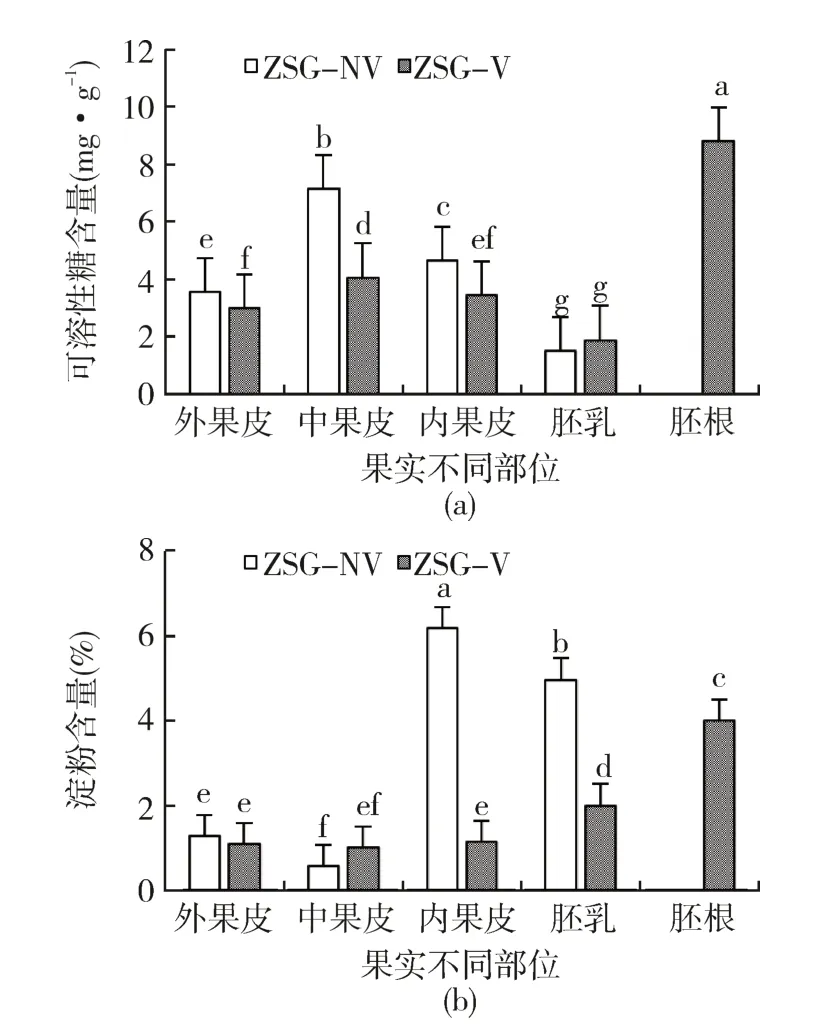
图2 朱砂根胎生与非胎生果实可溶性糖和淀粉含量变化Fig.2 Changes of soluble sugar and starch contents in viviparous and non-viviparous fruit of A. crenata
2.3 朱砂根胎生与非胎生果实抗氧化酶活性差异
从图3可以看出,朱砂根胎生与非胎生果实不同部位的抗氧化酶活性存在显著差异性(P<0.05),非胎生果实从外向内不同部位的SOD酶活性先降低后升高,其中以胚乳中的SOD活性最高,而胎生果实外果皮的SOD酶活性最高,为918.08 U·g-1,依次向内逐渐降低,以胚根中的SOD活性最低,仅为9.61 U·g-1(图3a),胎生与非胎生各部位间的SOD活性均差异显著(P <0.05);对于POD酶活性(图3b),从不同部位来看,果实从外向内的POD酶活性均呈先升后降的趋势,其中以胎生果实中果皮的POD酶活性最高,其次为内果皮,胚根中的POD酶活性最低;从胎生与非胎生果实来看,除两者的外果皮间无显著差异性以外(P >0.05),胎生果实的POD活性较非胎生果实高且差异显著(P <0.05),说明胎萌发生后,胚根突破种皮,果实各部位均启动了抗氧化酶系统来抵御外界不良环境。
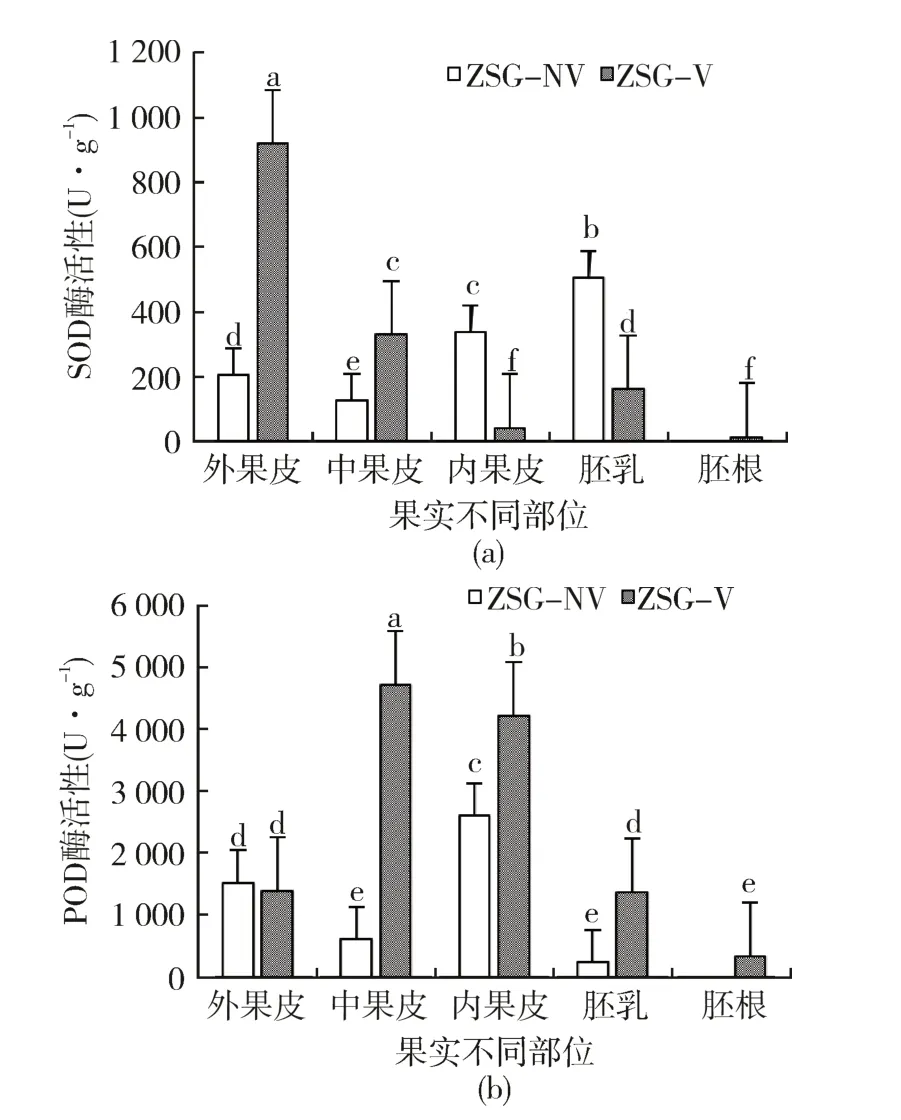
图3 朱砂根胎生与非胎生果实SOD和POD酶活性变化Fig.3 Changes of SOD and POD activities in viviparous and non-viviparous fruit of A. crenata
2.4 朱砂根胎生与非胎生果实淀粉酶活性差异
朱砂根胎生与非胎生成熟果实不同部位的淀粉酶活性变化如表1所示,可以看出,无论是胎生还是非胎生果实,不同部位的淀粉酶活性均存在较大的差异性,其中以内果皮的α-淀粉酶和β-淀粉酶活性最高,均显著高于同一果实其它部位(P< 0.05),表现为内果皮>外果皮>中果皮>胚乳;相比较两种淀粉酶活性,果实不同部位中的β-淀粉酶远高于α-淀粉酶,且非胎生果实均高于胎生果实;从α+β淀粉酶活性来看,胎萌发生后,胚根中的淀粉酶活性较高,而胚乳中的淀粉酶活性虽略有增加,但差异不显著(P> 0.05),而其它部位的α+β淀粉酶活性均显著下降(P< 0.05),说明淀粉酶在朱砂根种子胎萌过程中主要对果皮中的淀粉发生了水解作用。
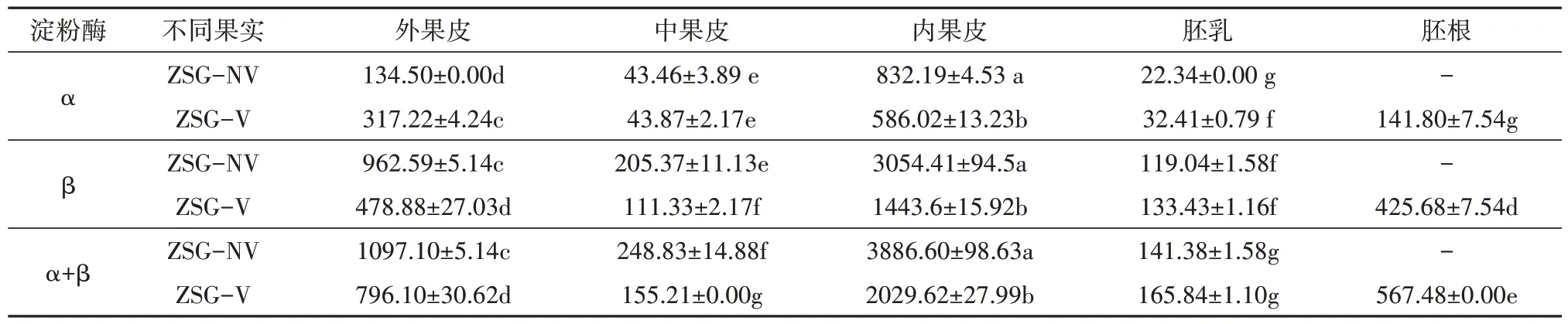
表1 朱砂根胎生与非胎生果实淀粉酶活性变化Tab.1 Changes of amylase activity in viviparous and non-viviparous of A. crenata(单位:µg·g-1·min-1)
2.5 朱砂根胎生与非胎生果实生理特性相关性分析
朱砂根胎生和非胎生果实碳水化合物含量及酶活性相关性分析如表2所示,可以看出,无论是胎生还是非胎生,果实中的蛋白质含量与α-淀粉酶和β-淀粉酶之间均呈显著或极显著正相关(P <0.05),对于非胎生果实而言,淀粉含量与蛋白质、SOD、淀粉酶活性之间呈显著或极显著正相关(P <0.05),可溶性糖含量与SOD呈极显著负相关(P <0.01);对于胎生果实而言,淀粉与可溶性糖含量呈极显著正相关(P <0.01),而与POD活性呈极显著负相关(P <0.01)。
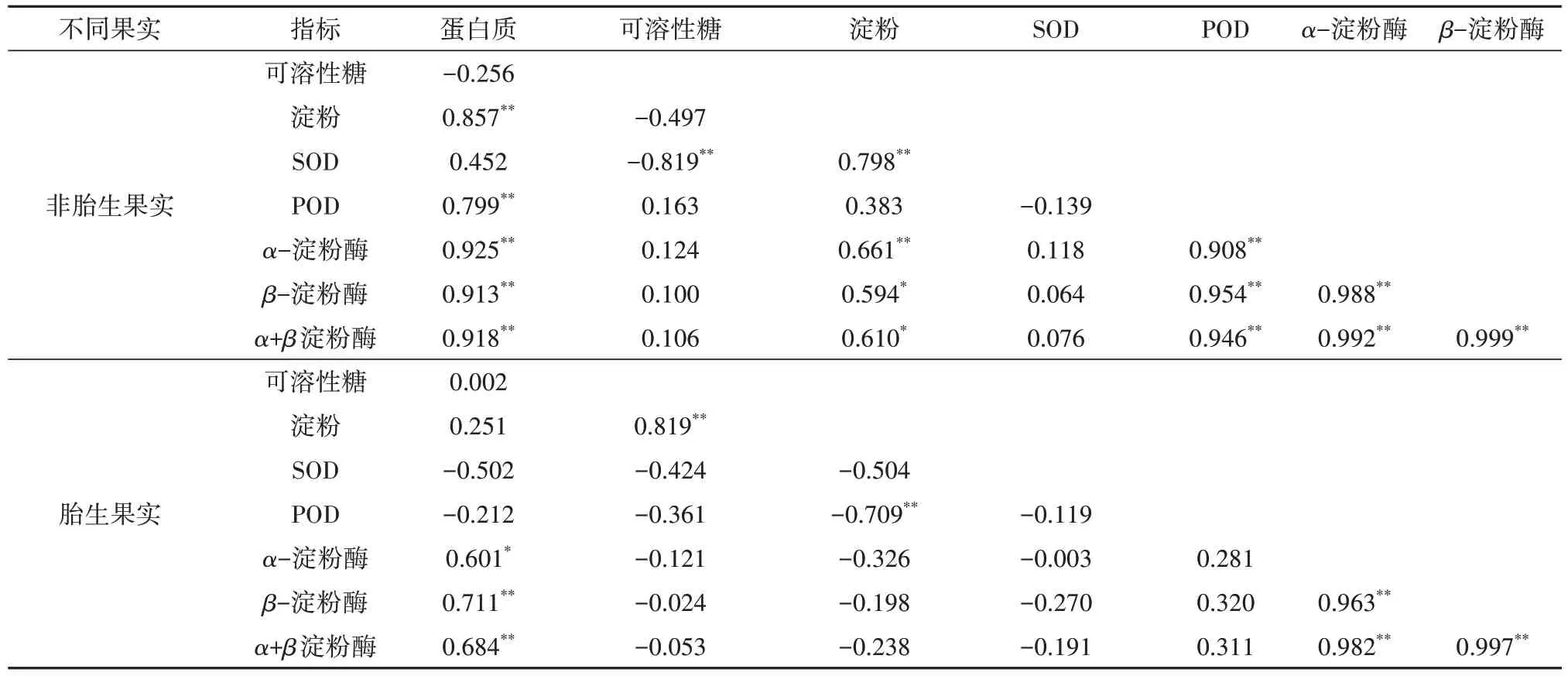
表2 胎生与非胎生果实碳水化合物及酶活性相关性分析Tab.2 Correlation analysis of carbohydrates content and enzymes activities in viviparous and non-viviparous fruits
3 结论与讨论
种子胎萌是自然界的一种常见现象,与动物胎生现象相仿,种子离体前直接在植株上发芽,脱落后可快速成苗,受自身遗传因素或外界高温、高湿、降雨等特殊的环境条件的诱导,以使植物适应更多的生长环境,从而提高其竞争力[19]。近年来国内外学者越来越关注植物胎萌机理的研究,其中在遗传上以胎萌为主要繁殖方式的红树科植物最为常见[20]。对于一些特殊植物来说,种子胎萌具有无可比拟的优势,仙人掌科植物[21-22]、珠芽蓼[23]等兼具胎生繁殖特性,通过适应海拔、温度、光照的变化来改变有性和无性繁殖器官形成的比例[24],甚至植株的高度和胎萌的数量。研究表明,胎萌种子的发芽活力、抗逆性等均高于非胎萌种子[25-26],是后代为了避免和适应不良生存环境的一种表现[27-28],且优良性状能遗传给后代群体[29]。
生长于热带、亚热带的某些顽拗性种子同时具有胎萌现象[22],从某种意义上来说也是植物对周围环境适应的结果[30],顽拗性种子生理代谢活跃,且种皮薄,可增强与环境的气体交换[31],但不会阻止胚胎轴的伸长和发芽,一旦发生会导致最终的水分损失[32]。本研究中,成熟后的朱砂根种子与许多热带物种如枹栎[33]、紫荆木属植物[34]的顽拗性种子一样,对脱水敏感,成熟采收时含水量较高,不能耐受成熟脱水,也没有明显的休眠特性,而种子胎萌反映了植株对不同环境的适应策略,是植物繁殖途径的一种重要补充。
种皮具有保护种胚和萌发时吸水和透气的作用,其性质和结构随植物种类而异[35],在长期的环境适应过程中,果种皮的相关生物学特性也会影响种子萌发及其生理特征[36]。研究表明,种子内的碳水化合物可为胎生器官的萌发提供所需能量,其代谢水平的变化与种子萌发密切相关,米勒魔芋珠芽萌发以后蛋白质含量逐渐下降[37];水稻籽粒中可溶性糖和淀粉含量越高越易发生胎萌[38];黄瓜种子胎萌后的可溶性糖含量逐渐降低,说明种子胎萌需要消耗大量的可溶性糖[39-40],而胎生后的桐花树种子淀粉含量不断积累,可能是由于可溶性糖转变成了贮存物质淀粉的结果[41],这与本研究的结果类似,朱砂根成熟胎生果实不同部位的蛋白质、可溶性糖和淀粉含量均显著低于非胎生果实,而胚根中的含量均较高,说明胎萌发生后果实不同部位的碳水化合物可能向胚部进行了转移。此外,抗氧化酶、α-淀粉酶等的活性变化也是影响种子萌发的重要因素。通常情况下,植物体内自由基的产生和消除之间处于动态平衡,各种酶活性均维持在一定水平。研究表明,易胎萌春大豆种子中的过氧化物酶活性比不易胎萌的种子高[42],而高枝假木贼种子胎萌也与过氧化物酶、超氧化物歧化酶和过氧化氢酶等活性在萌发不同阶段的协同作用有关,为种子在逆境中胎萌并正常生长提供保护[43]。本研究中,非胎生果实淀粉含量与SOD酶活性呈极显著正相关,但与POD酶活性差异不显著,而胎生果实中的SOD和POD酶活性大多高于非胎生果实,且淀粉含量与SOD呈负相关,而与POD呈极显著负相关,说明种子胎萌发生后,各部位的淀粉含量均逐渐降低,启动了相应的抗氧化酶防御系统。此外,种子中淀粉的分解需要一系列酶的催化与调控,较高的α、β-淀粉酶活性,有助于水稻种子内淀粉的分解[38],易于在穗上发芽的水稻种子在成熟后期α-淀粉酶活性明显高于不易在穗上发芽的种子[44],表明α-淀粉酶可能对后期胚的发育具有一定的调节作用[45]。本研究中,无论是胎生还是非胎生果实,内果皮中的α-淀粉酶和β-淀粉酶活性均显著高于同一果实其它部位,且非胎生果实高于胎生果实,说明胎萌发生后,淀粉酶活性迅速降低为胚胎提供能量。
朱砂根花芽分化的顺序为从下往上、自外向内[6],成熟果实均会发生不同程度的胎萌,但主要集中在植株中下部,这可能与挂果期较长、营养物质的优先分配和积累有关。与非胎生果实相比,胎生果实不同部位的碳水化合物可能协同参与胚发育的代谢过程,其含量被降解用作胚的组织分化发育所需的能量,并在胚内积累,尤其是内果皮中的碳水化合物、淀粉酶与POD酶活性均较高,可能对朱砂根的胎萌起重要的调控作用,需进一步从植物激素调控水平以及胎萌相关基因表达等方面深入研究,全面探讨胎生发育的遗传机制。
