内质网应激在IL-1β 诱导软骨细胞炎性损伤中的作用机制
2023-11-18符来想陈前永
夏 克,符来想,陈前永
骨性关节炎(osteoarthritis,OA)是常见的骨关节退行性疾病,以关节软骨变性、损伤、破坏及代偿性软骨下骨质增生、骨赘形成为特征。 软骨细胞是关节软骨组织中唯一的细胞类型, 参与软骨组织功能及形态、细胞外基质代谢的调控。 软骨细胞炎症性损伤与OA 的发生发展密切相关[1,2],但相关的分子机制尚不清楚,临床上也缺乏特异性的OA 治疗药物。 近些年的分子生物学研究证实内质网应激(endoplasmic reticulum stress,ERS)参与多种慢性疾病的发生发展;ERS 是指内质网稳态失去平衡后出现内质网中错误折叠蛋白聚集的病理状态, 持续激活的ERS 是细胞凋亡的上游信号、 介导促进细胞凋亡的生物学效应。软骨细胞凋亡是造成OA 进程中软骨细胞损伤的重要生物学因素, 相关细胞实验证实ERS 激动剂促进软骨细胞凋亡、 相关动物实验证实ERS 拮抗剂改善OA 大鼠的关节软骨退行性变及细胞凋亡[3,4]。 基于此提出假说:ERS 调控软骨细胞凋亡参与OA 进程中软骨骨细胞炎症性损伤。 为验证该假说,笔者将在白细胞介素1β(interleukin-1β,IL-1β)诱导软骨细胞炎症性损伤模型中观察ERS 对细胞凋亡的调控作用,旨在为初步揭示OA 发病机制、发现OA 新的治疗靶点提供依据。
1 材料与方法
1.1 实验材料
1.1.1 细胞株
小鼠软骨细胞株ATDC5, 由中国武汉尚恩生物技术有限公司提供。
1.1.2 主要试剂与仪器
IL-1β( Sigma,美国);c-Jun 氨基末端激酶(c-Jun N-terminal kinase,JNK) 拮抗剂SP600125(MCE,美国); 含半胱氨酸的天冬氨酸蛋白水解酶-12(cysteinyl aspartate specific proteinase 12,Caspase-12)抑制剂氟甲基酮(Z-ATAD-fluoromethylketone,Z-ATADFMK)、CCK8 法增殖检测试剂盒(Abcam,美国);TdT介导的dUTP 缺口末端标记(TdT-mediated dUTP nick end labeling,TUNEL)凋亡检测试剂盒(荧光法)(上海碧云天生物公司, 中国); 葡萄糖调节蛋白78 (glucose-regulated protein 78,GRP78)、C/EBP 同源蛋白(C/EBP homologous protein,CHOP)、 磷酸化c-Jun 氨基末端激酶(phosphorylation c-Jun N-terminal kinase,p-JNK)、裂解型caspase-12(Cleaved caspase-12)、βactin 特异性一抗(CST,美国)。
1.2 方法
1.2.1 细胞培养及分组处理
将ATDC5 细胞复苏后,在含有10%胎牛血清的培养液中贴壁培养,定期更换培养液、用0.25%胰蛋白酶进行消化传代。分为对照组、IL-1β 组、si-NC 组、si-NC+IL-1β 组、si-CHOP+IL-1β 组、溶剂对照组、溶剂+IL-1β 组、JNK 拮抗剂+IL-1β 组、Caspase-12抑制剂+IL-1β 组。
传代后的细胞接种在培养板内进行分组处理。对照组用不含药物的培养液处理,IL-1β 组含有10 ng/mL IL-1β 的培养液处理,si-NC 组转染NC siRNA,si-NC+IL-1β 组在含有10 ng/mL IL-1β 的培养液中转染NC siRNA,si-CHOP + IL-1β 组在含有10 ng/mL IL-1β的培养液中转染CHOP siRNA,溶剂对照组用含有二甲基亚砜(dimethylsulfoxide,DMSO)(体积分数0.1%)的培养液处理,溶剂+IL-1β 组用含有DMSO(体积分数0.1%) 和10 ng/mL IL-1β 的培养液处理,JNK拮抗剂+IL-1β 组用含有10.0 μmol/L(DMSO 的体积分数为0.1%)JNK 拮抗剂SP600125 和10 ng/mL IL-1β 的培养液处理,Caspase-12 抑制剂+IL-1β 组用含有2.0 μmol/L(DMSO 的体积分数为0.1 %)Caspase-12 抑制剂Z-ATAD-FMK 和10 ng/mL IL-1β 的培养液处理。
1.2.2 细胞增殖的检测
将5×103个ATDC5 细胞接种96 孔培养板内,贴壁24 h 后按照1.2.1 节进行分组处理; 继续培养24 h后,在每个培养孔内加入10 μL CCK8 溶液,继续培养1.5 h 后将培养板放入酶标仪, 检测每孔细胞在波长490 nm 的光密度(optical density,OD)490nm值。
1.2.3 细胞凋亡的检测
将1×104个ATDC5 细胞接种24 孔培养板内,贴壁24 h 后按照1.2.1 节进行分组处理; 继续培养24 h 后弃去培养液, 用4%多聚甲醛溶液固定细胞后,采用试剂盒进行TUNEL 染色和4",6- 二脒基-2-苯基吲哚 (4",6-diamidino-2-phenylindole,DAPI)复染,抗荧光猝灭封片液封片后在显微镜高倍(×400)视野下观察。凋亡细胞显绿色荧光,细胞核显蓝色荧光。计算凋亡率(%)=凋亡细胞数/细胞总数×100%。
1.2.4 蛋白表达的检测
将5×105个ATDC5 细胞接种12 孔培养板内,贴壁24 h 后按照1.2.1 节进行分组处理;继续培养24 h后弃去培养液、提取细胞蛋白,将含有20 μg 蛋白的样本加入聚丙烯酰胺凝胶中电泳,而后将凝胶中的蛋白电转移至硝酸纤维素膜, 将膜放入5%脱脂牛奶中、 摇床上室温孵育1 h, 洗膜3 次后分别孵育GRP78(1 ∶500 稀释)、CHOP(1 ∶600 稀释)、p-JNK(1 ∶400 稀释)、Caspase-12(1 ∶800 稀释)及β-actin(1 ∶5 000 稀释)的特异性一抗、4 ℃过夜。 次日洗膜3次后室温孵育辣根过氧化物酶标记的二抗(1 ∶4 000稀释)1 h,再次洗膜3 次,在凝胶成像系统中对膜上的蛋白进行显影,以目的蛋白/β-actin 的灰度值分析蛋白表达水平。
1.3 统计学方法
采用SPSS 23.0 版本软件对实验数据进行统计学处理。 计量资料以均数±标准差表示,两组间比较采用独立样本t检验, 多组间比较采用单因素方差分析,两两比较采用LSD-t法。P<0.05 为差异有统计学意义,P<0.01 为差异有显著统计学意义。
2 结果
2.1 IL-1β 组与对照组细胞增殖和凋亡水平比较
采用CCK8 法检测细胞增殖的OD490nm水平,采用TUNEL 法检测细胞的凋亡率。IL-1β 组ATDC5 细胞的OD490nm水平低于对照组 (0.51 ± 0.07vs1.06 ±0.12), 细胞凋亡率高于对照组 (9.58%±1.24%vs1.35%±0.17%),差异有显著统计学意义(t=8.853、14.703,P<0.001)。
2.2 IL-1β 组与对照组ERS 标志蛋白和凋亡蛋白表达水平比较
采用Western blot 检测ERS 标志蛋白GRP78 和ERS 凋亡蛋白CHOP、p-JNK、Cleaved caspase-12 的表达水平。IL-1β 组ATDC5 细胞中GRP78、CHOP、p-JNK、Cleaved caspase-12 的相对表达水平高于对照组, 差异有显著统计学意义 (t= 9.339、17.206、11.667、10.217,P<0.001)。 见图1、表1。
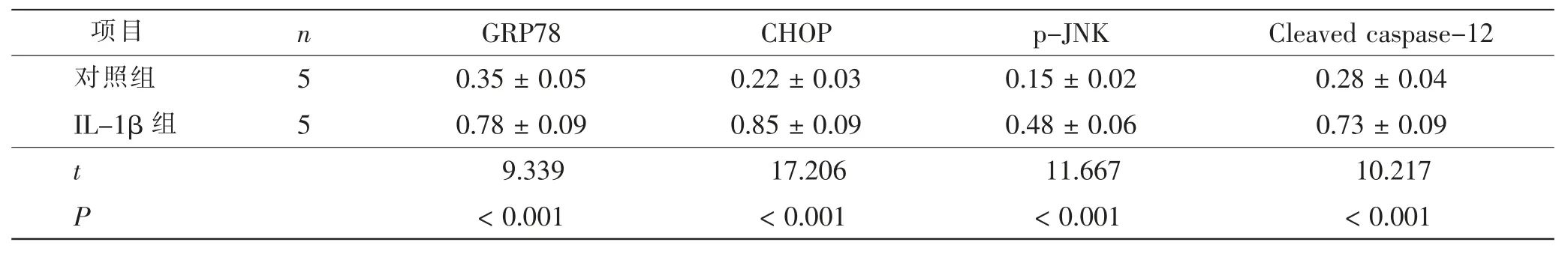
表1 对照组与IL-1β 组ATDC5 细胞GRP78、CHOP、p-JNK、Cleaved caspase-12 表达比较Tab.1 Comparison of GRP78,CHOP,p-JNK,Cleaved caspase-12 expression in ATDC5 cells between control group and IL-1β group

图1 对照组和IL-1β 组ATDC5 细胞中GRP78、CHOP、p-JNK、Cleaved caspase-12 的表达电泳图Fig. 1 Electrophoretograms of GRP78, CHOP, p-JNK and Cleaved caspase-12 expression in ATDC5 cells of control group and IL-1β group
2.3 si-NC 组、si-NC + IL-1β 组、si-CHOP +IL-1β 组组间细胞增殖和凋亡水平比较
si-NC+IL-1β 组ATDC5 细胞增殖OD490nm水平低于si-NC 组,细胞凋亡率及CHOP 表达水平高于si-NC 组(t=14.378、10.001、14.536,P<0.05);si-CHOP+IL-1β 组ATDC5 细胞OD490nm水平高于si-NC+IL-1β 组,凋亡率及CHOP 表达水平低于si-NC+IL-1β组(t=16.250、9.395、11.757,P<0.05)。 见图2、表2。

图2 si-NC 组、si-NC+IL-1β 组、si-CHOP+IL-1β 组CHOP表达电泳图Fig. 2 Electrophoretograms of CHOP expression in si-NC group,si-NC+IL-1β group and si-CHOP+IL-1β group
2.4 溶剂对照组、溶剂+ IL-1β 组、JNK 拮抗剂+IL-1β 组组间软骨细胞增殖和凋亡水平比较
溶剂+ IL-1β 组ATDC5 细胞增殖OD490nm水平低于溶剂对照组, 细胞凋亡率及p-JNK 的表达水平高于溶剂对照组(t=12.964、9.833、15.408,P<0.05);JNK 拮抗剂+ IL-1β 组ATDC5 细胞增殖OD490nm水平高于溶剂+IL-1β 组, 细胞凋亡率及p-JNK 的表达水平低于溶剂+ IL-1β 组 (t= 14.795、9.779、11.339,P<0.05)。 见图3、表3。
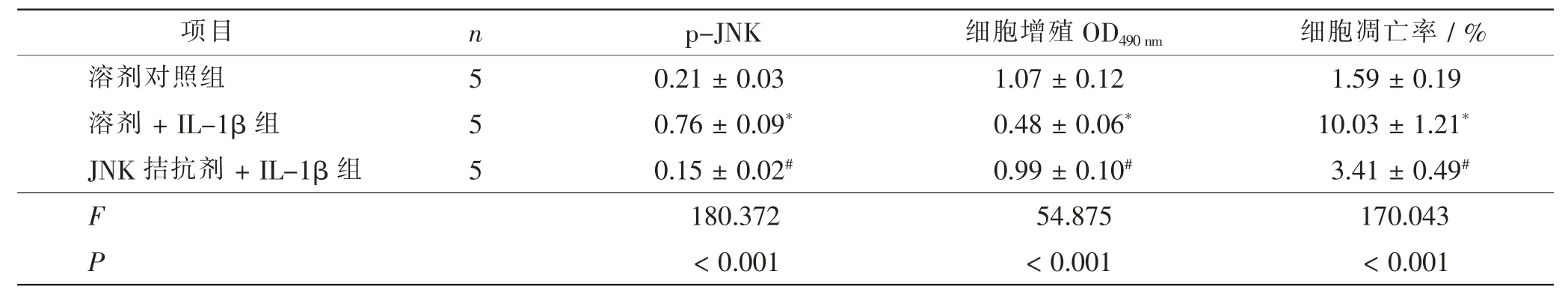
表3 溶剂对照组、溶剂+IL-1β 组、JNK 拮抗剂+IL-1β 组组间p-JNK 表达、增殖和凋亡比较Tab.3 Comparison of p-JNK expression,proliferation and apoptosis in solvent control group,solvent+IL-1β group and JNK antagonist+IL-1β group
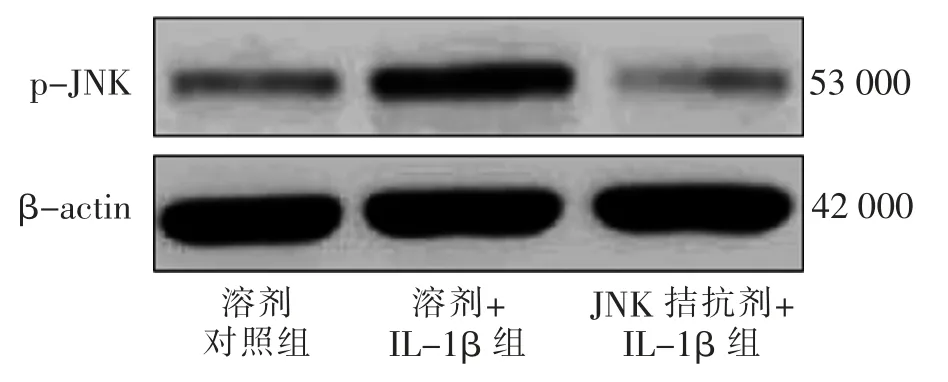
图3 溶剂对照组、 溶剂+IL-1β 组、JNK 拮抗剂+IL-1β 组p-JNK 表达电泳图Fig.3 Electrophoretograms of p-JNK expression in solvent control group,solvent+IL-1β group and JNK antagonist+IL-1β group
2.5 溶剂对照组、 溶剂+ IL-1β 组、Caspase-12 抑制剂+ IL-1β 组组间细胞增殖和凋亡水平比较
溶剂+ IL-1β 组ATDC5 细胞增殖OD490nm水平低于溶剂对照组, 细胞凋亡率及Cleaved caspase-12表达水平高于溶剂对照组 (t= 11.600、11.289、15.912,P<0.05);Caspase-12 抑制剂+IL-1β 组ATDC5 细胞增殖OD490nm水平高于溶剂+IL-1β 组,细胞凋亡率及Cleaved caspase-12 表达水平低于溶剂+IL-1β 组(t=13.080、9.117、11.658,P<0.05)。见图4、表4。

表4 溶剂对照组、溶剂+IL-1β 组、Caspase-12 抑制剂+IL-1β 组组间Cleaved caspase-12 表达、增殖和凋亡比较Tab. 4 Comparison of Cleaved Caspase-12 expression, proliferation and apoptosis in solvent control group, solvent + IL-1β group and Caspase-12 inhibitor+IL-1β group

图4 溶剂对照组、 溶剂+IL-1β 组、Caspase-12 抑制剂+IL-1β 组Cleaved caspase-12 表达电泳图Fig. 4 Electrophoretograms of Cleaved caspase-12 expression in solvent control group, solvent + IL-1β group and Caspase-12 inhibitor+IL-1β group
3 讨论
关节软骨细胞的炎性损伤在OA 的进程中发挥重要作用,在炎性介质的作用下软骨细胞发生过度凋亡会导致软骨组织退行性变和破坏、 软骨下骨赘形成。 临床上常用的OA 治疗药物包括非甾体类药物、软骨营养和修复药物, 但受限于OA 发病机制不明确,目前缺乏能够抑制软骨细胞凋亡的特异性OA 治疗药物[5,6]。多项国内外OA 相关的细胞实验研究采用10 ng/mL IL-1β 诱导软骨细胞炎性损伤的方式建立OA 细胞模型[7~9],笔者也采用该方法建立OA 细胞模型, 细胞模型增殖水平降低、 凋亡率增加的变化与OA 进程中软骨细胞凋亡的特征符合。 笔者将在该OA 细胞模型中研究软骨细胞凋亡的调控机制。
细胞凋亡的调控途径包括线粒体凋亡途径、死亡受体凋亡途径、ERS 凋亡途径。 ERS 是新近受到越来越多关注的凋亡调控机制[10,11]。 内质网在细胞内负责蛋白质合成转运、糖基化修饰、钙离子再分布的调控;慢性炎症、缺血缺氧、高糖等病理因素刺激会造成未折叠蛋白或错误折叠蛋白在内质网中聚集,进而激活ERS。 适度的ERS 有助于维持细胞内蛋白质稳定、保证细胞的适应性存活; 但是持久且强烈的ERS 不利于细胞内环境稳定, 会造成GRP78 表达增加并通过下游信号转导促进细胞凋亡、加重组织损伤[12~14]。 既往OA 相关的动物实验和细胞实验[4,15,16]、笔者研究的细胞实验均证实OA 软骨组织及软骨细胞中内质网标志基因GRP78 的表达水平均明显增加,OA 进程中存在ERS 的过度激活。 已有研究证实ERS 激动剂对软骨细胞的凋亡具有促进作用,ERS 拮抗剂抑制OA大鼠关节软骨的细胞凋亡[3,4],提示ERS 可能通过促进软骨细胞凋亡的途径参与OA 的发生发展,但ERS调控软骨细胞凋亡的具体机制尚不明确。
ERS 对细胞凋亡的途径依赖于下游CHOP、JNK、Caspase-12 途径,CHOP 通过调控下游B 细胞淋巴瘤/白血病-2 削弱细胞的抗凋亡能力、促进细胞色素C 的释放及细胞凋亡;JNK 磷酸化为有活性的p-JNK及Caspase-12 裂解为有活性的Cleaved caspase-12均能作用于Caspase 级联激活途径, 引起Caspase-3激活并引起细胞凋亡[17,18]。 笔者研究在OA 软骨细胞模型中检测到CHOP、p-JNK、Cleaved caspase-12 的表达水平均增加, 提示OA 进程中ERS 的激活可能引起下游CHOP 的高表达、JNK 的磷酸化活化、Caspase-12 的裂解后活化,进而介导了促进细胞凋亡的生物学效应。 为了进一步验证OA 细胞模型中ERS下游CHOP、JNK、Caspase-12 对凋亡的调控作用,笔者在OA 细胞模型中分别进行了CHOP 表达的敲低、JNK 拮抗剂干预、Caspase-12 抑制剂干预, 经上述干预后细胞的增殖水平增加、凋亡率降低,表明OA 细胞模型中ERS 激活通过下游CHOP、JNK、Caspase-12介导了细胞凋亡。
综上所述,IL-1β 诱导软骨细胞炎性损伤模型中ERS 通过CHOP、JNK 和Caspase-12 引起细胞凋亡。在OA 进程中关键软骨细胞在炎症反应的作用下会发生ERS 激活,激活的ERS 进一步通过下游CHOP、JNK、Caspase-12 促进软骨细胞发生凋亡。 根据以上结果, 软骨细胞中ERS 介导的细胞凋亡参与OA 的发病。
