我国玉米南方锈病的抗病育种进展和防治策略
2023-11-18任雅琨朱胜男计怀春王启磊
任雅琨 韩 托 朱胜男 计怀春 王启磊
(1 农业农村部玉米小麦等作物优质抗逆精准育种重点实验室,郑州 450001;2 河南金苑种业股份有限公司,郑州 450001)
玉米南方锈病是由多堆柄锈菌引起的一种气流性传播病害,主要危害玉米叶片、叶鞘。最初广泛发生在非洲、东南亚、美洲南部等热带和亚热带地区,随着全球热带气旋的活动,该病害在全球范围内不断扩大,已在我国22 个省份发生,对黄淮海夏玉米区和南方玉米区的危害尤为严重,严重时可导致减产50%以上。2015 年南方锈病在全国致病区域达到一个高峰,发病面积达523.9 万hm2,实际损失约75.6 万t[1]。2021 年玉米南方锈病在黄淮海区域再次大爆发,山东省发生面积达历史之最,发病面积占山东省玉米总种植面积的43.2%,平均病株率46%[2]。近年来受西太平洋热带气旋频繁发生的影响,南方锈病在我国的发病面积和严重程度不断扩大,对我国玉米主产区的安全生产构成严重威胁。抗病育种是防治南方锈病最经济有效的方法。了解我国玉米品种的抗性水平、鉴定抗病种质、挖掘和利用抗病基因仍是抗病育种的重要工作,并在此基础上探讨了南方锈病的防治策略。
1 我国玉米品种对南方锈病的抗性评价
了解我国当前玉米品种对南方锈病的抗性水平,是品种改良和抗病育种的基础。近年来多位学者对我国参试、已审和主推玉米品种进行南方锈病抗病性鉴定。刘骏等[3]从黄淮海区域47 个主推品种中鉴定出2 份高抗南方锈病品种,14 份抗性品种,先玉335、郑单958、浚单20 均为高感品种。王群[4]在2017-2018 年从山东省400 份玉米品种中鉴定出63 份南方锈病抗性品种,其中高抗和抗性品种分别有3 份和24 份。刘炎琨[5]在2019-2020 年对358份河南省引种和区域试验品种进行抗性鉴定,高抗、抗和中抗南方锈病的品种分别有66 份、80 份和111 份。黄莉群等[6]在2020 年对国内85 个主栽玉米品种进行田间抗性评价,高抗品种仅有9 份,占比10.59%,中抗品种19 份,占比22.35%。郭宁等[7]在2018-2020 年黄淮海夏玉米区国审的437 个品种中未鉴定出高抗品种,抗和中抗品种占比4.45%和4.92%。在47 个主栽品种中,抗性品种和中抗品种占比12.22%和27.66%。总体而言,我国玉米品种对玉米南方锈病的抗性较差,高抗品种少,可能是由于我国玉米育种使用的抗性种质资源狭窄,过度使用78599 系列,造成本土种质和品种趋同化严重且抗性较差[8]。然而,近年来抗病品种比例呈逐年上升趋势,说明科研院所和育种单位对抗锈病品种的选育工作的重视程度增加。因此,拓宽种质资源、创新育种模式以及选用不同的抗病种质仍是未来抗病育种的重要方向。
2 我国玉米抗南方锈病种质的鉴定和利用
2.1我国南方锈病抗性种质的鉴定培育抗病品种的关键是选育抗病自交系,鉴定和利用多种抗病种质是选育抗病自交系的基础。较多学者不断地搜集和鉴定南方锈病抗性种质。黄飞燕[9]对1136 份玉米种质资源进行南方锈病抗性鉴定,高抗、抗、中抗种质所占比例分别为2.46%、7.13%和29.93%,国外地方品种的抗性比例高于国内地方品种的抗性比例。江凯等[10]从1589 份玉米种质资源中鉴定出137 份抗南方锈病材料;高抗材料仅26 份,其中16份来自于国外材料(美国材料选系5 份、CIMMYT材料7 份、非洲津巴布韦材料4 份),国内材料只有10 份。姚国旗等[8]鉴定了34 份热带亚热带自交系,发现仅自交系CML144、CML247、CML451 和CML470 高抗南方锈病。陈文娟等[11]在广西和北京对903 份种质资源做了南方锈病抗性鉴定,高抗材料占比0.9%,抗性材料占比2.3%,中抗材料占比11.1%,高抗和抗性材料主要源自内蒙古和山西。此外作者用56 份抗性材料进行了聚类分析,可将抗性材料分为两大类群,第一大群又被划分成5 个亚群,包含PB 亚群、唐四平头亚群、旅大红骨亚群、BSSS亚群和Lan 亚群;第二大群为PA 群,与其他亚群材料的遗传相似度较低,表明抗病材料有较高的遗传多样性。以上研究结果显示我国国内高抗南方锈病的种质资源较少,且多数高抗南方锈病的资源来自国外热带种质资源。
2.2南方锈病抗性种质资源的利用热带种质资源在我国玉米育种中多数无法直接利用,需要与本土优异种质多次回交,将热带抗病种质中的抗锈病基因导入到本土优异材料中,才能实现本土自交系南方锈病抗性基因的多元化。任转滩[12]将热带种质CML311 与掖478 杂交,经多代混粉后自交选择,选出了既具有掖478 的优异性状又兼备南方锈病抗性的新材料;用同样的方法将CML246 与CA971杂交后再与Mo17 杂交,经多代自交选择,最终筛选出了高抗南方锈病的适应性种质;除此之外,B73、K12、黄早四、黄金96B 等常用自交系也通过此方法提高了抗锈病能力。张志方等[13]以高抗南方锈病的M119 为基础材料,经多代自交和姊妹交,经河南、贵州、海南三生态区的混合抗性选择和配合力测定,选出了塘四平头类型的抗病优异自交系浚M9;之后用浚M9 对本地种质进行改良,形成高抗茎腐病和南方锈病的优异种质浚96、浚M97、浚M98。同时用浚M9 与国外优异种质和国内骨干自交系进行选系,构建了1 个广适多抗父本群。新的抗病自交系的育成也证明了利用热带抗病资源培育本土化抗病自交系的可行性。进一步利用CIMMYT、Suwan 和非洲等热带地区的抗南方锈病种质资源,仍是抗病自交系选育的关键环节。这些资源能够为我国抗病育种提供非常优秀的抗性基础材料。
3 玉米南方锈病抗病基因的挖掘及应用
3.1玉米南方锈病抗病基因的定位和克隆基于南方锈病的发病范围不断扩大,对玉米产量造成的损失程度不断增加,国内外科研人员对玉米南方锈病基因的挖掘开展了较多研究。Storey 等[14-15]从抗南方锈病材料AFRO.29 和AFRO.24 中分别发现了完全显性抗病基因Rpp1和不完全显性抗病基因Rpp2,Rpp1专抗小种EA1,Rpp2对小种EA1和EA2 具有部分抗性;Rpp1与Rpp2连锁,遗传距 离 约 有12.23cM。Ullstrup[16]从PI 186208 中 定位到一个专抗小种PP.9 的单显性抗病基因Rpp9;Storey 等[17]又发现了对EA1 和EA3 有抗性的显性基因Rpp10和对EA1 和EA3 有不完全抗性的抗病 基 因Rpp11,且Rpp1、Rpp10和Rpp11均 不 连锁。我国研究显示多数南方锈病抗病位点定位在10 号染色体短臂上,且为单显性主效基因。不同抗性材料中定位的抗性基因及连锁的分子标记见表1[18-28]。Zhou 等[18]将齐319 的抗病基因RppQ定位于分子标记MA7、M-CCG/EAGA157 之间。Liu等[19]将抗性基因RppP25定位在10 号染色体上,与phi059 的遗传距离为5.8cM;Zhao 等[20]进一步将RppP25定位在分子标记P091 和M271 之间,GRMZM2G060884可能为候选基因。Zhang 等[21]从W2D 和W222 构建的分离群体中定位主效单显性抗病基因RppD,与分子标记umc1291 和CAPS858的遗传距离分别为2.9cM 和0.8cM。RppD与RppQ和RppP25不等位。Wu 等[22]用热带材料SCML205与京24 构建F2分离群体,将抗病基因RppS定位在10 号染色体短臂远端,与IDP4283 和G047 的遗传距离为8.4cM。RppS与RppQ、RppD和RppP25均不等位。姚国旗等[23]发现CML470 的抗性主要由显性单基因RppC控制,该基因定位在分子标记umc1380 与umc1291 之间,与2 个分子标记的遗传距离分别为3.5cM 和8.8cM。王兵伟等[24]用S313×PHW52 的F2分离群体将抗病基因RppS313定位在SNP 分子标记A005915 与 A009920 之间,区间内有3 个候选基因。艾堂顺等[25]从P178 和G41 构建的BC2F5群体中发现主效抗南方锈病位点qSCR10.01,可解释45.31%的表型变异,该QTL 被定位在分子标记umc1380 和C(10)3595071 之间,物理距离1.34Mb。王文洁等[29]将高抗南方锈病材料K381 的抗病位点定位在分子标记umc2018 与phi059 之间,与phi059 紧密连锁。
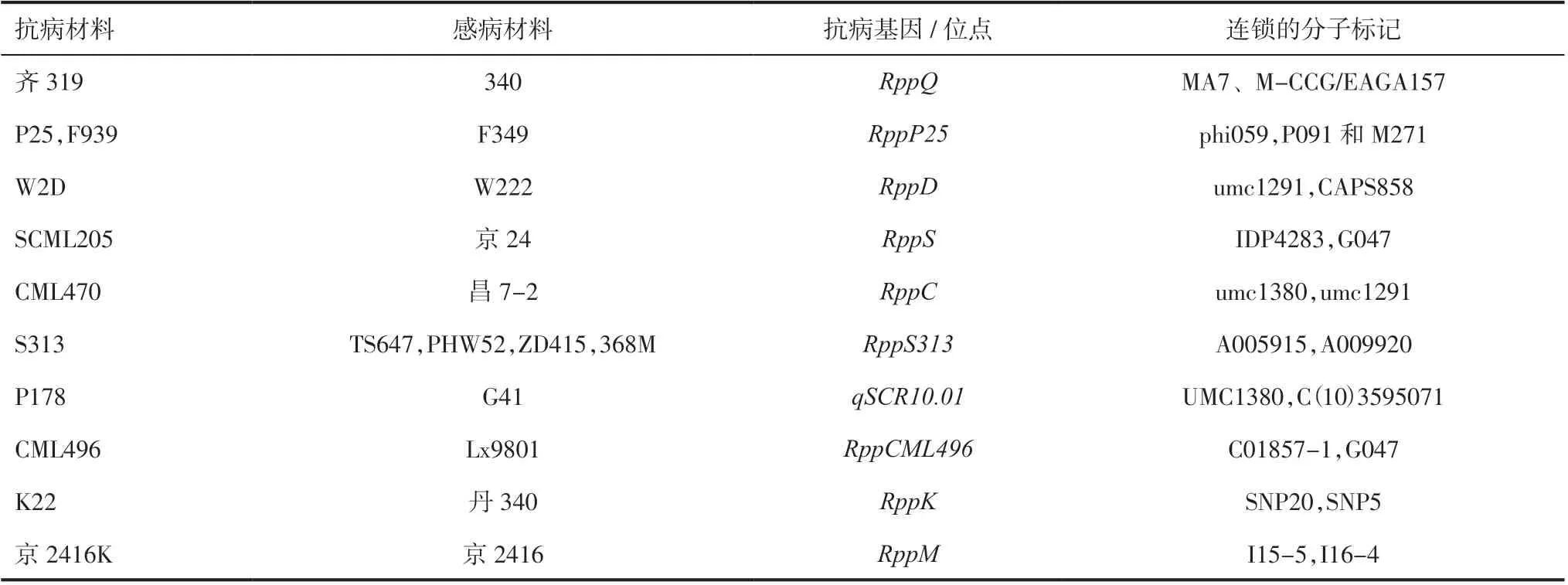
表1 不同抗性材料中抗性基因及连锁的分子标记
除主效基因的定位,科研人员在玉米的1、2、3、4、5、6、8、9 号染色体上也发现了南方锈病的抗病位点[25,30-32]。其中Lu 等[30]用齐319 和掖478 构建重组自交系,在第6 号染色体上发现的qSCR6.01位点抗病效应最大,可解释表型总变异的24.15%。
南方锈病抗性位点报道较多,但目前只有3个 抗 病 基 因 被 克 隆。Lv 等[26]和Deng 等[27]以CML496 和Lx9801 构建的重组自交系为材料,采用图位克隆和转基因验证的方法,从CML496 的10 号染色体上克隆到一个NLR 类的抗玉米南方锈病的新基因RppC。利用转录组分析和玉米原生质体超敏反应筛选体系等多种研究方法,在南方锈病病原菌中鉴定到被RppC识别的效应蛋白AvrRppC,并发现新的病原菌小种通过AvrRppC氨基酸突变的方式来逃避RppC的识别。Chen 等[28]用K22 和丹340 构建的F6:7分离群体和重组自交系,在10 号染色体上定位到一个主效的单显性基因RppK,转基因验证确定K22 基因组中的R3为抗病基因,编码典型的NLR 蛋白,能够与AvrRppK互作激活植物体内的ETI 反应,从而提高植物抗病性,在抗病育种上应用潜力巨大。Wang 等[33-34]通过对京2416K 和京2416 构建的F2群体进行定位、测序分析和转基因验证,在10 号染色体短臂上定位并克隆到南方锈病抗性基因RppM,并且基于RppM两个保守位点(AA576、AA817)开发了可以鉴定RppM的特异性KASP 分子标记KM23 和KM19。
3.2南方锈病抗病功能基因在玉米品种改良上的应用目前已克隆抗南方锈病基因RppC、RppK为基因与基因互作,进而触发植物体的ETI 和PTI防御反应,提高植物的抗病性,在育种中有较高的应用价值[27-28]。RppC在商业品种中应用较广,裕丰303、中科玉505、农大108、汉单777、登海3622、蠡玉16、蠡玉88、农大108、豫单9953 等多个主栽抗锈病品种均含有RppC基因,具有较好玉米南方锈病抗性[33];12 个浚单玉米自交系含有RppC抗病基因[35];在审定品种中辽单707、康农玉598、康农2 号、高玉14022 对玉米南方锈病有较好抗性,经检测均含有抗病基因RppK[28],含有RppM的京2416K自交系已成为我国高抗玉米南方锈病的核心种质,为我国玉米南方锈病品种的改良提供了基础材料,先后组配出京农科767、京科682、现代965 等玉米品种,对南方锈病具有较好抗性。
4 玉米南方锈病的防治策略
4.1挖掘抗病基因进行种质创新和抗病育种利用抗病基因进行抗病种质资源创新和品种选育是控制玉米南方锈病最经济有效的方法。RppQ、RppP25、RppD、RppC、RppK和RppM等优良抗病基因的定位和克隆以及与抗病基因连锁的分子标记的开发,促进了传统育种与现代育种技术相结合,为抗病种质资源创新和品种选育、改良提供了更精准有效的方法。还应加大对含有未知抗病基因的高抗南方锈病的基因挖掘工作,加强高抗南方锈病的热带材料在本土优异自交系的改良和品种选育方面的应用。另外分子标记辅助育种、单倍体育种、全基因组选择和基因编辑等现代育种技术为南方锈病抗病育种提供了一种更精确快速的途径。常规育种和现代分子技术相结合,可加快抗病育种进程。
4.2加强抗病种质的利用南方锈病的高抗种质在国内较少,已鉴定的抗病种质多为热带或亚热带材料。对已鉴定的国内高抗南方锈病的种质可以直接利用或者进行骨干材料的改良。CIMMTY、Suwan、非洲津巴布韦等高抗南方锈病的材料不能直接利用,需多代轮回选择改良自交系。在改良过程中,可使用分子标记辅助选择进行背景筛选,利用与南方锈病抗病基因连锁的分子标记进行前景筛选,结合单倍体等现代育种技术可以加快品种改良进程,但材料改良还要以接种抗病鉴定结果和配合力测定为主;改良的育种材料要在黄淮海、海南、广西等多个生态区进行田间抗性鉴定,更有利于选出广谱抗性的优质自交系。
4.3加强病源菌的监测和预警我国南方锈病的发生和热带气旋活动路径有关,在锈病发生前期结合气象变化,监测病源菌的动态变化,为病情发生做出预警。使用南方锈病特异性分子标记和快速检测体系监测南方锈病在田间的侵染动态[36];发病严重区域和经常发病区域可建立玉米南方锈病的病害预测模型,为病情预测提供指导[30]。加快开发和利用对南方锈病检测准确度高的田间无损化监测工具[37],建立玉米不同生态区的南方锈病病害预测模型,在病害发生前做出预警并及时采取干预措施,可有效抑制病源菌的侵染和扩散,防止病害的大规模爆发。
4.4适期配合使用化学防控监测玉米南方锈病的侵染动态,在适宜的时期及时施用化学药剂能显著抑制南方锈病的发生和蔓延[38-40]。在南方锈病初发期,用250g/L 的吡唑醚菌酯乳油、125g/L 的氟环唑悬浮液、5%己唑醇、25%丙环唑和43%戊唑醇处理,可显著抑制病害的蔓延,降低病害对植株和产量的危害程度[38,40]。大喇叭口期至抽雄期是南方锈病激发期;在大喇叭口期喷施戊唑醇悬浮剂430g/L、氟硅唑乳油400g/L、吡唑醚菌酯乳油250g/L、30%肟菌·戊唑醇悬浮剂、吡唑醚菌酯乳油250g/L 与戊唑醇悬浮剂混剂430g/L,可对南方锈病有较好的防治效果[39]。由于大喇叭口期玉米植株高大、田间郁闭,玉米病虫害防治比较困难,建议有条件的可用植保无人机进行飞防;进行人工防治时,可以用高压喷枪进行粗防或进行精细喷雾防治,控制病情,抑制病源菌的繁殖、扩散和传播。
4.5加强玉米主产区致病小种的鉴定工作前人研究初步确定了我国多堆柄锈菌的主要来源,辽宁、黄淮海区域和浙江、福建的南方锈病初始菌源主要来自我国台湾地区;广东、广西和海南的初始菌源主要来自菲律宾;云南、贵州等西南地区初始菌源主要来自泰国、老挝、缅甸等国家,并在境内形成病害的周年循环[41]。我国不同玉米生态区的气候差异较大,不同区域的南方锈菌的病原小种复杂。海南菌群年度间遗传差异较大,来源较多且不稳定;山东多堆柄锈菌遗传多样性丰富,说明病原小种较多;广西、广东、浙江、福建、海南的初始菌源相近,但广西河池的菌株致病性最强。致病小种的更迭或病菌变异会导致推广的抗病品种丧失部分抗性,因此加强各区域的致病小种的鉴定有助于开展针对性的抗病育种。
随着全球气候变暖,暴雨、台风等灾害性气候频发,南方锈病在我国玉米主产区的发病面积不断扩大,后期爆发速度快,尤其对黄淮海区域和南方沿海地区的玉米产量危害严重。抗病育种是病害防治最绿色、经济、有效的措施。但我国高抗南方锈病种质资源较少,且抗病种质存在一定的地域限制,因此加快抗病种质的搜集,对已鉴定出的高抗种质加以利用,建立起科研院所和育种企业自己的核心抗病自交系尤为重要。深度挖掘南方锈病抗病基因,利用已开发的分子标记和前沿分子育种技术培育抗南方锈病品种。推广部门重点推广抗性品种,同时在抗病品种不多的情况下,动态监测病害的变化,及时有效地采取化学防控,多措并举,以保证玉米主产区的安全生产。
