海洋酸化与水体净碳酸钙形成/溶解的关系
——以黄海冷水团为例
2023-11-15翟惟东
翟惟东
(南方海洋科学与工程广东省实验室(珠海),广东 珠海 519082)
工业革命以来,化石燃料的使用和土地利用改变等人类活动显著地改变了全球碳循环,使得大气CO2摩尔分数由工业革命前的约280×10−6升高至2020年的412×10−6。这期间,海洋持续吸收人类来源的CO2,使得全球海表pH平均下降0.1~0.2,引起海洋酸化现象(Caldeira et al, 2003; Orr et al,2005; Doney et al, 2009)。由于海洋酸化直接导致海表碳酸钙饱和度(Ω)下降,因而将严重影响海洋钙化生物的钙质骨骼/外壳的形成与维持。有实验证据表明,严重的海洋酸化对一些鱼类的正常生理活动和行为特征也会造成干扰(Cattano et al, 2018; Esbaugh, 2018; Porteus et al, 2018; Rong et al,2018)。
与开阔大洋相比,近海的海水酸化过程机制更加复杂,发展进程更快。除了沿岸上升流和河流冲淡水给一些近海海域引入低Ω水体(Feely et al, 2008; Salisbury et al, 2008; Lui et al, 2015; Zhai et al,2015)加剧海洋酸化的负面影响以外,在有季节性层化现象发生的高生产力近海水域,藻类或其他生源颗粒沉降之后在跃层以下的水体里发生微生物介导的耗氧矿化作用(Li et al, 2018),同时释放CO2造成酸化,而层化现象又阻碍了酸化水体通过气体交换排酸,使得这种海域表层以下的自然生态系统和养殖生物早早地受到海水酸化的威胁(翟惟东等, 2012; Zhai et al, 2014)。这种在近海发生,与群落呼吸相伴的酸化现象是局部的和季节性的(Borges et al, 2010; Feely et al, 2010; Melzner et al,2013),不同于响应全球大气CO2升高的总体性的“海洋酸化”问题(Duarte et al, 2013)。然而,近海季节性海水酸化必然与响应大气CO2升高的海洋酸化叠加起来,共同作用,甚至因为高CO2条件下海水对pH的缓冲容量下降,而使得这两种酸化现象的负面效应相互加强(Cai et al, 2011)。
近年来,在大气CO2升高导致趋势性的海洋酸化以及近海富营养化、河流冲淡水、沿岸上升流等因素导致近海季节性酸化的双重背景下,国内在很多海域开展了大量海水pH和Ω的时空分布及生物地球化学调控方面的调查研究工作,也有很多研究通过CO2加富培养的方法分析受控条件下各种生物对海水酸化的生理生态响应,然而还罕有在生态系统水平上基于碳酸盐体系参数评估水体净碳酸钙形成/溶解的研究报道。本文主要基于黄海的研究工作对此进行评述,对我国后续的海洋酸化监测与生态效应评估工作有一定借鉴意义。
1 海洋酸化的生物生态效应
海水pH和Ω都是海水化学环境的重要指标。正常海水的pH在8.1左右,偏碱性。当海水pH低于正常值,就表明海水中酸性物质积累,大多数海洋生物的正常生理活动都会受到干扰。Ω则是海水中碳酸钙颗粒(CaCO3)形成的化学势,其定义为海水中钙离子浓度和碳酸根离子浓度的乘积与文石型或方解石型CaCO3表观溶度积的比值。珊瑚、贝类等钙化生物的骨骼或外壳以碳酸钙为主要成分,从化学定义来看,这些钙化生物适于在碳酸钙饱和度高于1的海水中生长,而直接暴露在碳酸钙饱和度低于1的海水中的任何钙质物体都将趋于溶解。实际海水环境则远比化学定义更加复杂,例如,中、低纬度海水环境中一些珊瑚的钙质骨骼在文石型碳酸钙饱和度(Ω文石)还高达2~3.5时就会发生溶解(Yamamoto et al, 2012; Eyre et al, 2018)。另有研究表明,贝类幼体的生长和发育在很大程度上与二氧化碳分压和pH值都不十分相关,而直接受制于Ω文石(Waldbusser et al, 2015),当Ω文石低于1.5时,幼体贝壳的发育明显受到抑制(图1)。
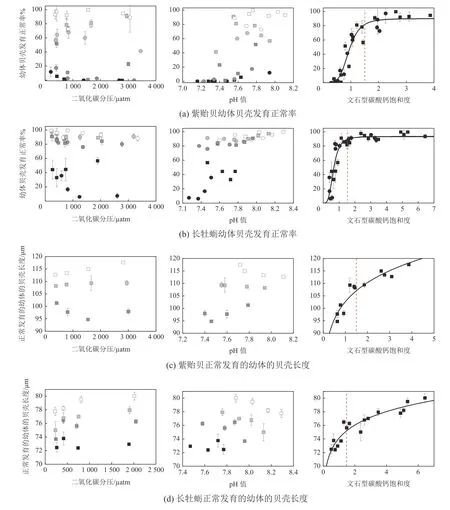
图1 美国近海2种贝类生物的幼体贝壳发育正常率和正常发育的幼体的贝壳长度与海水碳酸盐体系诸参数的关系Fig. 1 Relationship between carbonate parameters and the shell development of two species of shellfish along the United States coast
对不同CO2水平的自然生态系统开展对比研究的结果表明,海洋酸化条件下贝类群落可能被迫向小型化的方向演化(Garilli et al, 2015),相应地,其筑礁、滤食等生态功能减弱,经济价值下降。尽管海洋酸化对贝类生长的负面影响可以在很大程度上通过补充营养而加以改善(Ramajo et al, 2016;Leung et al, 2019),然而实验室研究表明,酸化的海水环境严重干扰贝类的行为(Peng et al, 2017)、免疫(Liu et al, 2016; Su et al, 2018)与能量代谢(Liao et al, 2019),因此,单纯补充营养而不考虑水化学环境改变的做法是有条件并且是有限度的。有研究表明,生活在秘鲁沿岸强上升流海域的智利扇贝可能已适应当地酸化环境(Ramajo et al, 2019),但是大多数贝类暴露于低pH海水中就会出现呼吸、排氨等正常代谢活动受到干扰的情况(Liu et al, 2012; Zhao et al, 2017)。另一项实验室研究则显示,采集自黄海沿岸潮间带的底栖有孔虫群落对持续数月的海水酸化呈现出负面响应(Dong et al, 2020),因此,即使是生活在高度动态变化的潮间带环境里的钙化生物,其对海水酸化快速发展这样的环境胁迫仍然难以适应。
2 在生态系统水平上基于碳酸盐体系参数对水体净碳酸钙形成/溶解的评估
2.1 原理与方法
边缘海碳酸盐体系动态变化受到群落生产和呼吸作用、生物钙化作用和碳酸钙溶解,以及水体混合、温度变化、海气交换等多种过程共同影响。鉴于北黄海冷水团在每年春季到秋季是一个持续存续的稳定水团,并且水体混合和海气交换对冷水团季节变化的影响可以忽略,而溶解无机碳(Dissolved Inorganic Carbon,DIC)和总碱度(Total ALKalinity,TAlk)又不受温度变化影响,所以,冷水团DIC和TAlk的季节变化就只受群落生产和呼吸作用、生物钙化作用和碳酸钙溶解这2组过程的调制。
冷水团中的生物好氧呼吸作用可描述为:
式(1)表明,生物呼吸作用释放CO2进入水中,每消耗138 mol O2,DIC增加106 mol,TAlk减少17 mol。
而碳酸钙溶解过程可以描述为:
式(2)表明,每当1 mol CaCO3溶解时,海水DIC将增加1 mol,TAlk将增加2 mol。仿照Chen(1978)分析深海碳化学调查资料的思路,并结合式(1)和式(2),则如果Xmol有机物和Ymol CaCO3分解,DIC和TAlk的总变化将为:
将式(3)和式(4)联立解方程,可以得到:
因此,当忽略温跃层上下的海气交换和垂直混合,并忽略其与其他水团的混合作用时(从春季到秋季,黄海冷水团的盐度只有很小的变化),就可以根据式(5)和式(6),通过DIC和TAlk的月际改变(ΔDIC和ΔTAlk)估算基本生物地球化学过程的贡献率。
在实践中,冷水团基本生物地球化学过程对碳酸盐体系参数改变的贡献率可由北黄海冷水团内相邻调查航次期间各参数平均值的变化量来计算,也可由邻近的并且相邻月份间盐度变化很小(说明水体基本保持在原地)的站位数据来计算。在本研究中,相邻月份间邻近站位的选择标准是:经、纬度的差别控制在0.5°以内,底层水盐度的差别控制在0.10以内,并且综合考虑水温的月际变化是否合理,进而从空间覆盖度比较充足的月际调查数据集里面筛选出尽可能符合条件的碳酸盐体系数据。
2.2 北黄海的数据与结果
2011年、2013年和2017年间,在北黄海以冷水团为中心开展了若干月际调查(Zhai et al, 2014,2015; Li et al, 2021),利用冷水团底层水的平均数据及相邻月份间邻近站位的可比数据,以式(6)结合相邻调查之间的时间间隔,计算出北黄海冷水团海域的群落净钙化作用/碳酸钙溶解速率。数据结果呈现出春夏两季发生净钙化作用而秋季发生净的碳酸钙溶解的总体格局(表1)。数据结果还表明北黄海冷水团的群落净钙化作用/碳酸钙溶解速率与水体Ω文石正相关(图2),并首次在中国近海揭示出,群落净钙化作用速率降为 0 (μmol·kg−1)·d−1的Ω文石阈值为1.5~1.6(李成龙, 2019; Li et al,2019),与Waldbusser等(2015)报道的美国海岸带贝类幼体正常生长的临界阈值(图1)基本吻合。这一结果对于正确认识海洋酸化对中国近海的环境影响非常重要。
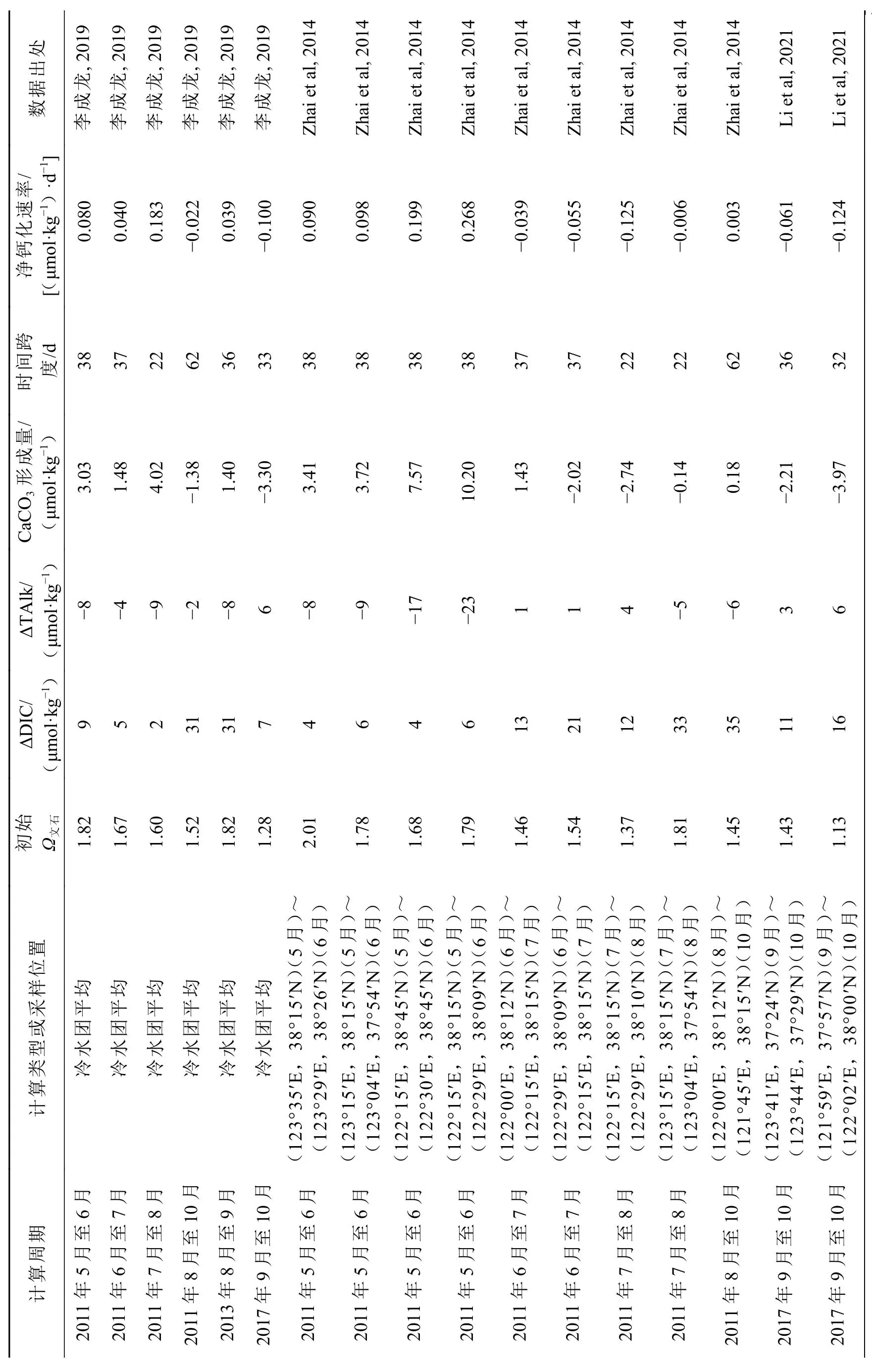
表1 北黄海冷水团的Ω文石与计算的群落净钙化速率之间的关系Table 1 Calculation of net community calcification rate and its relationship with aragonite saturation state in the North Yellow Sea cold water mass
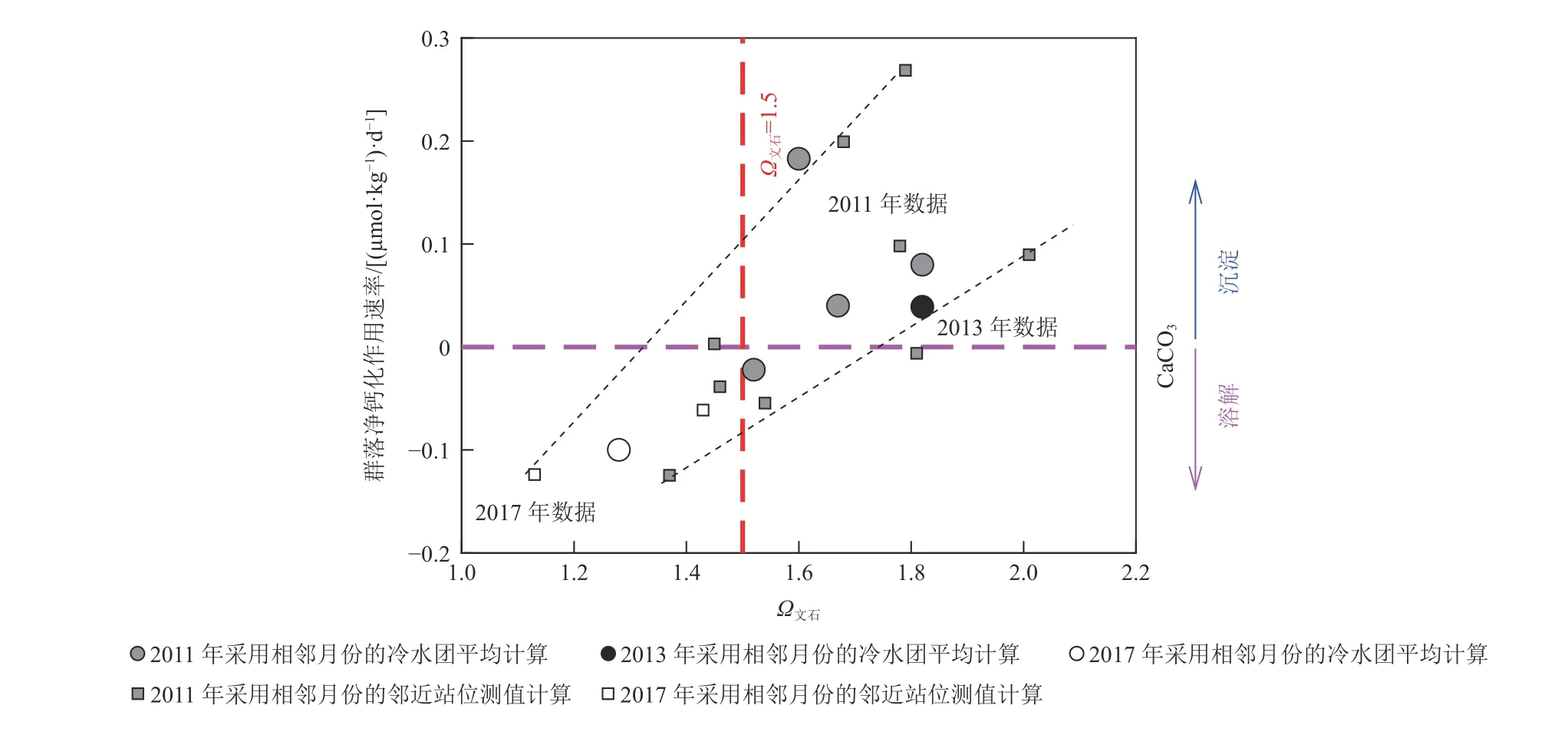
图2 北黄海冷水团群落净钙化速率与文石型碳酸钙饱和度之间的关系Fig. 2 Net community calcification rate versus bottom-water aragonite saturation state (Ωarag)in the North Yellow Sea cold water mass
2.3 与珊瑚礁系统数据结果的比较
需要指出的是,珊瑚礁系统的群落净钙化作用速率降为0 (μmol·kg−1)·d−1的Ω文石阈值高达(2.9±0.2)(Eyre et al, 2018),与北黄海冷水团的Ω文石阈值很不相同,说明不同钙化物种或不同生态系统的情况差别较大,需要分别开展研究。尽管如此,国际上一些权威的近海酸化评估研究报道大都把Ω文石=1.5作为海水碳化学变异对贝类等钙化生物构成严重威胁的临界指标(Gruber et al, 2012;Ekstrom et al, 2015)。
3 讨论
3.1 黄海冷水团季节性酸化的过程机制
在黄海开展月际调查的数据比较结果表明,黄海底层大水体的耗氧作用速率为每天0.4~1.5 μmol O2kg−1(Zhai et al, 2014; Li et al, 2019; Yu et al, 2022),比长江口外富营养化海域2003年6月表层水体实测耗氧作用速率——每天2.2~34.4 μmol O2kg−1(Chen et al, 2006)低得多,也比邻近的渤海中部海域底层水体的最高耗氧作用速率——每天2~3 μmol O2kg−1(翟惟东等, 2012; Song et al, 2020)略低。黄海生物群落净耗氧的同时,释放的CO2造成水中溶解无机碳升高(翟惟东, 2018; Yu et al,2022),导致pH和Ω文石下降(图3)。在南黄海的冷水团区域,2019年从4月至8月pH值可下降0.17,Ω文石下降0.55(即年度初始水平的27%)(Yu et al, 2022),相当于大洋表层过去150 a或未来50~150 a的酸化进程,从而对水生生物产生较大的环境胁迫。
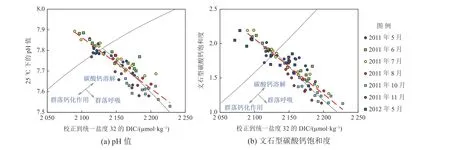
图3 北黄海冷水团海水酸化指标pH值和文石型碳酸钙饱和度与溶解无机碳浓度之间的关系Fig. 3 Relationship of pH values and aragonite saturation state versus salinity-normalized dissolved inorganic carbon concentration
除耗氧过程以外,碳酸钙溶解(式(2))及钙化过程也会影响海水酸化指标,前者消耗CO2并且释放碱度(HCO3−),可在一定程度上缓解海水酸化,后者则导致海水酸化程度加剧。分析北黄海2011年至2012年碳酸盐体系调查数据的结果(图3)显示,该海域呈现出碳酸钙过程季节循环的现象,即2011年5月至8月存在钙化作用信号,而2011年10月则表现为碳酸钙净溶解(Zhai et al,2014)。基于2011年至2016年数据的定量评估结果则表明,北黄海碳酸钙过程对pH值月际变化和碳酸钙饱和度月际变化的贡献远低于呼吸/矿化作用(Li et al,2019)。
就长期变化而言,国家海洋局北海环境监测中心在北黄海西侧一条西北—东南走向断面(121°42′~122°12′E,38°00′~38°48′N)开展过长达40 a的季节性调查,结合他们的实测温度、盐度、溶解氧和pH数据集综合分析表明,北黄海冷水团在过去40 a中呈现出夏秋季表观耗氧量(AOU)和海洋酸化指标的季节性变化幅度都显著增大的现象(Wei et al, 2019; Li et al, 2022)(图4)。北黄海冷水团严重酸化(秋季平均Ω文石<1.5)是本世纪以来才出现的,而夏秋两季的海水酸化都严重(Ω文石<1.5)的情况则是2010年以来的事情(Li et al, 2022)。耐人寻味的是,2014年以来大连獐子岛海域频繁爆出底播扇贝大量逃逸或死亡的灾害现象,这与北黄海夏秋两季都发生严重海水酸化仅发生于2010年以后的情况基本上能对应,说明海洋酸化或许已经对我国部分海域的海水养殖业构成现实威胁。
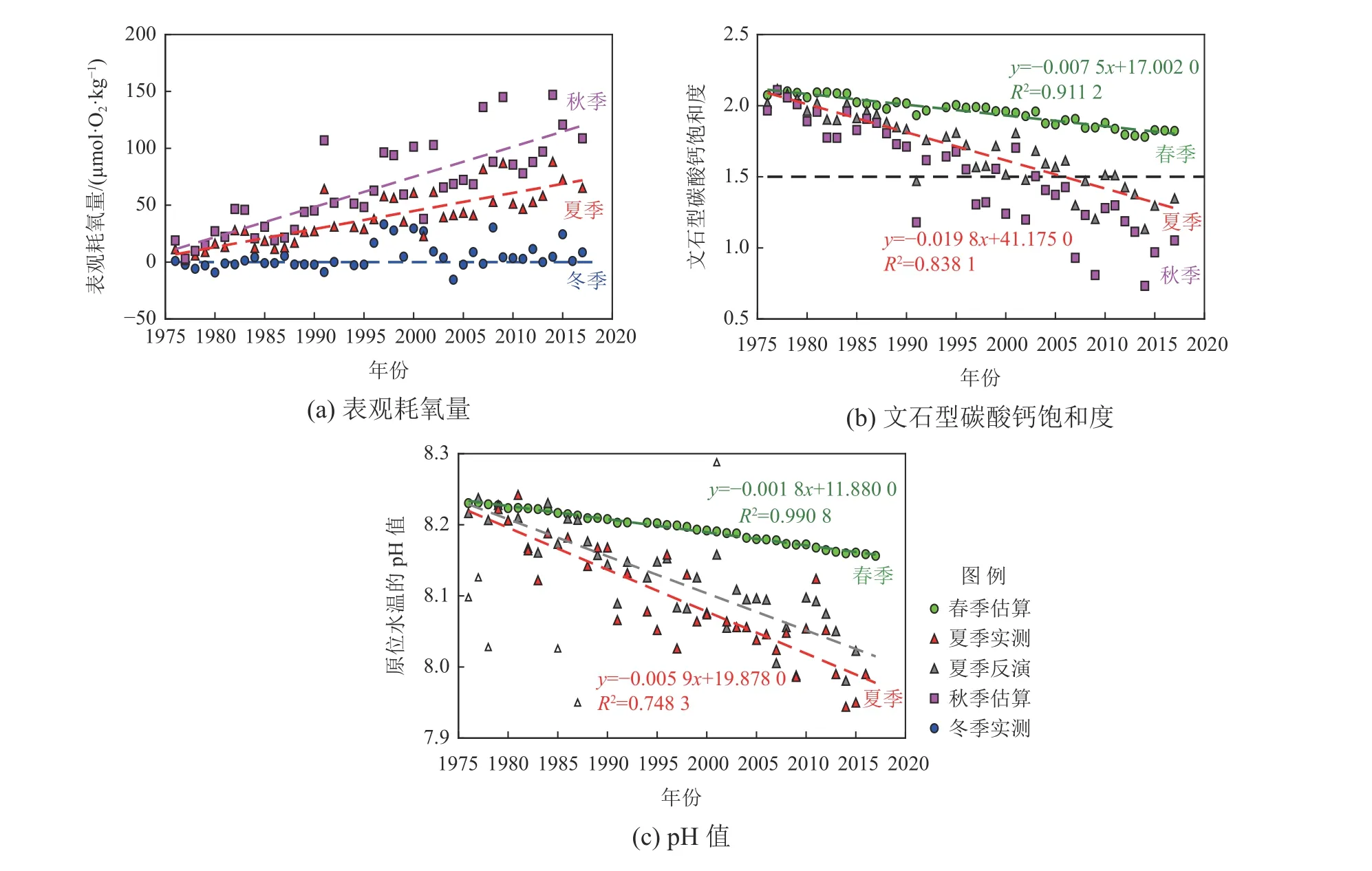
图4 北黄海西侧断面底层水体表观耗氧量、文石型碳酸钙饱和度和pH值的季节变化幅度显著增加的现象Fig. 4 Evolution of amplitudes of seasonal variations in apparent oxygen utilization, aragonite saturation state,and in situ pHNBS values from spring to autumn
基于反演数据的趋势分析(图4b)显示,北黄海冷水团夏季Ω文石在1976年至2017年间的平均下降速率为每年(−0.0197±0.0014),其中由于全球大气CO2升高导致的变率为每年−0.0070,只占35%,而区域富营养化导致的耗氧量增加导致的变率为每年−0.0121,贡献率高达61%,其余4%左右是由水温和盐度的略微改变造成的(Li et al, 2022)。类似地,北黄海冷水团夏季实测pH值在1976年至2017年间的平均下降速率为每年(−0.0058±0.0005)(图4c),其中由全球大气CO2升高导致的变率为每年−0.0018,只占31%,而区域富营养化导致的耗氧量增加导致的变率为每年−0.0039,贡献率高达67%,其余2%左右是由水温和盐度的略微改变造成(Li et al, 2022)。这表明,北黄海冷水团过去40 a的酸化趋势主要是由于区域富营养化引起生态系统净呼吸加剧造成,该因素的贡献率在夏季是大气CO2升高的贡献率的2倍左右(Li et al, 2022)。由于黄海冷水团在秋季积累的表观耗氧量及过饱和CO2更多(图4a)( Yu et al, 2022),预计区域富营养化引起生态系统净呼吸加剧的因素在秋季对北黄海冷水团40 a酸化的贡献率将更高。这再次说明,区域生物地球化学改变作用于边缘海海洋酸化指标的强度可以大大超过大气CO2升高的影响(Borges et al, 2010),必须引起重视。
3.2 发展动态分析
近海海洋酸化的实证研究从Feely等(2008)、Salisbury等(2008)、Feely等(2010)和Cai等(2011)算起,迄今已逾15 a。其化学成因解析基本告一段落,当前研究大多聚焦于生物或生物群落对酸化环境的响应及适应方面,以实验室内或各种围隔体系的模拟实验研究为主要技术手段,然而基于生态系统过程分析手段直接调查生态系统响应的研究设计(Eyre et al, 2018;Li et al, 2019)还比较少。
研究显示,棘皮动物对黄海大型底栖生物多样性的贡献率正呈下降趋势,据认为可能跟海水酸化有关(Jin et al, 2015)。目前,黄海底层海水酸化程度严重威胁底栖贝类的海域(Ω文石<1.5)面积在每年夏、秋季都已达黄海调查面积的1/3左右(翟惟东, 2018; Xiong et al, 2020),甚至在南黄海中南部海域偶尔出现Ω文石<1.0的底层水(Choi et al, 2020)。近期报道的南黄海韩国一侧底层海水季节性酸化调查结果(Kim et al, 2023)显示,该处底层水Ω文石在秋季可低至1.15,秋季底层水Ω文石平均值为1.57,而在水温不到12 °C的冷水团区域,秋季底层水Ω文石都在1.30以下,平均值只有1.22。这样的观测结果与黄海中国一侧的数据报道比较接近,都表明黄海冷水团区域的底栖钙化生物正在承受海水碳化学环境改变造成的重大环境胁迫。
据推测,2050年前后大气CO2摩尔分数可能升高至500×10−6,届时如果黄海的水文条件和群落呼吸速率不发生大幅改变的话,则黄海50%左右海底(主要是冷水团区域)在夏、秋季都将被Ω文石不足1.5的季节性酸化水体覆盖(翟惟东, 2018),底栖钙化生物将承受比当前更大的环境压力。实验室研究结果显示,当Ω文石从2.56下降到1.24,黄海底栖有孔虫的净钙化作用速率将下降89%(Dong et al, 2023)。因此,这种连续半年或更长时间的环境胁迫将如何影响黄海底栖生态系统的结构与功能,以及近海生物地球化学将如何响应,都是迫切需要深入研究的现实问题。此外,由于近海的水动力环境十分复杂,有时酸化的冷水团海水会在潮流或风海流等因素的驱动下脉冲式影响周边的贝类栖息地和养殖区,从而造成难以预料的严重损失。这对于黄海周边那些依赖冷水条件的海水养殖企业而言,也是个需要高度重视的环境风险。
在发达国家,一些产业经济学家的预测分析结果显示,严重的海水酸化将对沿海地区的贝类养殖业和相关的海产品供应造成极大冲击(Narita et al, 2012)。据估算,欧洲的贝类养殖业到本世纪末可能因海洋酸化而每年损失10亿美元(Narita et al, 2017),仅英国贝类养殖与消费的损失就可能高达每年2300万至8800万英镑(Mangi et al, 2018)。鉴于此,美国科学家倡导开展海洋酸化对海洋产业的影响评估(Ekstrom et al, 2015),包括脆弱性(包含经济损失评估)和适应性评估两类指标。从脆弱性评估框架来看,海水酸化脆弱性评估至少要包含3个方面:一是某个地域海洋生态系统面对海水酸化的暴露性;二是当地社会的易感性,主要指当地对可能受到海水酸化影响的渔业的依存度;三是当地人们的适应性,便于当地渔民或者产业对未来海水酸化可能造成的影响早作准备,并进行产业(宏观)和生计(微观)调整。其中易感性和适应性为社会脆弱性,暴露性为自然脆弱性。
4 结语
本文综述了黄海冷水团海洋酸化研究的进展,其中,利用2011年、2013年和2017年月际调查获取的碳酸盐体系数据资料,研究得出,北黄海冷水团的净钙化作用速率与底层水体的文石型碳酸钙饱和度正相关,而北黄海冷水团净碳酸钙生产与净碳酸钙溶解之间的临界阈值是文石型碳酸钙饱和度降至1.5~1.6。这对我国后续的海洋酸化监测与生态效应评估工作有一定借鉴意义。
鉴于近海季节性海水酸化可在两三个月时间内迅速发展,其在数月内的环境效应相当于大洋表层未来50~150 a的酸化进程,从而对水生生物产生较大的环境胁迫,甚至形成生态灾害。因此,通过更多现场数据和旁证资料来理解其调控和影响,以期在未来全球海洋环境的变局中提前规避一些产业风险,既有重要的科学意义,也对黄海冷水团资源开发和深水养殖业部署具有指导意义。只有在提升科学认识的基础上提前布局防控措施抵御环境风险,才能有效维护产业的平稳发展。
致谢:作者指导毕业的研究生李成龙在数据分析方面给予了很大帮助,国家自然科学基金委共享航次计划及原中国海洋大学王厚杰教授、中国水产科学研究院黄海水产研究所单秀娟研究员等渤黄海共享航次项目首席科学家群体给作者提供了大量航次机会,在此一并致谢。
