桉树引种对林下植物功能群的物种组成及其多样性的影响
2023-11-13唐媛媛邓福英赵筱青陶俊逸周世杰储博程
唐媛媛 邓福英 赵筱青* 黄 佩 陶俊逸 周世杰 储博程
(1.云南大学国际河流与生态安全研究院,昆明 650500; 2.云南大学地球科学学院,昆明 650500; 3.云南风蓝项目投资咨询集团有限公司,昆明 650500)
桉树(Eucalyptus)是我国南方人工林的重要优势树种,其发展产生了巨大的经济和社会效益[1]。但随着种植规模扩大,桉树纯林会不会破坏生态环境这一问题引发了民众和学者的关注与争论,尤其对生物多样性的影响一直是争论的焦点[2]。如Lemessa 等[3]的研究表明天然林的林下植物和土壤种子库物种丰富度高于桉树人工林;Zhou 等[4]的研究结果显示植物多样性会随着桉树轮伐次数的增加而下降;温远光等[5]认为桉树连栽降低林下植物的物种多样性。但Otuba 等[6]的研究表明,桉树林的植物多样性高于其他常见的热带和亚热带人工林;Michelson[7]的研究发现雨量丰富地区或经营良好的桉树人工林林下植物种类极为丰富。总之,桉树林对林下植物物种多样性有何影响尚无定论。
物种多样性是生物多样性最直接的体现[8],是维持生态系统结构、功能和稳定性的基础[9]。林下植物是人工林生态系统的重要组成部分,在保护物种多样性、维持地力、促进森林群落恢复等方面发挥关键作用[10]。林下植物物种多样性在一定程度上可体现生态系统健康水平、反映森林可持续经营水平[11],开展桉树林下植物物种多样性的有关研究,有助于回答桉树引种是否降低植物多样性的争议问题,对桉树林经营管理具有重要参考价值。但以往对桉树林下植物物种多样性的研究主要是分灌木层、草本层分析林下植物组成和多样性特征,不能深入解释物种在森林生态系统中的运作机制[12-13]。而植物功能群将对环境适应特征相似的物种聚集,能体现植物群落对环境变化的响应[14],有助于评价生态系统功能和结构的完整性、系统稳定性[15]。目前,国内外开展的植物功能群相关研究多是针对淡水生态系统[16-17]和草原生态系统[18-19],对于森林生态系统植物功能群的研究还较少。森林生态系统的植物功能群划分标准也尚未统一,大多数研究将植物八大类型作为划分依据[20-21],少数研究依据植物功能性状或生态适应特征划分[22-23]。本研究以植物的生长型为基础,结合对温度、光照、水分的生态适应特征作为划分因子划分植物功能群,揭示植物-环境的动态平衡机制,有助于分析桉树引种对植物物种多样性的影响。
云南省是桉树引种的主要省份之一,澜沧县是大规模种植桉树纯林的典型区[24],但近年来对桉树林下植物多样性的研究集中在两广地区[8,11],而针对云南地区桉树林下植物多样性的研究较少,难以回答在生物多样性丰富的云南引种桉树是否对它会有影响。因此,本研究以云南澜沧县桉树大规模引种区为研究区,以不同原用地类型的桉树纯林及次生常绿阔叶林、人工思茅松(Pinus kesiya)林为研究对象,比较分析桉树林与参照林林下植物功能群的物种组成和物种多样性差异,揭示桉树引种后植物与环境的动态变化过程,旨在回答桉树引种是否影响植物多样性的争议问题,并为桉树引种区的生态保护提供理论支撑和科学依据。
1 材料与方法
1.1 研究区概况
澜沧拉祜族自治县(简称“澜沧县”)位于云南省西南部(22°31′—23°16′N,99°29′—100°35′E),国土面积8 807 km2,是省内面积第二大县[25]。澜沧县属于典型亚热带山地季风气候,雨量充沛,日照充足,垂直气候明显,年均气温19.10 ℃,年均降雨量1 624 mm。澜沧县植被类型多样,水平地带性植被属季风常绿阔叶林,垂直地带性植被分异明显,常见植被类型包括常绿针叶林、季风常绿阔叶林、山地雨林、季雨林、落叶季雨林、落叶阔叶林、半湿润常绿阔叶林等[26]。其中,桉树引种树种主要为尾叶桉(Eucalyptus urophylla)、巨桉(Eucalyptus grandis)及其杂交品系等,面积达到32 514.68 hm2,占县域总面积的3.73%[27]。
1.2 试验设计与样地调查
采用群落学调查法,设置20 m×20 m 的7 年生林龄桉树样地(原为次生常绿阔叶林地)6块、桉树样地(原为人工思茅松林地)6块,并在邻近位置选择立地条件相似的次生常绿阔叶林样地6块、人工思茅松林样地6块,作为参照样地。记录样地的经纬度、海拔、坡向、坡位、坡度、盖度等环境因子(见表1)。在样地中平均划分4 个10 m×10 m 乔木层样方,每木调查记录树种、胸径、树高等指标;在样地四角及中心划分5 个3 m×3 m 灌木、草本层样方,调查记录植物种类、株/丛数量、高度等指标。
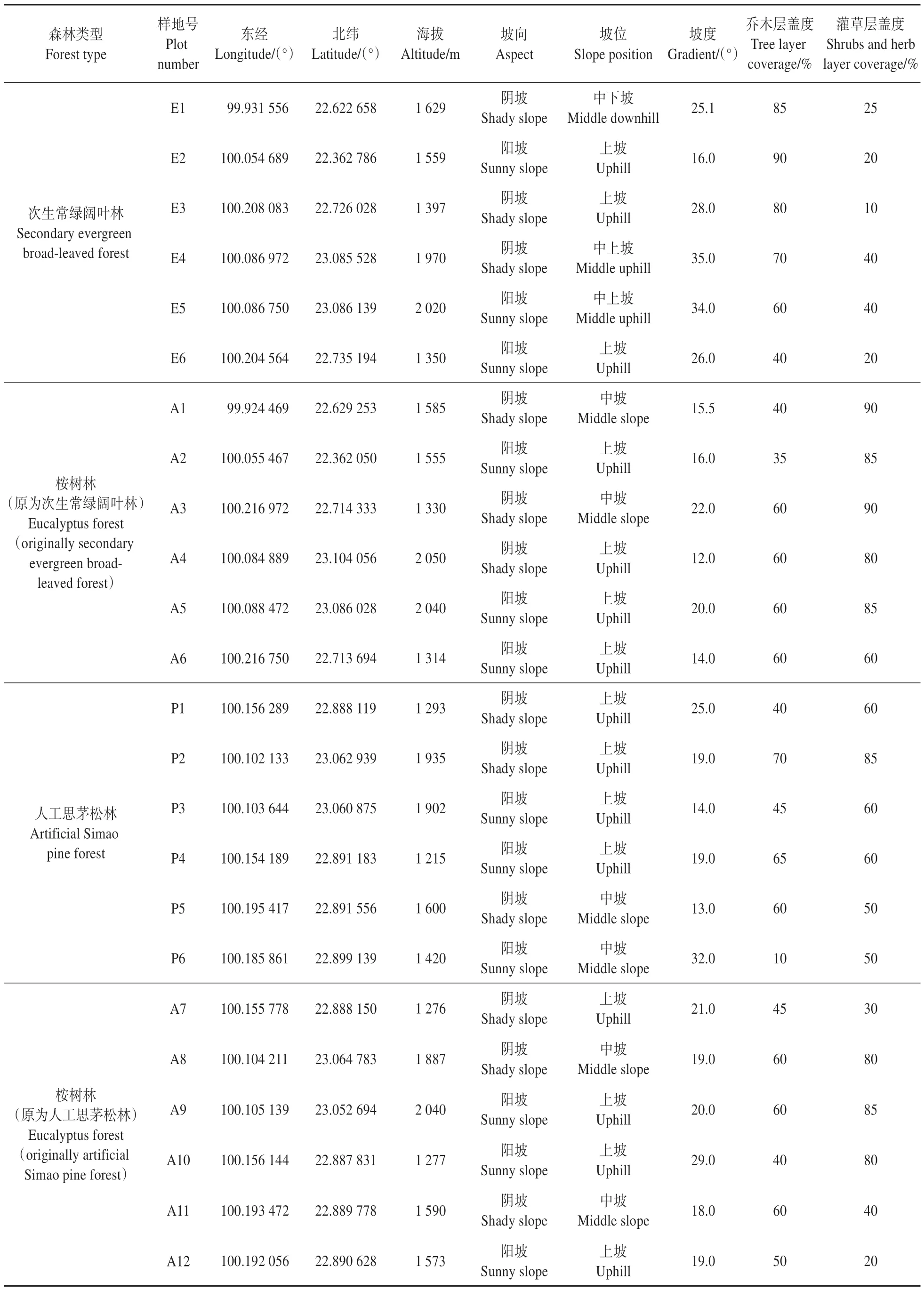
表1 样地概况Table 1 Sample plot overview
1.3 数据统计分析
1.3.1 植物功能群的划分方法
参考宋永昌[28]的植物功能群划分系统,鉴定植物的生长型及温度、光照、水分的生态适应特征(见表2),建立物种-特征矩阵,运用K-prototype 聚类分析方法,划分植物功能群。

表2 植物功能群划分系统Table 2 Division system of plant functional groups
1.3.2 数据处理与分析
根据重要值判断各功能群的优势种;采用α-多样性的4 个物种多样性指数测度各功能群的物种多样性,并进行差异性分析。
重要值(VI)计算公式:
式中:Dr表示相对密度;D表示某物种的个体数量;Hr表示相对高度;H表示某物种的高度;Fr表示相对频度;F表示某物种的频度。
物种多样性指数包括:Margalef 丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数[29]。
采用单因素方差分析(One-Way ANOVA)和LSD 事后多重比较检验桉树林与参照林林下植物功能群物种多样性指数的差异,显著性水平设为α=0.05。
2 结果与分析
2.1 植物功能群的划分
桉树及参照样地共调查到林下植物219种,隶属75 科165 属,将其聚类为2 类植物功能群,即暖性-阳生-中生功能群和温性-阴生-喜湿功能群(P<0.01)。暖性-阳生-中生功能群的植物有58 科124属152 种,其中常绿乔木、多年生草本植物种类最多,常绿灌木中等,暖性、阳生、中生植物居多;温性-阴生-喜湿功能群的植物有40 科60 属67 种,其中常绿灌木、温性、阴生、喜湿植物种类最多。
2.2 桉树林与参照林林下植物功能群的物种组成对比分析
2.2.1 桉树林与参照林林下植物功能群的物种科属种对比分析
如表3 所示,次生常绿阔叶林与桉树林对比,暖性-阳生-中生功能群的属、种数量是次生常绿阔叶林少于桉树林,而科数量相反;温性-阴生-喜湿功能群的科、属、种数量均是次生常绿阔叶林多于桉树林。表明桉树林与次生常绿阔叶林相比,内部温度较高、郁闭度较低、水分较少,暖性-阳生-中生功能群的植物更适应桉树林的环境条件。
人工思茅松林与桉树林对比,暖性-阳生-中生功能群的科、属、种数量均是人工思茅松林多于桉树林,而温性-阴生-喜湿功能群的科、属、种数量则相反。表明桉树林与人工思茅松林相比,内部温度较低、郁闭度较高、水分较多,温性-阴生-喜湿功能群的植物更适应桉树林的环境条件(表3)。
如表4 所示,次生常绿阔叶林中,暖性-阳生-中生功能群的物种重要值>5%的有杯状栲(Castanopsis calathiformis)、水锦树(Wendlandia uvariifolia)、木姜子(Litsea pungens),其中杯状栲重要值最大,为17.43%;温性-阴生-喜湿功能群的物种重要值>5%的有铁芒萁(Dicranopteris linearis)、山姜(Alpinia japonica)、荩草(Arthraxon hispidus)、粉背菝葜(Smilax hypoglauca),其中铁芒萁重要值为36.39%,占绝对优势。桉树林中,暖性-阳生-中生功能群的物种重要值>5%的有紫茎泽兰(Ageratina adenophora)、飞机草(Chromolaena odorata),重要值分别为44.31%、15.59%;温性-阴生-喜湿功能群的物种重要值>5%的有荩草、鳞毛蕨(Dryopteris chinensis)、岗柃(Eurya groffii),其中荩草重要值为39.07%,占绝对优势。表明桉树林替代次生常绿阔叶林后,暖性-阳生-中生功能群的优势种由杯状栲变成紫茎泽兰和飞机草;温性-阴生-喜湿功能群的优势种由铁芒萁变成荩草。
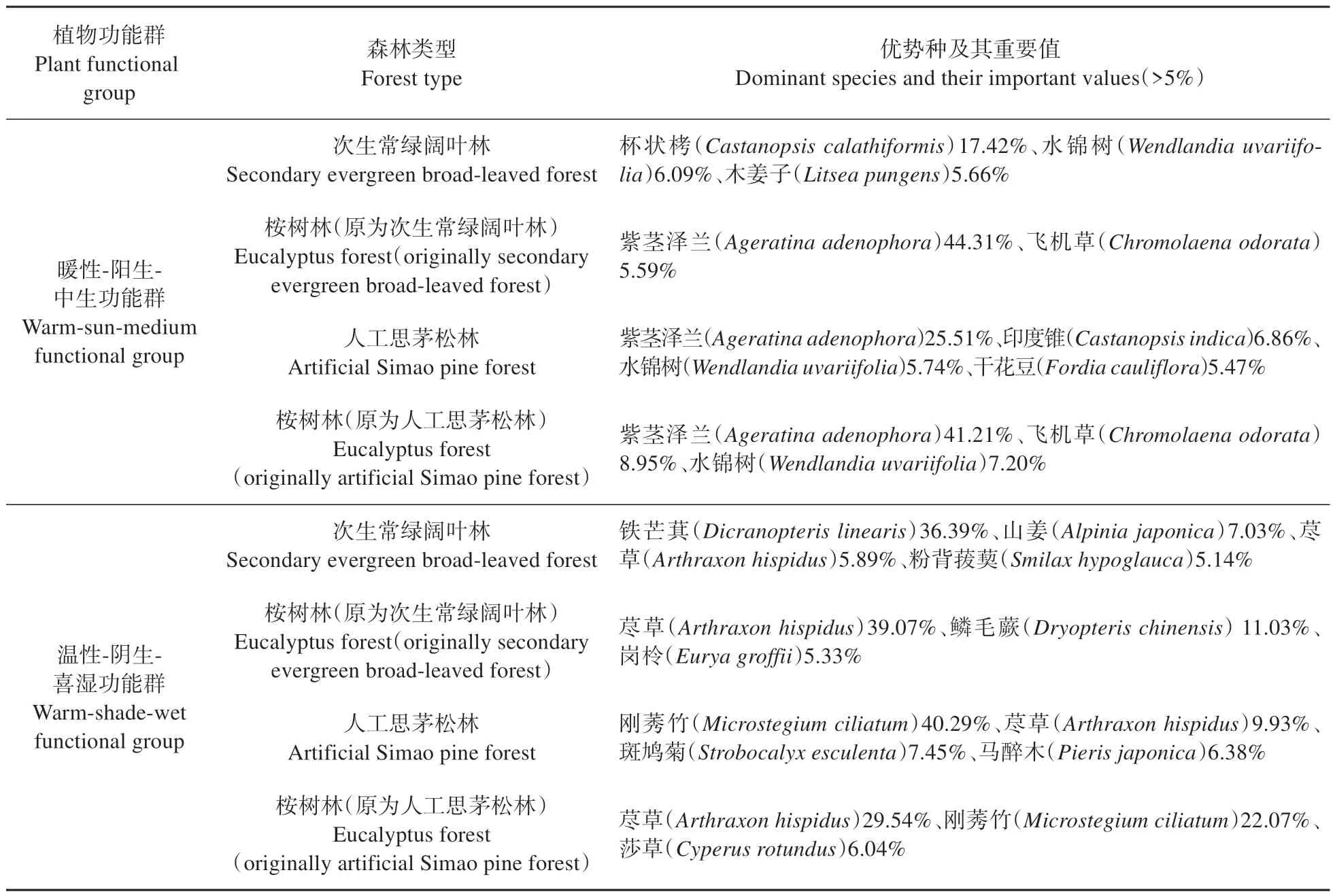
表4 不同林地各植物功能群的优势种及其重要值Table 4 Dominant species and their important values of plant functional groups in different woodlands
人工思茅松林中,暖性-阳生-中生功能群的物种重要值>5%的有紫茎泽兰、印度锥(Castanopsis indica)、水锦树(Wendlandia uvariifolia)、干花豆(Fordia cauliflora),其中紫茎泽兰的重要值最大,为25.51%;温性-阴生-喜湿功能群的物种重要值>5%的有刚莠竹(Microstegium ciliatum)、荩草、斑鸠菊(Strobocalyx esculenta)、马醉木(Pieris japonica),其中刚莠竹重要值为40.29%,占绝对优势。桉树林中,暖性-阳生-中生功能群的物种重要值>5%的有紫茎泽兰、飞机草、水锦树,其中紫茎泽兰重要值为41.21%,占绝对优势;温性-阴生-喜湿功能群的物种重要值>5%的有荩草、刚莠竹、莎草(Cyperus rotundus),重 要 值 分 别 为29.54%、22.07%、6.04%。表明桉树林替代人工思茅松林后,暖性-阳生-中生功能群的紫茎泽兰仍占据绝对优势地位;温性-阴生-喜湿功能群的荩草取代了刚莠竹的绝对优势地位(表4)。
2.3 桉树林与参照林林下植物功能群的物种多样性指数对比分析
对比桉树林和参照林暖性-阳生-中生功能群的物种多样性指数,结果表明次生常绿阔叶林与桉树林的Margalef 丰富度指数无显著差异(P>0.05),Simpson 多样性指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数具有显著差异(P<0.05)。人工思茅松林与桉树林的Margalef 丰富度指数、Simpson 多样性指数无显著差异(P>0.05),Shannon-Wiener 多样性指数、Pielou 均匀度指数具有显著差异(P<0.05)。4 种多样性指数均是次生常绿阔叶林>桉树林、人工思茅松林>桉树林。这表明桉树引种后会显著降低暖性-阳生-中生功能群的物种均匀度,可能是少部分植物更适应桉树林的生境条件,大量繁殖,导致物种均匀度与多样性下降。对比桉树林和参照林温性-阴生-喜湿功能群的物种多样性指数,结果表明次生常绿阔叶林与桉树林、人工思茅松林与桉树林的Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou 均匀度指数均无显著差异(P>0.05), 4种多样性指数均是次生常绿阔叶林>桉树林、人工思茅松林<桉树林(见图1)。这表明桉树林替代次生常绿阔叶林后温性-阴生-喜湿功能群的物种多样性略微下降,而替代人工思茅松林后略微上升,但并无明显变化。因此,桉树引种对温性-阴生-喜湿功能群的物种多样性无显著影响。
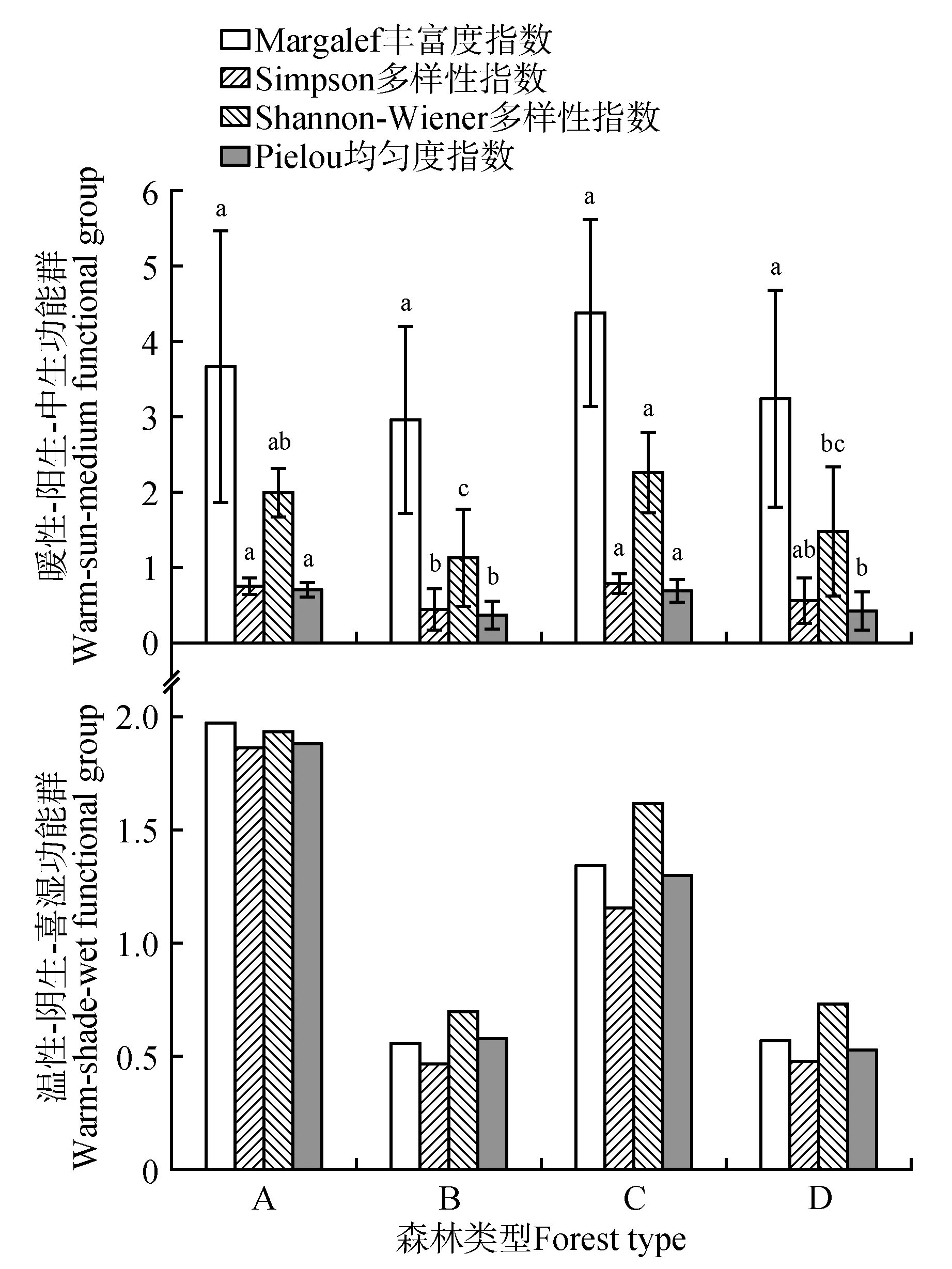
图1 不同样地类型各植物功能群的物种多样性指数A.次生常绿阔叶林;B.桉树林(原为次生常绿阔叶林);C.人工思茅松林;D.桉树林(原为人工思茅松林);图中不同小写字母表示同一指数在不同林型间差异显著(P<0.05)Fig.1 Species diversity index of plant functional groups in different land types A.Secondary evergreen broad-leaved forest;B.Eucalyptus fores(toriginally secondary evergreen broad-leaved forest);C.Artificial Simao pine forest;D.Eucalyptus fores(toriginally artificial Simao pine forest);Different lowercase letters in the figure indicated that the same index was significantly different between different forest type(sP<0.05)
3 讨论
桉树给引种区带来了较大的经济和社会效益,但桉树引种是否会引发生态问题在世界范围存在争议[30],其是否影响植物多样性的问题也尚不明确。
3.1 桉树引种对林下植物功能群物种组成的影响
桉树林替代次生常绿阔叶林后,会增加暖性-阳生-中生功能群的物种数量,减少温性-阴生-喜湿功能群的物种数量,导致温性-阴生-喜湿功能群退化;而桉树林替代人工思茅松林后,会增加温性-阴生-喜湿功能群的物种数量,减少暖性-阳生-中生功能群的物种数量,导致暖性-阳生-中生功能群退化。黄润霞等[31]的研究结果也表明不同林型林下植物的科属种存在差异。这是由于桉树林中生、耐旱植物的种类略高于次生常绿阔叶林,喜湿植物种类略少于次生常绿阔叶林;桉树林中生植物种类略少于人工思茅松林,喜湿植物种类略多于人工思茅松林。表明桉树引种后土壤水分条件稍变差。
桉树林替代两种参照林后,紫茎泽兰和飞机草均成为暖性-阳生-中生功能群的优势种,尤其是紫茎泽兰,重要值达40%以上,占有绝对优势。莫雅芳等[32]的研究结果不同林分下灌木和草本层优势种各不相同,优势种重要值特征也各不相同,此结论与本研究相似。这是由于桉树引种前清理林地的措施破坏了原有植被,而外来入侵植物紫茎泽兰生长力强,极易在裸地和稀疏植被的生境中定植生长,原有植被的恢复速度远不及紫茎泽兰和飞机草的繁殖扩散速度,导致桉树引种后林下植物的种类组成和优势地位发生变化。温性-阴生-喜湿功能群中荩草的重要值大幅增加,由5.89%~9.93%上升到29.54%~39.07%,具体表现是桉树林替代次生常绿阔叶林后,荩草取代铁芒萁的绝对优势地位;替代人工思茅松林后,荩草取代刚莠竹的绝对优势地位。这可能是土壤种子库中保留了一定的荩草种子,荩草适生力强、耐贫瘠,从而发展成为优势种,且桉树林的内部气候更适宜喜暖性的荩草生长,而对于喜温性的铁芒萁和喜亚高温的刚莠竹,内部气候条件并不是最适宜的。
3.2 桉树引种对林下植物功能群的物种多样性的影响
近年来,有学者进行了桉树林地与其他林地林下植物多样性的研究,如段文军等[33]比较尾巨桉人工林与乡土人工林林下植物多样性的差异,罗毓明等[10]分析了我国南方4 种常见人工林林下植物多样性特征等。但这些研究大都未考虑到桉树引种前的林地类型,试验设计的可对比性不足。本研究根据桉树引种前的林地类型选取2 类桉树林(原用地为次生常绿阔叶林的桉树林、原用地为人工思茅松林的桉树林),分别与参照林(次生常绿阔叶林、思茅松林)进行对比,分析桉树林与参照林林下植物多样性特征的差异,试验设计具有较高的对比性。
本研究结果表明桉树林替代次生常绿阔叶林后,暖性-阳生-中生功能群的Simpson多样性指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数显著下降(P<0.05),而温性-阴生-喜湿功能群的各物种多样性指数变化不显著(P>0.05)。这是由于次生常绿阔叶林暖性-阳生-中生功能群中如杯状栲、圆叶菝葜(Smilax bauhinioides)这类偏阳生、中生的木本、藤本植物生长环境良好,数量多,群落稳定性较高,有助于其他植物的生长。而桉树林下虽然光照条件较好,使得暖性-阳生-中生功能群产生许多新的草本植物,但数量较少,导致物种多样性较次生常绿阔叶林略小。因地理位置相近,气候条件相似,群落植物整体上对温度的适应性保持不变;7 年生成熟桉树林郁闭度适中,桉树引种后偏阳生植物的物种数并无显著变化,仅有略微增多;喜湿植物虽有略微下降,耐旱、中生植物虽有略微上升,但也并无显著差异。因此,Margalef丰富度指数无显著变化(P>0.05)。温性-阴生-喜湿功能群中草本、藤本植物种类略增加,灌木种类略减少;并且由于桉树的深根可以吸收更多水分,导致土壤水分减少,进而促使桉树林下阴生喜湿植物种类减少。因此,温性-阴生-喜湿功能群有所退化,但退化程度尚不明显。桉树林替代人工思茅松林后,暖性-阳生-中生功能群的Shannon-Wiener 多样性指数、Pielou 均匀度指数显著降低(P<0.05),而温性-阴生-喜湿功能群的各物种多样性指数变化不显著(P>0.05)。这可能是人工思茅松林暖性-阳生-中生功能群中木本、藤本植物数量多,而桉树林以草本植物占优势,木本植物仅有水锦树略显优势,群落稳定性不如人工思茅松林。并且7 年生成熟桉树林的郁闭度比人工思茅松林大,桉树引种后阳生、中生植物略微减少,导致暖性-阳生-中生功能群有不明显退化,而温性-阴生-喜湿功能群中木本植物略增加,温性喜湿植物略增加,但无明显变化。刘平等[34]对比分析桉树林与次生季风常绿阔叶林、撂荒地的植物多样性水平,结果显示在海拔1 700 m 以下,桉树林的Simpson 多样性指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数均小于其他两种林地类型与本研究结果相似。总之,桉树引种后,导致暖性-阳生-中生功能群的部分物种多样性指数降低,而不影响温性-阴生-喜湿功能群的物种多样性指数。
4 结论
桉树林替代次生常绿阔叶林后,会增加暖性-阳生-中生功能群的物种数量,减少温性-阴生-喜湿功能群的物种数量;而桉树林替代人工思茅松林后,会增加温性-阴生-喜湿功能群的物种数量,减少暖性-阳生-中生功能群的物种数量。
桉树林替代两种参照林后,林下植物种类组成及重要值改变,紫茎泽兰占据暖性-阳生-中生功能群的绝对优势地位,荩草占据温性-阴生-喜湿功能群的绝对优势地位;林下暖性-阳生-中生功能群的Shannon-Wiener 多样性指数、Pielou 均匀度指数显著降低,而对温性-阴生-喜湿功能群的各物种多样性指数均无显著影响。
本研究对象为7年生桉树林,虽然研究结果可以准确反映桉树林下植物多样性的特征,但存在未对桉树林下植物多样性进行长期监测的不足。因此,有待在未来的研究中对桉树人工林进行长期监测,探究桉树林自然生长15 年以上的植物多样性变化特征,更长时序的监测数据与研究结果回答桉树引种是否影响植物多样性的争议问题。
