基于有孔虫指数评估西沙群岛羚羊礁过去2600年的生态环境状况
2023-11-13梁日升余克服
梁日升,余克服
(广西大学 a.广西南海珊瑚礁研究重点实验室;b.海洋学院;c.中国珊瑚礁研究中心,南宁 530004)
Hallock等(2003a)于2003年基于珊瑚礁区不同类型底栖有孔虫对生态环境的指示意义提出了一种可评估珊瑚礁区生态环境状态的FORAM 模型,该模型的核心是有孔虫指数(Foram Index, FI)。FI是利用珊瑚礁区沉积物中不同类型的底栖有孔虫组合对珊瑚礁生态系统健康状态进行评估的生物指标,通过FI的高低可以评估珊瑚礁区海水环境状况是否适合珊瑚礁生长发育及在某些急性事件后珊瑚的恢复潜力。FI将珊瑚礁区底栖有孔虫分为3个种不同的功能群,分别为藻类共生种、非自养种和机会种(Prazeres et al., 2020)。藻类共生种通常指珊瑚礁区的大型底栖有孔虫(Larger Benthic Foraminifera, LBF)(Hallock, 2003b)。LBF 和珊瑚一样具有相似的光合共生体,与其他藻类共生,营养获取方式包括自养和异养,通常生活在营养和沉积物输入低、温度和盐度变化范围窄的环境中,与珊瑚生长所需的水质条件相似(Brown, 1997; Moberg et al., 1999; Hallock, 2003a, 2003b; Prazeres et al.,2018)。在以非常清澈、营养贫乏的海域为特征的健康珊瑚礁生态环境中,LBF在沉积物中占据主要比例,因此其FI值通常较高。常见的LBF包括马刀虫属(Peneroplis)、双丘虫属(Amphisorus)、马刺虫属(Calcarinidae)、异盖虫属(Heterostegina)、双盖虫属(Amphistegina)、小丘虫属(Soritidae)等(Hallock et al., 2003a; Prazeres et al., 2020)。非自养种有孔虫为较小的草食性和碎食性有孔虫类群,通常不存在共生藻,生长速度较快,生活在营养、光照和氧气含量充足的海域,其营养方式主要为异养型,当营养、食物等充分时而勃发,导致FI降低(Hallock et al., 1986; Prazeres et al., 2020)。非自养种主要包括玫瑰虫属(Rosalina)、抱环虫属(Spiroloculina)、五玦虫属(Quinqueloculina)、玦心虫属(Massilina)、三玦虫属(Triloculina)、多玦 虫 属(Schlumbergerina) 等(Hallock et al.,2003a; Prazeres et al., 2020)。机会种有孔虫指可以耐受高浑浊度、高营养和缺氧水体的有孔虫种属,在大多数珊瑚礁生态系统中占比较少只有在富含有机物和缺氧的环境中,机会种有孔虫才会大量生长,而这样的环境中几乎没有任何石珊瑚和LBF(Carnahan et al., 2009; Martínez-Colón et al., 2018),因此FI值通常很低。机会种有孔虫主要包括希望虫属(Elphidium)、小九字属虫(Nonionella)、转轮虫属(Ammonia)、箭头虫属(Bolivina)等(Hallock et al., 2003a; Prazeres et al., 2020)。FI有1~10的范围,其中FI<2 表示海域水质条件已不适合珊瑚生长;2<FI<4表示海域水质条件是珊瑚生长的边缘条件环境;FI>4 表明海域环境适合珊瑚生长和恢复(Hallock et al., 2003a)。
珊瑚礁区底栖有孔虫具有个体小、数量多、寿命短等特点(Cockey et al., 1996),因此FI 的获取相对简单,且具有代表性。自FI指数提出以来,已被较多地应用于全球珊瑚礁区研究。在珊瑚礁生态健康评估方面,Oliveira-Silva 等(2012)对巴西东部巴伊亚州近海珊瑚礁区的研究表明,FI与活珊瑚覆盖度有较好的相关性,在珊瑚礁保护区内FI 较高、珊瑚礁健康状况较好;Pisapia等(2017)对马尔代夫中部环礁的研究表明,无人岛海域的FI值比居住岛和旅游岛的高,珊瑚更健康;Chen 等(2017)利用FI 对东沙群岛珊瑚礁生态进行评估,认为目前东沙群岛珊瑚礁尚未从1998年白化事件中恢复,珊瑚健康情况较差,FI 为2.9。在珊瑚礁区海域营养状况的评估方面,A'Ziz 等(2021)对南海南部刁曼岛珊瑚礁的研究发现,FI值与良好的海域环境条件呈正相关,而与有机质富集的环境呈负相关;Fabricius等(2012)对大堡礁近岸珊瑚礁的研究发现,FI 与叶绿素浓度等营养指标呈反比关系,FI随着水域营养物质升高而下降。
迄今FI的应用还基本局限在现代珊瑚礁区,未涉及地质历史时期的珊瑚礁。若FI能用于对地质历史时期珊瑚礁健康状况的评估,则其意义更显著,一方面,因为地质历史时期的珊瑚礁(如南海各群岛)很少有大面积的露头可像现代珊瑚礁一样进行生态状况评估;另一方面,对地质历史时期珊瑚礁健康状况演化过程的了解,有利于理解珊瑚礁发育与气候的关系,以及进一步评估全球气候变暖背景下现代珊瑚礁的演化趋势。在对历史时期珊瑚礁的研究中,近年来南海及世界各珊瑚礁区开展了一系列的深钻。如西沙群岛琛航岛的琛科2井,珊瑚礁岩厚873.55 m,覆盖的历史为19.6 Ma(Fan et al.,2019),在对深钻小样品量岩芯的研究中,有孔虫以其个体小、数量多、代表性好等特点有其独特的研究优势,但尚未有学者利用有孔虫开展历史时期FI变化的相关研究。因此,本文以从西沙群岛羚羊礁的潟湖中钻取的LYJ2岩芯为材料(全长287 cm,对应的底部年代为2665 a BP),探索FI对历史时期珊瑚礁生态状况的指示意义,以期了解历史时期西沙群岛羚羊礁珊瑚的健康状况变化以及FI对气候变化的响应。
1 材料与方法
1.1 样品和研究区域
采 用 的 岩 芯(LYJ2,16°27′59.32″N、111°36′09.62″E)是2013年从羚羊礁封闭潟湖中采集所得,具体采集位置见(图1),岩芯取自羚羊礁的内礁坪区域(相对海平面以下150 cm),总长287 cm,采集过程中没有任何损失。LYJ2 底部年代约为2 665 a BP,其年代框架构建参阅文献(Yue et al.,2019),14C 测年所用材料为沉积物中的有孔虫(Calcarina sp.),LYJ2年代信息见表1所示。
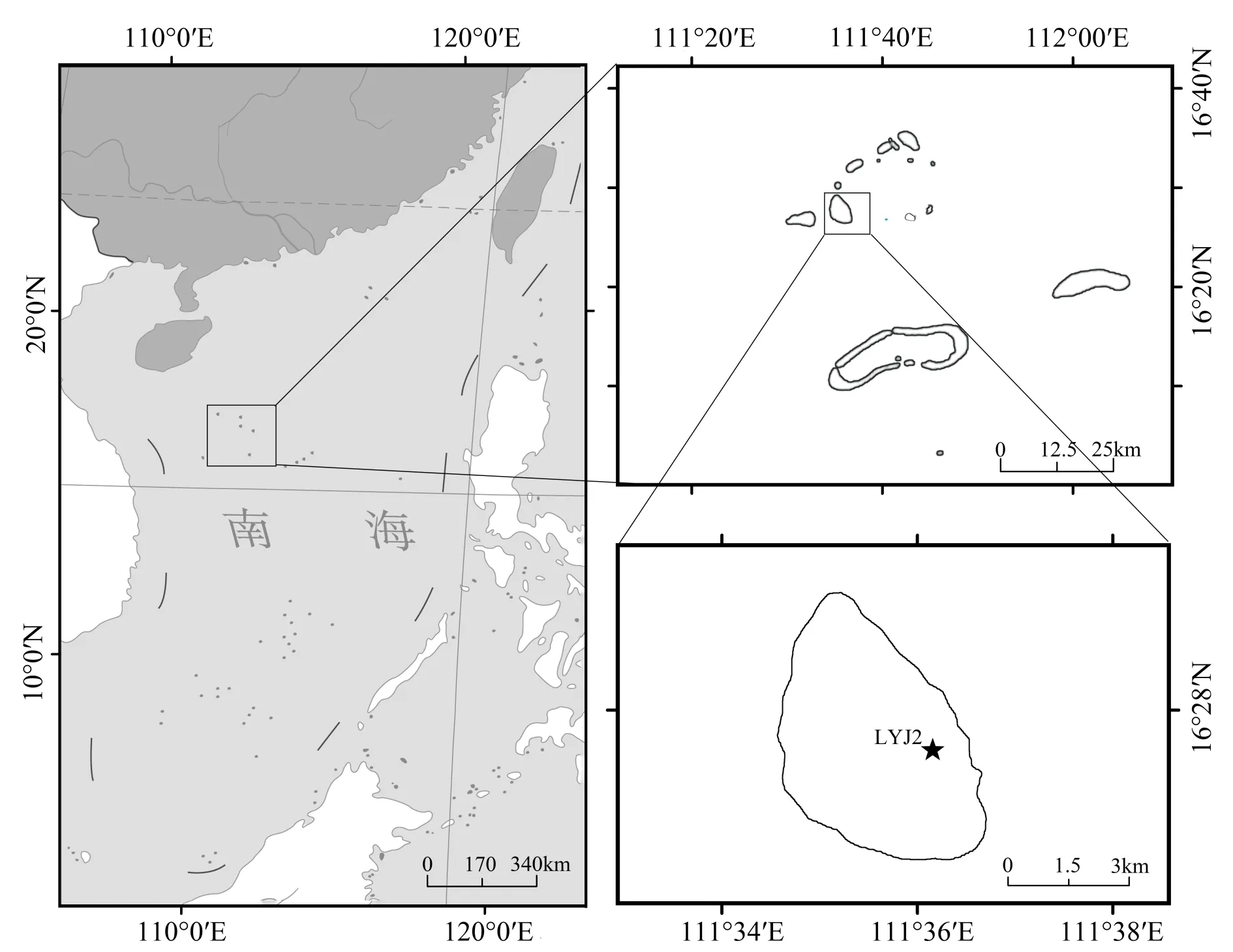
图1 研究区域及LYJ2岩芯取样地点Fig.1 Location of the study area and LYJ2 core sampling site
羚羊礁(16°28′N、111°35′E)位于中国南海西沙群岛内的永乐环礁西南处(见图1),是一个椭圆形礁体,东西宽度约为4 km,南北长约12 km,内部有长约5 km,宽约3 km 的封闭潟湖。根据西沙气象站1951―2001年的仪器数据记录,该区域年均温为26.6℃,海面温度较高,约为27.5℃。
1.2 有孔虫指数(FI)计算
以LYJ2 岩芯为材料,根据LYJ2 的年代信息,每隔0.5 cm采集用于有孔虫分析的沉积物样品,共计493份。所有样品先在48℃烘箱干燥48 h,然后将干燥的样品手动过2和0.064 mm筛网,以去除泥沙以及破碎有孔虫,将每份样品装袋保存,以进行后续的有孔虫鉴定。将样品放置在挑选器皿中,利用显微镜和细笔刷对每份样品进行有孔虫的鉴定和计数[3种功能群有孔虫常见代表种属见图2。有孔虫的鉴定参照郑守仪等(1979)和秦国权(1987)所记录的西沙群岛现代有孔虫组合描述,根据FI的计算,每份样品需统计150~200 枚有孔虫(Prazeres et al., 2020)]。不同底栖有孔虫被归类为3 种不同功能群体(藻类共生种、非自养种、机会种),在珊瑚礁区中,藻类共生种(LBF)数量较多,而非自养种和机会种数量和属种较少,因此在对样品有孔虫进行鉴别和统计时,以非自养种和机会种为主要鉴定对象确定数量,剩余底栖有孔虫根据其形态归类为LBF。FI 计算公式为(Hallock et al.,2003a):
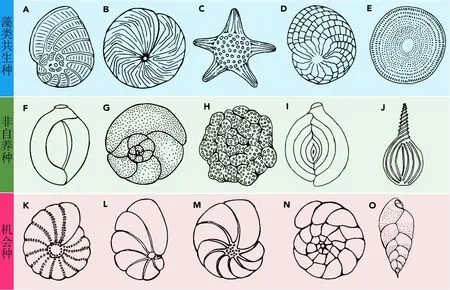
图2 三种功能群常见代表种属(Prazeres et al., 2020)Fig.2 Common representative species of three functional groups
式中:Ps=Ns/T;Ph=Nh/T;Po=No/T。T表示样品中被筛选出来的底栖有孔虫总数,Ns表示藻类共生种的有孔虫数量;Nh表示其他壳体小、生长快的非自养小有孔虫数量;No表示耐受高浑浊度、高营养和缺氧水体的机会种有孔虫数量;Ps表示藻类共生种在样品中的占比;Ph表示非自养种在样品中的占比;Po表示机会种在样品中的占比。FI范围为1~10,其值伴随海域营养条件升高而出现降低(Pisapia et al., 2017)。当FI>4 时,藻类共生种有孔虫占据沉积物主要成分,表示该海域环境条件适合珊瑚生长;当2<FI<4时,表示该海域环境条件处于珊瑚生长的边缘环境;当FI<2 时,适应高营养和缺氧水体的机会种有孔虫占据主要成分,表示该海域环境条件不适合珊瑚的生长(Hallock et al., 2003a)。
2 结果分析
本文共获得493个FI值,结合表1的LYJ2年代控制点,通过内插的方法获得FI 的年代序列(图3),时间间隔为16 a。结果显示,羚羊礁海域FI值在4.1~7.9 范围波动,均值为5.9,最高值在572 a BP出现,为7.9,最低值为4.1,出现2次,在52和18 a BP。

图3 三种功能群底栖有孔虫时间序列(a)和LYJ2岩芯FI时间序列及SRSD处理结果(b)Fig.3 Time series of three functional groups(a) and LYJ2 core FI time series and SRSD results (b)
用序列状态转换检测方法(Sequential Regime Shift Detection, SRSD)分析FI 在过去2600 年期间的变化幅度和趋势(Rodionov, 2004)。SRSD 是用于对时间序列的年代际变化处理的一种算法,基于数据的平均值以及加权数据等对时间序列的变化趋势进行分析。SRSD 方法是在Excel 中使用软件包Registy shift test-v6.2.xlsm①软件包见https:/sites.google.com/site/climatelogic/。对数据进行处理。
结果表明(见图3),FI在2665 a 期间表现出2个显著特征:1)波动性。整体上FI 变化表现为不同时期的先波动下降后波动上升趋势。可分为3个明显的上升期:2380―1628、1212―572、252―92 a BP,3个明显的快速下降期:1628―1212、572―252、92 a BP,1个小幅下降期:2665―2380 a BP;2)周期性。FI 在长期变化趋势上叠加了不同尺度的年代际波动(图4),变化呈现不稳定性,存在较为明显的66.7、54.4 a等变化周期。

图4 过去2600年FI的显著周期Fig.4 Significant cycle of FI for over the past 2600 years
3 讨论
3.1 过去2600年羚羊礁珊瑚礁总体处于健康状况
过去2600 年期间羚羊礁海域FI 在4.1~7.9 范围波动,平均值为5.9。依据FI 对生态状况的指示意义(Prazeres et al., 2020),当FI>4时,表明海域环境适合珊瑚等藻类共生生物的生长发育,该海域珊瑚处于良好的健康状态。该观点在全球许多珊瑚礁区也得到印证,如加勒比海区(Velásquez et al.,2011)、巴西巴伊亚州(Oliveira-Silva et al., 2012)、马尔代夫阿里环礁(Pisapia et al., 2017)、南海东沙群岛(Chen et al., 2017)、南海南部刁曼岛(A'Ziz et al., 2021)等。据此,本文推测过去2600 年羚羊礁珊瑚礁总体处于健康状况。
珊瑚在生长发育的过程中通常需要较为苛刻的环境条件,主要受到温度、光照、盐度、浑浊度等自然因素的调控(余克服,2018)。羚羊礁位于西沙群岛西部,远离大陆,没有陆地径流等因素的影响,且除近代以来,羚羊礁珊瑚生态系统基本没有受到人类活动的影响。因此,海温是影响过去2600年羚羊礁珊瑚礁生态健康的主要因素。造礁珊瑚最适宜在年均海温25~28℃的海域内生长,温度的异常升高或降低都会对珊瑚礁的生态健康造成破坏。近年来,部分学者在西沙群岛开展了一些列利用古珊瑚化石重建海温的研究,如陶士臣等(2021)利用西沙群岛滨珊瑚生长率,重建得出西沙海域近500 年的海温变化范围为26.13~28.43℃;黄博津(2013a)和张会领(2014)等重建的西沙海域晚全新世不同时期的海温结果在26.5~27.27℃波动,其均在珊瑚生长的适宜范围;Jiang等(2021)利用西沙群岛的3 个化石珊瑚重建的罗马暖期期间(2070-1740 a BP)西沙海域年平均海温变化未超过珊瑚生长适宜的海温阈值。综上所述,过去2600年西沙群岛海域海温均在适合珊瑚生长发育的范围内波动,表明过去2600年西沙群岛羚羊礁珊瑚礁生态系统基本保持健康,与FI所揭示的结论相符。
3.2 FI与海温变化之间具有较好的对应关系
晚全新世是距离现代最为接近的一个地质年代时期(Wanner et al., 2008),过去2600年大致存在5个特征气候时段,分别为:罗马暖期(-300-300 AD)、黑暗时代冷期(300-800 AD)、中世纪暖期(900-1400 AD)、小冰期(1500-1850 AD)和20世纪异常暖期(黄博津 等,2013b)。为探讨FI 对海温变化的响应,将重建的2600 年FI 与已有研究构建的西太平洋SST、石笋重建的气温等温度序列(图5)进行对比,发现在冷期时段,FI相较之前有所下降,暖期时段上升,据此推测海温对FI变化产生影响。
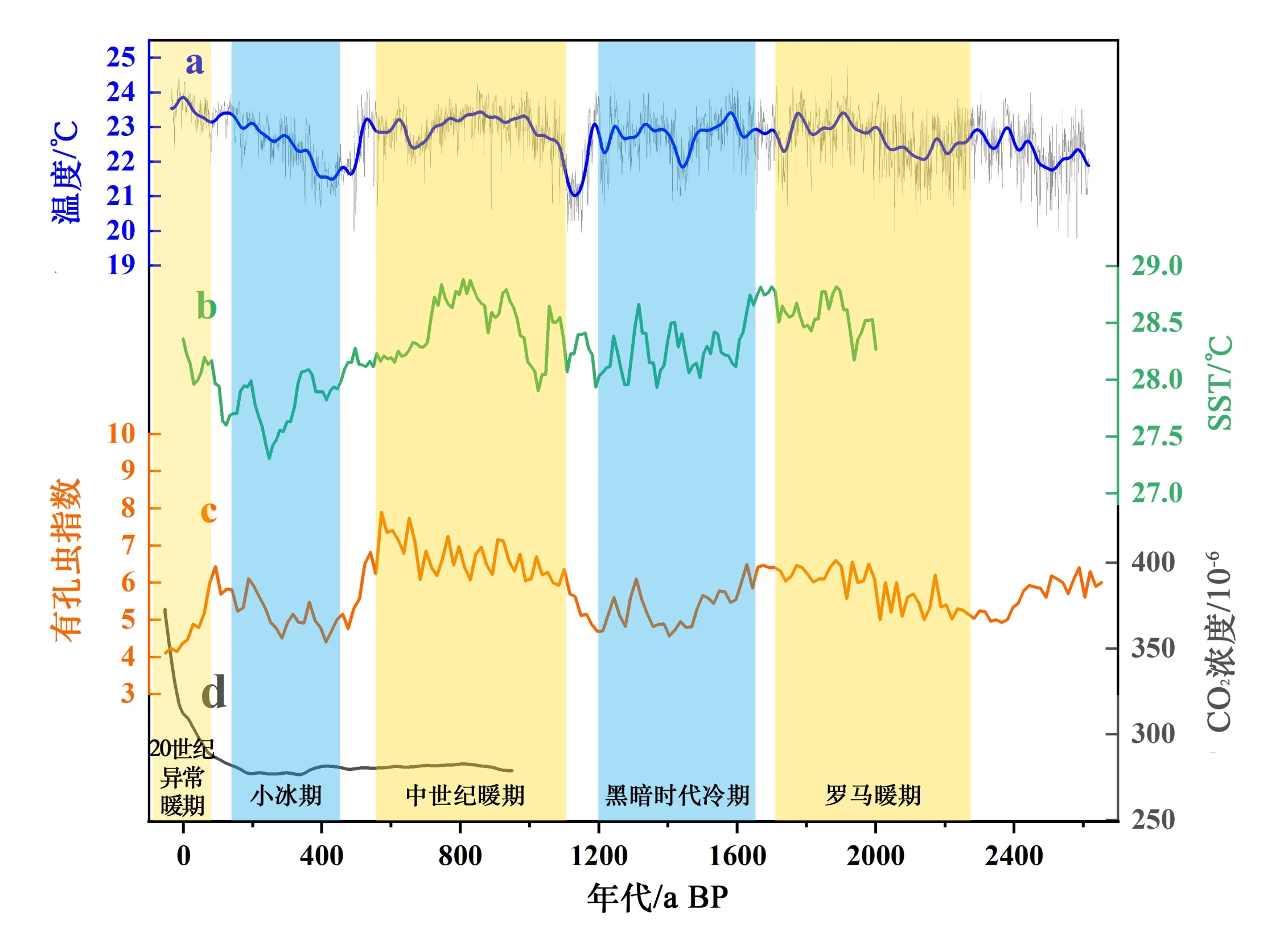
图5 FI结果和其他气候记录的比较Fig.5 FI results compared with other paleoclimate records
FI 的2 个明显下降阶段(1628-1212、572-252 a BP)与黑暗时代冷期、小冰期对应。其中,黑暗时代冷期(300-800 AD),LYJ2 岩芯年代框架对应时期约为1212-1648 a BP。根据深海沉积物有孔虫记录的SST,黑暗时代冷期的SST比现代低约0.5℃(Saenger et al., 2011)。由图5 可知,FI 于黑暗时代冷期阶段内,相较之前发生小幅度下降,表明冷期时段海温对珊瑚礁区大型底栖虫的数量产生影响。与黑暗时代冷期相似的特征气候时期为小冰期(1500-1850 AD),在LYJ2 岩芯中对应时期约为141-457 a BP。小冰期指紧随着中世纪暖期之后一个相对而言较冷的特征气候时段,许多代理指标均记录了小冰期的低温现象(Ogilvie et al., 2001;Mann et al., 2009),本文中FI 也存在明显下降,与黑暗时代冷期记录的下降趋势一致。整体上,FI的下降时段和2个冷期相对应,表明温度降低对LBF的数量产生影响,FI的降低在一定程度上记录了历史时期的降温事件。
FI的2个上升阶段(2380-1628、1212-572 a BP)与中世纪暖期、罗马暖期对应。中世纪暖期指相对小冰期出现的一个明显的温暖气候时期,罗马暖期是罗马帝国从建立到灭亡的一段相对温暖时期。LYJ2中FI在2个时期出现明显的升高,且持续时间与暖期相似。根据室内实验发现,在一定温度范围内,温度升高会导致LBF的钙化率和存活率增高(Fujita et al., 2014),这可能是暖期FI 较高的原因之一,不同LBF 对于温度升高的响应有所差异(Waters et al., 2017),这可能导致FI 出现波动。整体上,FI的上升期与暖期相对应,是对升温事件的较好响应。
综上,由重建的FI 与SST 等指标对比可发现,FI在百年尺度上与气候变化具有一致性,FI的高值和低值区间与暖期和冷期大致对应,海表温度是影响羚羊礁的FI值主要因素之一,表现为海温升高,FI升高,反之下降。
3.3 1628—1404 和572—252 a BP 两个冷期FI下降因素分析
1628―1404 和572―252 a BP 的FI 表现出快速下降的趋势,分别与黑暗时代冷期和小冰期对应。依据FI的指示意义(Hallock et al., 2003a),导致FI下降的原因可能羚羊礁海域水体营养物质增多有关,重建的历史记录也显示西沙地区在冷期时,海洋生产力较高(Liu et al., 2002; Han et al., 2019)。据此推测,FI在冰期下降可能是由于冷期羚羊礁海域的营养物质增加所致。
相对其他时期,黑暗时代冷期和小冰期的海温较低,冬季风强度较高。受强烈的冬季风的影响,冰期表层海水温度较低,促进海域上下层海水混合以及营养物质的交换,导致表层海水营养物质增加(Huang et al., 1997; Liu et al., 2002)。同时,冰期较强的冬季风从亚洲大陆等地携带大量的气溶胶粉尘到达西沙群岛海域上空,气溶胶粉尘中的营养元素会提高海域初级生产力,导致海域营养物质增加(Liu et al., 2009),重建的历史记录也显示,西沙地区在冬季风较强时海洋生产力较高(Liu et al.,2002; Han et al., 2019)。此外,降雨的增加会加速空气中的营养元素沉降到海水,导致海水营养物质升高。根据重建的西沙2000 年南方涛动指数(Southern Oscillation Index, SOI,赤道西太平洋和东南太平洋两处极点的气压差)结果可知(Yan et al., 2011),小冰期和黑暗时代冷期的SOI均为正偏状态,表明西沙群岛在小冰期和黑暗时代冷期处于拉尼娜状态,降雨量相对较多。通过与SOI 指数(图6)对比发现,SOI 和FI 之间存在负相关关系(R=-0.52,P<0.01),指示西沙群岛在降水较多的拉尼娜时期,FI出现下降,这与已有研究发现的拉尼娜时期FI较低的结论(Fabricius et al., 2012; Clemens et al., 2018)相符。
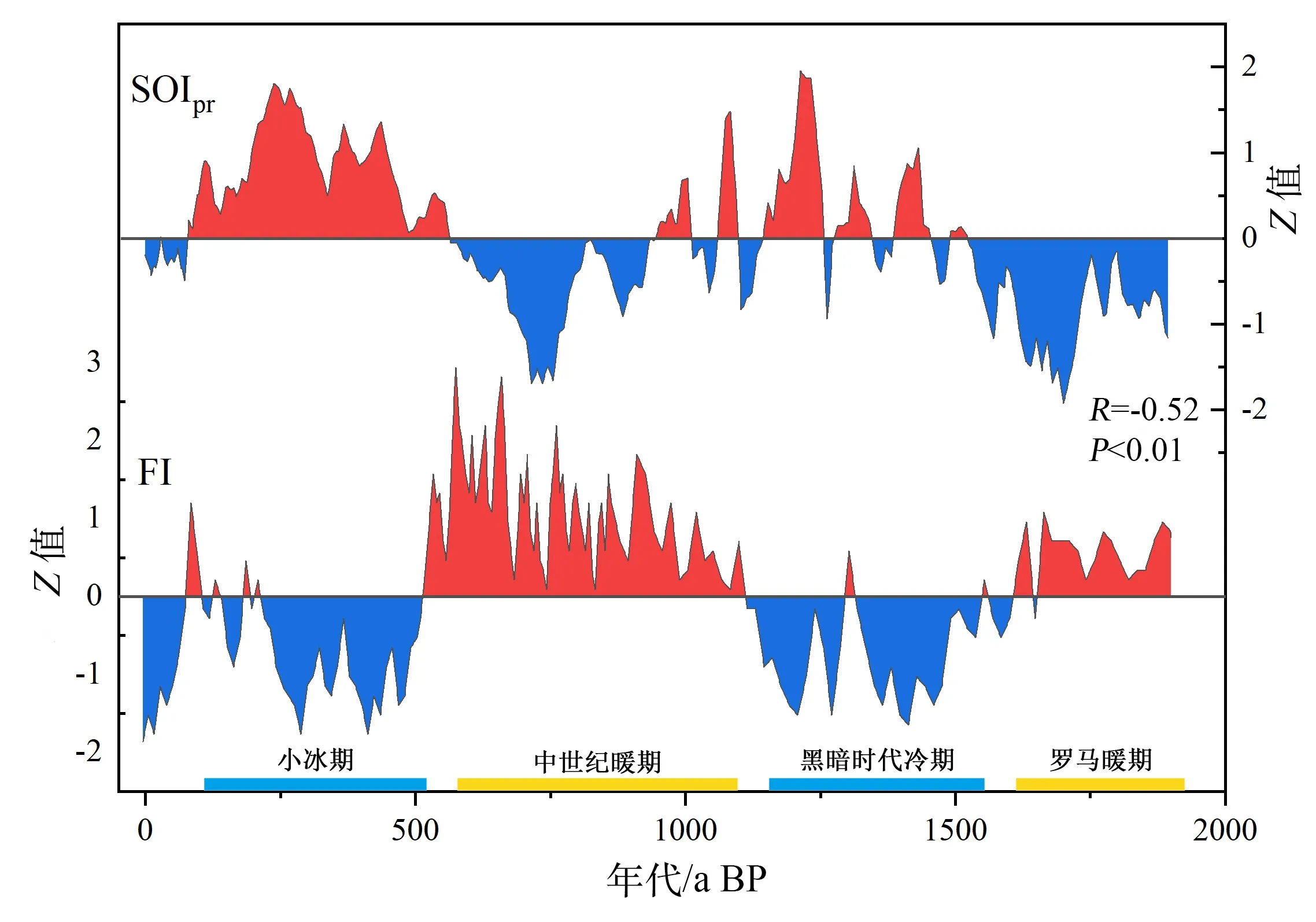
图6 近2000年LYJ2的FI与重建的SOI指数(Yan et al., 2011)比较Fig.6 Comparison between the FI of LYJ2 and the reconstructed SOI index(Yan et al., 2011) in recent 2000 years
综上,两个冰期FI的快速下降,表明羚羊礁海域营养物质出现增加,这与冰期西沙群岛较高的生产力状况符合,FI下降的原因可能是由于冷期冬季风增强导致大气粉尘物质增多、降雨增加、羚羊礁海域的营养物质增加所致。
3.4 现代时期FI快速下降因素分析
基于FI 变化情况可知,过去2600 年羚羊礁珊瑚健康状况基本保持良好,总体呈现健康状态,但92a BP至今,FI出现非常明显的下降趋势,与上文关于海温上升对应FI升高的推论相矛盾。这是工业时期以来的温度异常升高时期(20世纪异常暖期),人类活动的影响逐年加剧(Jones et al., 2004)。目前,在全球气候变暖,人类活动增加的威胁下,全球大部分珊瑚礁都已发生严重退化(Hoegh-Guldberg, 2010)。极端的高温导致全球珊瑚礁出现大面积的白化,南海珊瑚礁健康状况亦出现较大幅度的退化,西沙群岛珊瑚礁覆盖度从原来的90%下降至不到10%(吴钟解 等,2011;余克服,2012;Chen et al., 2019)(图7)。温室气体(如CO2)排放增加(见图5),不仅加速了气候变暖,还导致海水酸化,进而降低珊瑚钙化速率,甚至导致珊瑚死亡(Anthony et al., 2008)。频繁的人类活动(过度捕捞、破坏性渔业、工程建设等)更是对西沙群岛珊瑚礁生态环境造成较为严重的破坏(Zhao et al., 2012)。同时,由于南海周边地区的人类活动(工业和农业)加剧,由此产生的氮、磷等物质排放增加,南海的大气养分增多,在季风盛行期间,西沙海域大气氮沉降通量变多,导致西沙海域营养物质增加,大型藻类数量增加,进而破坏珊瑚礁的生态健康(Duce et al., 2008; Chen et al.,2019)。此外,近年来西沙群岛发生的长棘海星爆发、珊瑚疾病爆发等自然因素也使西沙海域珊瑚礁生态环境出现恶化(Tang et al.,2019;李元超 等,2019)。总之,由于人为、自然等因素,西沙群岛珊瑚礁生态环境在近40 年来现退化,这与FI 快速下降所指示的较差的珊瑚礁生态环境一致。
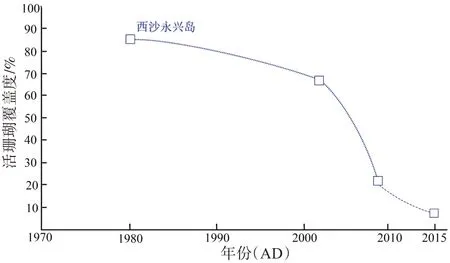
图7 南海珊瑚礁的退化(余克服,2012;Chen et al., 2019)Fig.7 Degradation of Coral Reefs in the South China Sea(Yu, 2012; Chen et al., 2019)
综上,92 a BP 至今FI 的快速下降,表明羚羊礁珊瑚礁生态健康状况出现恶化,这与近40 年来西沙群岛珊瑚礁生态系统的快速退化相对应,其可能原因包括人类活动加剧、大气氮沉降通量增加导致的羚羊礁海域营养浓度升高等。
4 结论
本文以西沙群岛羚羊礁LYJ2岩芯为材料,重建了过去2600 年以来的FI 变化序列,结合SST、特征气候时段、SOI 等指标进行对比分析,得出如下主要结论:
1)过去2600 年南海西沙群岛羚羊礁区海域FI 变化幅度为4.1~7.9,平均为5.9,据此推测近2600年西沙羚羊礁海域环境均适合珊瑚生长,珊瑚礁总体为健康状态。
2)FI 在百年尺度上与气候变化具有一致性。通过FI值与海表温度的对比分析,推测海表温度会对FI产生一定影响,FI高值与中世纪暖期、罗马暖期大致对应,而低值与小冰期、黑暗时代冷期大致对应。
3)1628―1404、572―252 a BP 两个冰期阶段FI快速下降的原因可能是由于冷期冬季风增强导致大气粉尘物质增多、降雨增加、羚羊礁海域的营养物质增加所致。
4)92 a BP至今FI的快速下降与近40年来西沙群岛珊瑚礁生态系统的快速退化相对应,其可能原因包括人类活动加剧、大气氮沉降通量增加导致的羚羊礁海域营养浓度升高等。
